配对静水论坛
静水论坛 时间:2021-01-26 阅读:()
第卷第期,年月水生生物学报!
,,卵巢发育阻滞的人工三倍体鱼减数分裂染色体配对的光镜观察桂建芳陈丽梁绍昌蒋一建中国科学院水生生物研究所,武汉.
.
提要采用界面铺张制片和硝酸银一步染色的方法,对人工三倍体水晶彩螂卵巢发育阻滞型个体的减数分裂染色体配对进行了光镜观察.
在分化有初级卵母细胞的卵巢发育阻滞型的三倍体鱼中,减数分裂粗线期细胞主要由二价体和单价体组成,也见有少量三价体和其它多价体,其染色体成员数大多在左右在不同细胞间,染色体的大小变化较大配对联会过程中形成的配对又和产生的特异蛋白在一些细胞中明显可见.
文中讨论了三倍体染色体配对紊乱与卵母细胞发育受阻的关系,并认为人工三倍体鱼是进行减数分裂染色体配对和联会机理研究的好材料.
关键词人工三倍体鱼,卵巢发育,减数分裂,染色体配对人工三倍体鱼大多数是不育的【'·",其不育的主要原因一般只是依据植物三倍体中的观察,认为是紊乱的减数分裂染色体配对和分离所致〔,而有关人工三倍体鱼减数分裂染色体行为的研究迄今还未见报道.
本研究是在作者采用静水压休克批量生产出三倍体水晶彩螂〔〕,并在雌性三倍体鱼中发现有多种卵巢发育类型,可明显区分为阻滞型、间性型、败育型和极个别正常型「习的基础上,采用界面铺张法制片和硝酸银快速一步银染的方法,首先对人工三倍体水晶彩螂卵巢发育阻滞型个体的减数分裂前期染色体配对等行为进行了光镜的初步观察.
材料与方法材料人工三倍体水晶彩螂是在卵子受精后采用耐的静水压持续处理诱导而成.
用作制片观察的卵巢发育阻滞型个体共尾,制片同时分别进行了组织切片的细胞学观察.
根据形态和组织学观察,卵巢发育阻滞型的三倍体雌鱼又可分为两个亚类,一类卵巢呈细线状,仅含有卵原细胞另一类呈线状,已分化有较多的初级卵母细胞〔.
用作制片观察的前一类鱼为,尾,后一类鱼为尾.
作为对照,还进行了尾雄性二倍体水晶彩螂精巢细胞的制片.
界面铺张法制片界面铺张法制片主要参考和马昆等叨报道的方法进本工作得到中国科学院青年科学基金和院长基金的资助.
武汉大学生物学系届毕业实习生.
年月日收到一年月修回.
水生生物学报一卷行.
首先剥净卵巢或精巢上的脂肪组织,取组织块洗净,于.
培养液中制成细胞悬液,静置左右,取上层细胞悬液经离心一次后,加少许生理盐水制成细胞悬液.
然后在一涂有石蜡的平皿上滴加一直径约的低渗液液珠,取少量细胞悬液与液珠表面接触,细胞即在液珠表面铺张.
约后,用一个干净的玻片与载有细胞的液珠表面接触,细胞即附着于玻片上.
平置一,将玻片置于新配制的并多聚甲醛一中固定,取出凉干.
硝酸银染色凉千的玻片按和报道的一步银染法进行硝酸银染色.
光镜观察.
结果在卵巢发育严重阻滞的第一亚类的尾三倍体鱼中,正如组织切片的细胞学观察很难找到初级卵母细胞一样,其染色体制片上也没有见到减数分裂的前期细胞.
在已分化有较多初级卵母细胞的第二亚类发育阻滞型三倍体鱼的卵巢细胞染色体制片上,均可见到一定比例的减数分裂粗线期细胞.
其共同的特征之一是染色体配对紊乱.
在同一细胞的染色体扩散相图版、和中,总可见到部分染色体染色较浅,呈细丝状,相互缠绕严重,难于区分,为没有配对的单价体大部分染色体已配对,染色较浓、较粗,相互之间易区分,主要为二价体,少数可明显看出是多价体.
