unbound7788kk.com
7788kk.com 时间:2021-03-21 阅读:()
INFECTIOUSDISEASE(MSPASTERNACK,SECTIONEDITOR)HostandViralFactorsinRespiratorySyncytialVirusInfectionPeterMastrangeloRichardG.
HegelePublishedonline:15May2013SpringerScience+BusinessMediaNewYork2013AbstractRespiratorysyncytialvirus(RSV)isamajorworldwidepathogenforwhichthereisstillnoeffectivevaccineorantiviraltreatmentavailable,andimmunopro-phylaxiswithRSV-specicantibodies(e.
g.
,palivizumab)isusedinlimitedclinicalsettings.
Inthisreview,wedis-cussvirus–hostinteractionsrelevanttoRSVpathobiologyandhowadvancesincellandsystemsbiologyhaveacceleratedknowledgeinthisarea.
WealsohighlightrecentadvancesinunderstandingtherelationshipbetweenRSVbronchiolitisandsequelaeofrecurrentwheezingandasthma,newndingsintoanintriguinginteractionbetweenRSVandairpollution,andexcitingdevelopmentstowardthegoalofrealizingasafeandeffectiveRSVvaccine.
KeywordsRespiratorysyncytialvirusViralpathogenesisVirus–hostinteractionsViruspersistenceAsthmaAllergySystemsbiologyNucleolinAirpollutionVaccinologyIntroductionRespiratorysyncytialvirus(RSV)isasingle-stranded,negative-polarityPneumovirusthatinfectspeopleofallagesworldwide[1].
RSVisafrequentcauseofupperrespiratorytractinfectionsandotitismedia,andalsocausesseriouslowerrespiratorytractinfectionssuchasbronchi-olitisandpneumonia,particularlyininfantsandyoungchildren,immunocompromisedindividuals,andtheelderly.
RSVinfectionsinearlypostnatallifehavealsobeenassociatedwiththeonsetofallergicsensitizationandrecurrentwheezing/asthma.
Despitealmost60yearshav-ingpassedsincethediscoveryofRSV,thereisstillnolicensedvaccineavailable,andprophylaxiswithimmu-noglobulin(palivizumab)isusedonlyforasubsetof''high-risk''children(e.
g.
,pretermbirths,andinfantswithcongenitalheartdisease,congenitalimmunodeciency,and/orbronchopulmonarydysplasia).
AlthoughanumberofpharmacologicalagentshavebeenusedinRSVtherapy,theirefcacyandeffectivenessarepoor[2].
Here,wewillbrieyreviewsomeviralandhostfactorsimplicatedinthepathobiologyofRSVinfections,focusingonhowthediversityofthesefactorsmightberelatedtothevariabilityseeninclinicaldisease.
OurdiscussionwillalsoincludecomparingandcontrastingcharacteristicsofRSVinfectionsinvitroversusinvivo;currentperspectivesontherelationshipbetweenRSVinfections,pediatricallergy,andasthma;recentworkthatdescribesanovelrelationshipbetweenRSVandparticulatematter(airpollution);andprospectsforthedevelopmentandimplementationofasafe,effectiveRSVvaccine.
ViralandHostFactorsinRSVInfection:TowardanIntegratedPerspectiveRSVStructureHumanRSVisalamentousenvelopedvirusinwhichtheviralnucleicacidandstructuralandinternalproteinsaresurroundedbyaproteinaceousoutercoat[3].
AsanP.
MastrangeloR.
G.
Hegele(&)DepartmentofLaboratoryMedicineandPathobiology,UniversityofToronto,6231-1King'sCollegeCircle,Toronto,ONM5S1A8,Canadae-mail:richard.
hegele@utoronto.
caR.
G.
HegeleDepartmentofPaediatricLaboratoryMedicine,TheHospitalforSickChildren,Toronto,ON,Canada123CurrPediatrRep(2013)1:149–157DOI10.
1007/s40124-013-0019-3envelopedvirus,RSV'sgeneralstructurehassimilaritiestothatofinuenzaandparainuenzaviruses,metapneum-oviruses,andcoronaviruses,andcontrastswiththatofothercommonrespiratorypathogenssuchasrhinovirusesandadenoviruses,whicharenonenveloped.
RSVhasthreeenvelopeproteins:G(glycoprotein),F(fusion),andSH(smallhydrophobic)[1].
HeterogeneityintheGglyco-proteindenesthetwomainsubgroupsofRSV,AandB[4].
SomeliteraturesuggeststhatRSVAstrainsproducemoreclinicallysevereinfectionsthanRSVBstrains[5],butthiscontentionhasprovendifculttoreproduceamongdifferentgroupsofinvestigators[6].