在多价体中,有已配对的二价体再与一价体排列,并在一些点和区段发生配对的三价体图版有由两条已联会的二价体通过端粒联合形成的四价体图版也有由多条非同源染色体相互交叉折叠配对形成的复合多价体图版.
在一些尚未联会完成的二价体上,配对叉清晰可见图版.
在一些已完成联会的染色体上,着丝粒区域与其它区段的结构差异相当显著,着丝粒区域细且着色浅,与粗且着色深的其它区段形成强烈的对比图版、.
多数细胞中的单价体、二价体和多价体的总染色体成员数一般在左右图版一.
与这些三倍体鱼相比,由二倍体鱼精巢细胞制备的粗线期染色体比较清晰,染色体配对正常,粗细一致,条二价体清晰可数图版.
此外,粗线期染色体的另一个显著特征是其细胞相互之间在结构和大小上的变化图版、、和,它们既表现有染色体的粗细之分,又表现有染色体的长短不同.
讨论三倍体水晶彩鲡含有三套染色体组,共个染色体.
从理论上推测,在同源染色体配对及联会过程中,有可能出现下列三种情况一是形成条二价体,条单价体二是形成条三价体三是单价体、二价体、三价体甚至更高的多价体同时并存.
从卵巢发育阻滞型三倍体鱼粗线期染色体配对图象来看,与第三种情形基本相似,主要由二价体和单价体组成,也含有少量三价体和其它多价体,其染色体成员数一般在左右.
已配对联会了的染色体发生了较大程度的凝缩,而未配对的单价体仍呈丝状,相互缠绕.
从这种染色体结构上的显著差异所造成的紊乱现象来看,它必定影响了细胞的继续发育和生长,导致发育阻滞.
然而,同样是人工三倍体鱼,为什么少数个体又能正常发育形成成熟的卵子呢为什么在人工三倍体水晶彩螂的卵巢发育间性型和败育型个体中,又有少数卵母期桂建芳等卵巢发育阻滞的人工三倍体鱼减数分裂染色体配对的光镜观察细胞能继续发育生长到卵黄生成和成熟阶段呢这些发育生长的卵母细胞其三倍体染色体又是如何配对并能突破这种配对联会所造成紊乱局面的呢最近,等从对虹缚.
口口,人工三倍体雄鱼的研究中揭示出三倍体鱼减数分裂的染色体配对和分离的一种新情况.
他们鉴定分析了由三倍体雄性虹缚的精子与正常二倍体雌鱼的卵子受精发育而成的胚胎的染色体,发现这些胚胎大多数含有一个染色体,即个染色体来源于卵子一,一个染色体来源于三倍体雄鱼一的精子.
依据这一发现,等认为,在精子发生期间,三倍体虹鳍的染色体可能是通过类似于普通减数分裂的方式分为两组的〔.
虽然这仅仅是一种推测,但由此可见,三倍体的减数分裂染色体配对、联会和分离等行为是相当复杂的,这种复杂性从天然多倍体物种的配子发生机制和生殖方式〔'也不难看到.
那些卵巢发育正常形成了成熟卵子的人工三倍体鱼的卵子发生机制是否同于天然三倍体鱼呢很明显,这是非常值得探究的.
卵巢发育阻滞型三倍体鱼粗线期细胞的另一显著特征是不同细胞之间染色体结构的粗细和长短的大小变化.
这种变化一方面可能是初级卵母细胞发育程度差异上的反映,其染色体结构的粗细可能主要与联会过程中产生的特异蛋白〔'"的多少有关.
从部分配对二价体的已配对区域染色较强、未配对区域染色较弱图版和着丝粒区域相对较细染色较弱图版的结果来看,也可明显看出配对过程中特异性蛋白的产生及其产生的结构差异性.
染色体大小变化的另一方面可能正如不同个体间卵巢发育状态表现出多种类型的多样性一样,同一个体中不同细胞的发育能力也可能不同,这也许正是三倍体鱼及其三倍体细胞的复杂之处.
显然,这种复杂性仅用染色体配对紊乱是解释不了的,它必定牵涉到基因表达和调控的许多基本问题.
减数分裂的染色体配对是一个多步骤的复杂过程,它包括同源染色体的识别和排列、联会复合体的形成以及允许发生交换的同源顺序的准确配对.
由于染色体配对始于细线期和偶线期,但这两个时期的期限较短,且不利于观察,因而在普通二倍体物种中很难观察到染色体配对的详细过程.
从作者在人工三倍体水晶彩螂卵巢发育阻滞型雌鱼的减数分裂粗线期细胞中所观察到的正在配对过程中的配对叉以及三价体及多价体的构型来看,它们之中仍保留有配对和可能联会的起始状态.
因此,人工三倍体鱼还可作为进行减数分裂染色体配对和联会机制研究的理想材料.
这在植物三倍体中已有一些报道,'',.
本研究虽然观察到一些现象,但由于光镜的分辨率所限,许多细微的结构还有待于电镜的观察和分析.
参考文献斗,,,'脚,,一,'!
,!
,一,,,!
##%&()+,).
)/,0)1%(),1986,115:84一848.
桂建芳、梁绍昌、孙建民、黄文郁、蒋一佳.
鱼类染色体组操作的研究I·静水压休克诱导三倍体水晶彩螂.
水生生物学报,1990,14(4):336一344.
桂建芳、肖武汉、陈丽、梁绍昌、蒋一佳.
人工三倍体水晶彩螂的性腺发育.
动物学报,1991,37(3):297一304.
FleteherJM.
,Lighrmieroseopeanalysisofmeioticprophasechromosomebysilverstaining.
Chromosoma,1979,72:241一248.
马昆、施立明、俞秀璋、王思凯、孙宝琪.
东北马鹿和东北梅花鹿Fl_杂种细胞联会复合体分析.
遗传学报,1988,水生生物学报一9卷15(3):197一200.
fs]HowellwM,BlaekDA,Controlledsilver一stainingofnue]eousorganizerregionswirhapro-tectiveeolloidaldeveloPer:al一stePmethod.
ExPerientia,1980,36:10一4一1015.
[9]UedaT,sauadaM,KobayashiJ,CytogeneriealcharaeteristicsoftheembryosbetweendiploidfemaleandtriPloidmaleinrainbow:rour.
JaP.
J.
Gener.
,1987,62(5):461一466.
「10]桂建芳.
单性脊椎动物的进化遗传学.
自然杂志,1989,一2(2):116一21.
皿l]吴清江、叶玉珍、陈荣德.
铿鱼染色体人工调控的核型证明.
水生生物学报,1986,10(2):101一108.
汇12]S:ernH,HottaY.
,Chromosomeorganizationintheregulation.
In:Evans,C.
W.
01、dDieki-nson,H.
G.
(Editors),ControllingEvenrsinMeiosis.
TheCompanyofBiologistsLtd,1984,pp.
161一175.
厂13]LoidlJ,JonesGH.
,synaptonemaleomplexspreadinginAlliu,.
1.
TriploidA.
sPhaorocePhlo,.
Chromosoma,1986,93:420一428.
f14]whiteJ,JenkinsG,parkerJ5.
,Eliminationofmultivalentsduringmeiotieprophaseinscilla口ur"仍"a115.
1.
DiploidandtriPloid.
Genome,1988,30:930一939.
LIGHTMICROSCOPEINVESTIGATIONONMEIOTICCHROMOSOMEPAIRINGINARTIFICIALTRIPLOIDFISHSVVITHBLOCKEDOVARIESGuiJianfang,ChenLi,LiangShaochangand些巡止迎匕(IoszfzuzoofHydrobiology,T人.
Chi,e,eAcadem夕Sci.
,cos,W二han430072)AbstraetChromosomepairingwaslightmicroscopieallyinvestigatedbyapvlyingsurfaeesPreadingtechniqueineombinationwithl一stePsilverstainingmethodforthePa-ehyteneehromosomeofartifieialtriploidtransparenteoloredcruciancarp(CarassiusauratustransParenteoloredvariety)withbloekedovaries.
ThetriPloidfisheswereProdueedfromhydrostatiePressureshoekforfertilizedeggsbyindueingseeondPo-larbodyretention.
Thestudywasperformedonthetriploidfemaleswithbloekedovaries,oneofmanydifferenttyPesoffemaleswithvariousdeveloPingovaries.