OnestrainofRSVA,so-calledline19[7],appearstobeofparticularinterestregardingmucusproductionandsecretionofIL-13,aTh2cytokineassociatedwithallergic-typehostresponses(seebelow),andtherebycontributinginparttotheheteroge-neityandseverityofclinicalmanifestations.
Inaddition,RSVGhasasecretedformthatbindshostantibodiesandtherebyservesasanimmunological''decoy''thatfacili-tatescontinuedviralreplication[8].
RSVGisalsocon-sideredtobeakeyattachmentproteinforthevirustobindtothecellsurfacetoinitiateinfection;however,withthediscoveryandengineeringofRSVstrainsdecientinG(so-calledRSVDG)thatareinfectious,albeitatwithmuchlowerefciencythanvirusexpressingG,RSVGisneithersufcientnornecessarytocauseinfection[9].
Bycontrast,RSVFiscrucialforfusionoftheviralenvelopetothehostcellsurface,sinceRSVDFcannotentercellsonitsownbutratherrequiresahelpervirustoinitiateinfection[10].
Onfreevirus,RSVFexistsina''prefusion''formthatundergoesanelegantlycharacterizedconforma-tionalchangeduringthefusionprocess[11,12].
IncontrasttoRSVG,RSVFishighlyconservedbetweenthetwomajorsubgroupsofthevirus[13].
RSVFandRSVGmayhaveanoverall''yin-yang''effectonhostadaptiveimmuneresponsesconcerningTh1–Th2balance.
Inbrief,Th1responsesinvolveasubsetofCD4lymphocytesproducingcytokinessuchasinterferon(IFN)-candinterleukin(IL)-2consideredimportantindelayedhypersensitivityandanti-viralimmunity,whereasTh2responsesinvolveproductionofinammatorymediatorssuchasIL-4,IL-5,andIL-13byadifferentsubsetofCD4lymphocytesandareimplicatedinallergy.
RSVFappearstostimulateTh1cells,whereasRSVGappearstostimulateTh2responses,andclinicalmanifestationsarebelievedtoreectwhethertheoverallresponseisshiftedtowardTh1orTh2,orremainsinbalancewithregardtocytokineproduction[14].
TheRSVSHprotein,byanalogytosimilarproteinsexpressedinotherviruses,mayhavearoleinenhancingthepermeabilityofthecellsurfacetomoleculesandions[15].
TheimportanceofthisputativefunctionofRSVSHisunclear,sincegeneticallyengineeredstrainsofRSVinwhichSHisdeletedappeartobeessentiallyasefcientaswild-typevirusincausingcellularinfections[16].
ThereisincreasinginterestinSHasapotentialviralantigentargetinvaccinedevelopment(seelater).
OtherRSVcompo-nents,includingnucleocapsid(N),polymerase(L),non-structuralcomponents(NS1,NS2),phosphoprotein(P),andmatrix(M,M2-1,M2-2)havebeenimplicatedinstimulatinghostantiviralresponses(e.
g.
,IFN-aandIFN-b)througheffectsinhostcellsignaltransductionpathwaysandimportantnucleartranscriptionfactors(e.
g.
,NF-jB,AP-1).
FurtherdetailsoftheroleofnonenvelopedproteinsinRSVreplicationandhostresponsesareprovidedinanexcellentreviewbyCollinsandMelero[1].
RSVInfectionsInVitroVersusInVivoOneofthebiggestenigmassurroundingRSVpathobiologyisthestrongtropismthevirusshowsforinfectingthehumanrespiratorytractinvivo;invitro,RSVestablishesproductiveinfectionofvirtuallyallmammaliancelltypesstudied,includingawidevarietyofprimaryandcontinuouscellslinesofepithelialcells[17],macrophages[18],otherleukocytes[19],broblasts[20],andevenneurons[21].
Infact,ndingacelltypethatisresistanttoRSVinfectioninvitroischallenging.
Aswillbediscussedin''NucleolinasaFusionReceptorforRSV:FromDiscoverytoDrugTargeting''inthecontextofthediscoveryofacellularfusionreceptorforRSV,Sf9cellsfromthemothSpo-dopterafrugiperdaaredifculttoinfectwithRSV[22].
InadenitiveautopsystudyofthelungpathologyofuntreatedacuteRSVinfections,Johnsonetal.
[23]repor-tedthatRSVantigenswereobservedwithinairwayandalveolarepithelialcells,aswellaspulmonarymacro-phages.