Twosubtypesweredistinguishedaccordingtothepresenceorabsenceofprimaryoocytes.
MeiotieProPhasecellswerenotobservedinthetriPloidfemaleswithoutPri-maryoocytes.
InthetriploidfemaleswithblockedovariescontainingdifferentiatedPrimaryooeytes,thePaehyteneehromosomesweremainlyeomPosedofbivalentsandunsynaPsedunivalents,andafewoftrivalentsandothermultivalentswerealsofound.
Thereareabout90ehromosomeelementsincludingunivalents,bivalents,tri-valentsandothermultivalentsinthepachyteneeells.
Thesizeofehromosomesamongdiffrentpaehyteneeellswasobviouslyvarious.
Thepairingforksandspeei-fiesynaptonemalproteinswereelearlyobservedatsomePairingandPairedehromo-somes.
Therelationshipbetweendisturbedehromosomepairingandblockedooeytedevelopinghasbeenbrieflydiseussed.
Theinvestigationalsosuggestedthatthearti-ficialtriploidfishesmightbeexeellentmaterialsforstudyingthemechanismofchromosomepairingandsynaPsing.
KeywordsArtificialtriploidfish,Ovarydevelopment,Meiosis,Chromosomepairing
,,卵巢发育阻滞的人工三倍体鱼减数分裂染色体配对的光镜观察桂建芳陈丽梁绍昌蒋一建中国科学院水生生物研究所,武汉.
.
提要采用界面铺张制片和硝酸银一步染色的方法,对人工三倍体水晶彩螂卵巢发育阻滞型个体的减数分裂染色体配对进行了光镜观察.
在分化有初级卵母细胞的卵巢发育阻滞型的三倍体鱼中,减数分裂粗线期细胞主要由二价体和单价体组成,也见有少量三价体和其它多价体,其染色体成员数大多在左右在不同细胞间,染色体的大小变化较大配对联会过程中形成的配对又和产生的特异蛋白在一些细胞中明显可见.
文中讨论了三倍体染色体配对紊乱与卵母细胞发育受阻的关系,并认为人工三倍体鱼是进行减数分裂染色体配对和联会机理研究的好材料.
关键词人工三倍体鱼,卵巢发育,减数分裂,染色体配对人工三倍体鱼大多数是不育的【'·",其不育的主要原因一般只是依据植物三倍体中的观察,认为是紊乱的减数分裂染色体配对和分离所致〔,而有关人工三倍体鱼减数分裂染色体行为的研究迄今还未见报道.
本研究是在作者采用静水压休克批量生产出三倍体水晶彩螂〔〕,并在雌性三倍体鱼中发现有多种卵巢发育类型,可明显区分为阻滞型、间性型、败育型和极个别正常型「习的基础上,采用界面铺张法制片和硝酸银快速一步银染的方法,首先对人工三倍体水晶彩螂卵巢发育阻滞型个体的减数分裂前期染色体配对等行为进行了光镜的初步观察.
材料与方法材料人工三倍体水晶彩螂是在卵子受精后采用耐的静水压持续处理诱导而成.
用作制片观察的卵巢发育阻滞型个体共尾,制片同时分别进行了组织切片的细胞学观察.
根据形态和组织学观察,卵巢发育阻滞型的三倍体雌鱼又可分为两个亚类,一类卵巢呈细线状,仅含有卵原细胞另一类呈线状,已分化有较多的初级卵母细胞〔.
用作制片观察的前一类鱼为,尾,后一类鱼为尾.
作为对照,还进行了尾雄性二倍体水晶彩螂精巢细胞的制片.
界面铺张法制片界面铺张法制片主要参考和马昆等叨报道的方法进本工作得到中国科学院青年科学基金和院长基金的资助.
武汉大学生物学系届毕业实习生.
年月日收到一年月修回.
水生生物学报一卷行.
首先剥净卵巢或精巢上的脂肪组织,取组织块洗净,于.
培养液中制成细胞悬液,静置左右,取上层细胞悬液经离心一次后,加少许生理盐水制成细胞悬液.
然后在一涂有石蜡的平皿上滴加一直径约的低渗液液珠,取少量细胞悬液与液珠表面接触,细胞即在液珠表面铺张.
约后,用一个干净的玻片与载有细胞的液珠表面接触,细胞即附着于玻片上.
平置一,将玻片置于新配制的并多聚甲醛一中固定,取出凉干.
硝酸银染色凉千的玻片按和报道的一步银染法进行硝酸银染色.
光镜观察.
结果在卵巢发育严重阻滞的第一亚类的尾三倍体鱼中,正如组织切片的细胞学观察很难找到初级卵母细胞一样,其染色体制片上也没有见到减数分裂的前期细胞.
在已分化有较多初级卵母细胞的第二亚类发育阻滞型三倍体鱼的卵巢细胞染色体制片上,均可见到一定比例的减数分裂粗线期细胞.
其共同的特征之一是染色体配对紊乱.
在同一细胞的染色体扩散相图版、和中,总可见到部分染色体染色较浅,呈细丝状,相互缠绕严重,难于区分,为没有配对的单价体大部分染色体已配对,染色较浓、较粗,相互之间易区分,主要为二价体,少数可明显看出是多价体.
在多价体中,有已配对的二价体再与一价体排列,并在一些点和区段发生配对的三价体图版有由两条已联会的二价体通过端粒联合形成的四价体图版也有由多条非同源染色体相互交叉折叠配对形成的复合多价体图版.
在一些尚未联会完成的二价体上,配对叉清晰可见图版.
在一些已完成联会的染色体上,着丝粒区域与其它区段的结构差异相当显著,着丝粒区域细且着色浅,与粗且着色深的其它区段形成强烈的对比图版、.
多数细胞中的单价体、二价体和多价体的总染色体成员数一般在左右图版一.
与这些三倍体鱼相比,由二倍体鱼精巢细胞制备的粗线期染色体比较清晰,染色体配对正常,粗细一致,条二价体清晰可数图版.
此外,粗线期染色体的另一个显著特征是其细胞相互之间在结构和大小上的变化图版、、和,它们既表现有染色体的粗细之分,又表现有染色体的长短不同.
讨论三倍体水晶彩鲡含有三套染色体组,共个染色体.
从理论上推测,在同源染色体配对及联会过程中,有可能出现下列三种情况一是形成条二价体,条单价体二是形成条三价体三是单价体、二价体、三价体甚至更高的多价体同时并存.
从卵巢发育阻滞型三倍体鱼粗线期染色体配对图象来看,与第三种情形基本相似,主要由二价体和单价体组成,也含有少量三价体和其它多价体,其染色体成员数一般在左右.
已配对联会了的染色体发生了较大程度的凝缩,而未配对的单价体仍呈丝状,相互缠绕.
从这种染色体结构上的显著差异所造成的紊乱现象来看,它必定影响了细胞的继续发育和生长,导致发育阻滞.
然而,同样是人工三倍体鱼,为什么少数个体又能正常发育形成成熟的卵子呢为什么在人工三倍体水晶彩螂的卵巢发育间性型和败育型个体中,又有少数卵母期桂建芳等卵巢发育阻滞的人工三倍体鱼减数分裂染色体配对的光镜观察细胞能继续发育生长到卵黄生成和成熟阶段呢这些发育生长的卵母细胞其三倍体染色体又是如何配对并能突破这种配对联会所造成紊乱局面的呢最近,等从对虹缚.
口口,人工三倍体雄鱼的研究中揭示出三倍体鱼减数分裂的染色体配对和分离的一种新情况.
他们鉴定分析了由三倍体雄性虹缚的精子与正常二倍体雌鱼的卵子受精发育而成的胚胎的染色体,发现这些胚胎大多数含有一个染色体,即个染色体来源于卵子一,一个染色体来源于三倍体雄鱼一的精子.
依据这一发现,等认为,在精子发生期间,三倍体虹鳍的染色体可能是通过类似于普通减数分裂的方式分为两组的〔.
虽然这仅仅是一种推测,但由此可见,三倍体的减数分裂染色体配对、联会和分离等行为是相当复杂的,这种复杂性从天然多倍体物种的配子发生机制和生殖方式〔'也不难看到.