Similarobservationshavebeenmadeinexperi-mentalanimals.
InaguineapigmodelofexperimentalRSVinfection,ourgrouphasdocumentedtheexpressionofviralantigensviaimmunohistochemistrywithinlungepithelialcells,alveolarmacrophages,interstitialcells(presumablydendriticcells),andcellswithinbronchus-associatedlymphoidtissue,foratleast100daysafterRSVinoculation[24].
ArecentstudyhasreportedthatRSVcanbedetectedinapproximately15%ofhumantonsilsofchildrenwithchronicadenotonsillardisease[25].
GiventheabundantliteratureconcerningtheeffectsofRSVinfectiononthesecretionofvariousinammatorymedia-tors,responsesofcellsignalingpathways,andotherhostresponsesobservedinvitro,onemustexercisecautioninextrapolatingthesignicanceofobservationsmadeinvitrototheinvivostate,andwhethertheyrepresentopportu-nitiesforpotentialclinicalinterventions.
OneonlyneedstoreectonthehistoricallydisappointingresultsseenwithattemptstotreatRSVinfectionsofexperimentalanimalsorpatientswithantiviraloranti-inammatoryagentstoappreciatethispoint[2,26].
150CurrPediatrRep(2013)1:149–157123Incontrasttoothervirusessuchasinuenzavirusandadenovirus,whereextensivecytotoxicityandcelldeathisthenormwithinfectioninvitro,RSVinfectiondoesnotelicitmuchinthewayofcelldamage[17].
Thereisevi-dencethatRSVcanactuallydecreasetherateofapoptosisinvitro[27].
Takenfurther,RSVhasclearlybeenshowntocausepersistent,noncytolyticinfectionofepithelialcellsinvitro[28].
RSVpersistence,characterizedbycontinu-ous,low-levelreplicationofinfectiousvirus,hasbeendescribedinvivo,rstinamodelinwhichcottonratswereimmunosuppressedwithcyclophosphamide[29],andsub-sequentlyinimmunocompetentguineapigs[30]andinmice[31,32].
ThereisalsoevidenceconsistentwithRSVpersistenceinhumans,bestcharacterizedinthesettingofadultpatientswithchronicobstructivepulmonarydisease[33].
ApossibleroleforRSVpersistenceinthepathobi-ologyofpostbronchiolitiswheezing/asthmaisdescribedin''RSVandPediatricAsthma:HaveWeFoundtheMissingLink''RSV–HostInteractions:MovingEverMoreProximallyFormanyyears,investigationofRSV–hostinteractionsfocusedoncharacterizingtheinammatorymediatorresponsesofinfectedcells,animals,andhumansubjectswiththehopethatanunderstandingofthevirus-specichostresponsestoRSVinfectionscouldinformpossibleinterventionsdesignedtomodulaterelevantinammatorymediatorsandtherebyleadtoimprovedclinicaloutcomes[34].
Theapparentheterogeneityinvirus-specicimmu-nityobservedinhostadaptiveimmuneresponsestoRSV(bothcell-mediatedandhumoral),dependingonthepop-ulationstudied,complicatestheabilitytomakeclinicallyrelevantadvances.
ExtensiveinquirieshavefocusedoninnateimmunemechanismsintheearlyhostresponsetoRSV[35,36].
Onerationaleforthisapproachisthatbyunderstandingtheearly,''proximal''eventsthatoccurinRSVinfections,itmaybeeasiertodevelopeffectiveclinicalinterventionsthatcouldbeusedeffectivelyamongdiversehostswhohavedifferentadaptiveimmuneresponsestothevirus[15,37].
Thisrationalehasbeenrecentlyquestioned,sinceapparentdifferencesininnateimmuneresponseshavebeenobservedbetweenRSVAandRSVBinfections[38].
Nevertheless,researchintotheroleofinnateimmunemechanismsinRSVpathobiologyledtoanumberofpro-foundinsights,oneofwhichwastheabilityofRSVtoincreasethelevelofpattern-recognitionreceptors(e.
g.
,Toll-likereceptors)expressedonthecellsurface[39].
Theseobservationswereinuentialtotheeldforanumberofreasons:forexample,thenatureofcellsurfacemoleculesinvolvedininnateimmunitythatcanbeaffectedbyRSVwaspoorlyunderstood,andpattern-recognitionreceptors(e.
g.
,Toll-likereceptor4andothers)stimulatecellsig-nalingpathwaysthatareamenabletopharmacologicalmanipulation[40],eveniftheyarenotactualreceptorsforthevirus[41].