那些卵巢发育正常形成了成熟卵子的人工三倍体鱼的卵子发生机制是否同于天然三倍体鱼呢很明显,这是非常值得探究的.
卵巢发育阻滞型三倍体鱼粗线期细胞的另一显著特征是不同细胞之间染色体结构的粗细和长短的大小变化.
这种变化一方面可能是初级卵母细胞发育程度差异上的反映,其染色体结构的粗细可能主要与联会过程中产生的特异蛋白〔'"的多少有关.
从部分配对二价体的已配对区域染色较强、未配对区域染色较弱图版和着丝粒区域相对较细染色较弱图版的结果来看,也可明显看出配对过程中特异性蛋白的产生及其产生的结构差异性.
染色体大小变化的另一方面可能正如不同个体间卵巢发育状态表现出多种类型的多样性一样,同一个体中不同细胞的发育能力也可能不同,这也许正是三倍体鱼及其三倍体细胞的复杂之处.
显然,这种复杂性仅用染色体配对紊乱是解释不了的,它必定牵涉到基因表达和调控的许多基本问题.
减数分裂的染色体配对是一个多步骤的复杂过程,它包括同源染色体的识别和排列、联会复合体的形成以及允许发生交换的同源顺序的准确配对.
由于染色体配对始于细线期和偶线期,但这两个时期的期限较短,且不利于观察,因而在普通二倍体物种中很难观察到染色体配对的详细过程.
从作者在人工三倍体水晶彩螂卵巢发育阻滞型雌鱼的减数分裂粗线期细胞中所观察到的正在配对过程中的配对叉以及三价体及多价体的构型来看,它们之中仍保留有配对和可能联会的起始状态.
因此,人工三倍体鱼还可作为进行减数分裂染色体配对和联会机制研究的理想材料.
这在植物三倍体中已有一些报道,'',.
本研究虽然观察到一些现象,但由于光镜的分辨率所限,许多细微的结构还有待于电镜的观察和分析.
参考文献斗,,,'脚,,一,'!
,!
,一,,,!
##%&()+,).
)/,0)1%(),1986,115:84一848.
桂建芳、梁绍昌、孙建民、黄文郁、蒋一佳.
鱼类染色体组操作的研究I·静水压休克诱导三倍体水晶彩螂.
水生生物学报,1990,14(4):336一344.
桂建芳、肖武汉、陈丽、梁绍昌、蒋一佳.
人工三倍体水晶彩螂的性腺发育.
动物学报,1991,37(3):297一304.
FleteherJM.
,Lighrmieroseopeanalysisofmeioticprophasechromosomebysilverstaining.
Chromosoma,1979,72:241一248.
马昆、施立明、俞秀璋、王思凯、孙宝琪.
东北马鹿和东北梅花鹿Fl_杂种细胞联会复合体分析.
遗传学报,1988,水生生物学报一9卷15(3):197一200.
fs]HowellwM,BlaekDA,Controlledsilver一stainingofnue]eousorganizerregionswirhapro-tectiveeolloidaldeveloPer:al一stePmethod.
ExPerientia,1980,36:10一4一1015.
[9]UedaT,sauadaM,KobayashiJ,CytogeneriealcharaeteristicsoftheembryosbetweendiploidfemaleandtriPloidmaleinrainbow:rour.
JaP.
J.
Gener.
,1987,62(5):461一466.
「10]桂建芳.
单性脊椎动物的进化遗传学.
自然杂志,1989,一2(2):116一21.
皿l]吴清江、叶玉珍、陈荣德.
铿鱼染色体人工调控的核型证明.
水生生物学报,1986,10(2):101一108.
汇12]S:ernH,HottaY.
,Chromosomeorganizationintheregulation.
In:Evans,C.
W.
01、dDieki-nson,H.
G.
(Editors),ControllingEvenrsinMeiosis.
TheCompanyofBiologistsLtd,1984,pp.
161一175.
厂13]LoidlJ,JonesGH.
,synaptonemaleomplexspreadinginAlliu,.
1.
TriploidA.
sPhaorocePhlo,.
Chromosoma,1986,93:420一428.
f14]whiteJ,JenkinsG,parkerJ5.
,Eliminationofmultivalentsduringmeiotieprophaseinscilla口ur"仍"a115.
1.
DiploidandtriPloid.
Genome,1988,30:930一939.
LIGHTMICROSCOPEINVESTIGATIONONMEIOTICCHROMOSOMEPAIRINGINARTIFICIALTRIPLOIDFISHSVVITHBLOCKEDOVARIESGuiJianfang,ChenLi,LiangShaochangand些巡止迎匕(IoszfzuzoofHydrobiology,T人.
Chi,e,eAcadem夕Sci.
,cos,W二han430072)AbstraetChromosomepairingwaslightmicroscopieallyinvestigatedbyapvlyingsurfaeesPreadingtechniqueineombinationwithl一stePsilverstainingmethodforthePa-ehyteneehromosomeofartifieialtriploidtransparenteoloredcruciancarp(CarassiusauratustransParenteoloredvariety)withbloekedovaries.
ThetriPloidfisheswereProdueedfromhydrostatiePressureshoekforfertilizedeggsbyindueingseeondPo-larbodyretention.
Thestudywasperformedonthetriploidfemaleswithbloekedovaries,oneofmanydifferenttyPesoffemaleswithvariousdeveloPingovaries.
Twosubtypesweredistinguishedaccordingtothepresenceorabsenceofprimaryoocytes.
MeiotieProPhasecellswerenotobservedinthetriPloidfemaleswithoutPri-maryoocytes.
InthetriploidfemaleswithblockedovariescontainingdifferentiatedPrimaryooeytes,thePaehyteneehromosomesweremainlyeomPosedofbivalentsandunsynaPsedunivalents,andafewoftrivalentsandothermultivalentswerealsofound.
Thereareabout90ehromosomeelementsincludingunivalents,bivalents,tri-valentsandothermultivalentsinthepachyteneeells.
Thesizeofehromosomesamongdiffrentpaehyteneeellswasobviouslyvarious.
Thepairingforksandspeei-fiesynaptonemalproteinswereelearlyobservedatsomePairingandPairedehromo-somes.
Therelationshipbetweendisturbedehromosomepairingandblockedooeytedevelopinghasbeenbrieflydiseussed.
Theinvestigationalsosuggestedthatthearti-ficialtriploidfishesmightbeexeellentmaterialsforstudyingthemechanismofchromosomepairingandsynaPsing.
KeywordsArtificialtriploidfish,Ovarydevelopment,Meiosis,Chromosomepairing
CheapWindowsVPS$4.5/月,美国VPS/免费Windows系统/1Gbps不限流量/,可选美洲、欧洲、亚洲等8大机房
国外商家提供Windows系统的并不常见,CheapWindowsVPS 此次提供的 2 款 VPS 促销套餐,提供 5 折永久优惠码,优惠后月付 4.5 美元起,价格还是挺诱人的,VPS 不限流量,接入 1Gbps 带宽,8 个机房皆可选,其中洛杉矶机房还提供亚洲优化网络供选择,操作系统有 Windows 10 专业版、2012 R2、2016、Linux等。Cheap Windows VPS是...
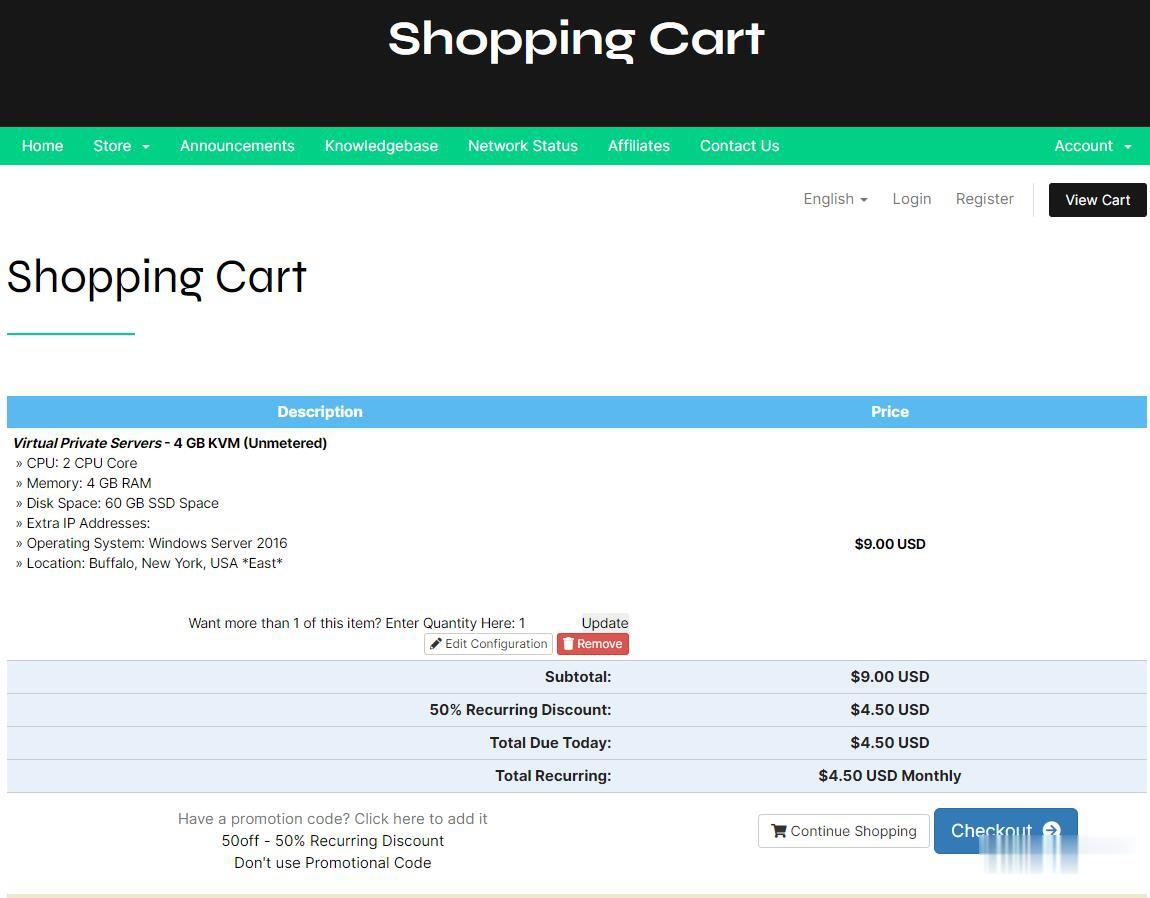
打开海外主机域名商出现"Attention Required"原因和解决
最近发现一个比较怪异的事情,在访问和登录大部分国外主机商和域名商的时候都需要二次验证。常见的就是需要我们勾选判断是不是真人。以及比如在刚才要访问Namecheap检查前几天送给网友域名的账户域名是否转出的,再次登录网站的时候又需要人机验证。这里有看到"Attention Required"的提示。我们只能手工选择按钮,然后根据验证码进行选择合适的标记。这次我要选择的是船的标识,每次需要选择三个,一...
HostKvm新上联通CUVIP线路VPS,八折优惠后1G内存套餐$5.2/月起
最近上洛杉矶机房联通CUVIP线路主机的商家越来越多了,HostKvm也发来了新节点上线的邮件,适用全场8折优惠码,基于KVM架构,优惠后最低月付5.2美元起。HostKvm是一家成立于2013年的国人主机商,提供基于KVM架构的VPS主机,可选数据中心包括日本、新加坡、韩国、美国、中国香港等多个地区机房,君选择国内直连或优化线路,延迟较低,适合建站或者远程办公等。以洛杉矶CUVIP线路主机为例,...
静水论坛为你推荐
-
软银赛富石家庄哪家少儿英语是连锁学校,教的好信得过?轿车和suv哪个好同价位的轿车和suv哪个更好?网页传奇哪个好玩求最好玩的网页传奇?手机杀毒哪个好手机杀毒软件哪个好 五大手机安全软件杀毒大比拼51空间登录51空间怎么进辽宁联通网上营业厅网辽宁省联通发短信查gprs流量方法qq网盘在哪里QQ网盘在哪??????首选dns服务器地址首选DNS服务器是什么意思dns服务器什么意思DNS服务器是什么意思啊?是指路由器吗?360云盘360云盘有多大?