OnecellsignalingmoleculerelatedtoToll-likereceptorinteractionwithRSVisp38mitogen-activatedproteinkinase,asrevealedbyascreenofculturedcellswithapanelofpharmacologicalkinaseinhibitors.
Inhibitionofp38mitogen-activatedproteinkinasewithSB203580(acompoundoriginallydevelopedfortreatmentofarthritisandcancer)wasassociatedwithmarkedinhibitionofRSVreplicationinculturedcells,withoutinducingsignicantdrug-relatedcytotoxicity[42].
Thistypeofexperimentisanexampleofso-calleddrugrepurposing,inwhichacom-pounddevelopedforanunrelatedclinicalindicationcanbegivenonthebasisofpathobiologydenedinanotherclinicalsetting[43].
Itremainstobeseenwhethersuchanapproachtodrugrepurposingwillbeusefulintheman-agementofRSVinfectionsinvivoinanimalmodels,andultimatelyinhumans.
NucleolinasaFusionReceptorforRSV:FromDiscoverytoDrugTargetingOurgroupidentiedcellsurfacenucleolinasamoleculethatfulllsthecriteriaofafunctionalreceptorforRSV[44].
Oneofthekeychallengestoovercomeinthisdis-coverywasndingacelltyperesistanttoRSVinfectioninvitrowhichcouldbecomepermissiveafterforcedexpressionofthehumannucleolingeneandectopicexpressionofhumannucleolinproteinonthecellsurface.
ThiswasachievedbyuseofinsectSf9cells[22].
Vali-dationofnucleolinasacellularreceptorinvivowasaccomplishedinamousemodelbysmallinterferingRNAknockdownofnucleolinpriortoRSVchallenge,whichwasassociatedwithasignicantdecreaseinRSVtitersasmeasuredbyquantitativeplaqueassaysoflunghomoge-nates.
WeshowedthatRSVF,notRSVG,interactswithnucleolin,andwecoinedtheterm''fusionreceptor''todenotethisfact.
NucleolinisanespeciallychallengingpotentialdrugtargetforRSV.
Nucleolinhasaveryshorthalf-lifewhenexpressedonthecellsurface,andconstantlyundergoesrecyclingbetweenthecytoplasmicandnuclearcompart-mentsofthecell[45].
Thus,approachesthatinterferewithcellsurfacenucleolinsuchasuseofnucleolin-specicblockingantibodies,resultintheantibody–nucleolincomplexbeinginternalizedintothecellandnew(''fresh'')unboundnucleolinmovingtothesurface[46].
Thisphe-nomenoncouldexplainwhyinourantibodyneutralizationexperimentsattemptingtovalidatenucleolinasanRSVreceptorwewereabletoachieveonlyontheorderapproximately70–80%decreasedinfectionofcellsthathadbeenpreincubatedwithnucleolin-specicantibodyCurrPediatrRep(2013)1:149–157151123priortoRSVchallenge.
Secondly,nucleolinisanotori-ously''sticky''moleculethatcanbindallsortsofmole-cules,includingproteinsexpressedbyotherviruses(e.
g.
,parainuenzavirustype3,HIV),certaindrugs(e.
g.
,mid-kine,lactoferrin,pseudopeptideHB-19),andevenDNAmoieties(reviewedin[47]).
HowtotranslatethissituationintoascenariothatisRSV-specicneedstobeaddressed.
Moreover,nucleolindoesnotexistinisolationonthecellsurface;rather,itispartofamultiproteincomplexofmolecularmassofapproximately500kDa[48].
Toaddfurthercontext,intranuclearnucleolinisessentialtocellsurvivalandreplication:inhibitingnucleolininanunde-siredcellularcompartmentcouldelicitseriousoff-targeteffects[49].
Overall,theidenticationofnucleolinasafusionreceptorforRSVposesanumberofprofoundchallengestobeovercomebeforeitmayserveasabonadedrugtarget.
ARenaissanceofRSVCellBiology,andtheGrowingRoleofSystemsBiologyOneareaofincreasingresearchactivityistheintracellularcompartmentalizationofRSVconstituentsduringthevirallifecycle.
CellbiologistsinvestigatingRSVreplicationkineticsdemonstratedthatthevirusdoesnotnecessarilyfollowastereotypedreplicativecycle,butrathercanbepreferentiallydirectedintointracellularcompartmentsthatfavororinhibitreplication[50,51].
Forexample,cyto-plasmicstressgranules[52],caveolae[53],hostcytoskel-eton[54],andlipidrafts[55]havebeenreportedtointeractwithRSV,andthesearepotentialnoveltargetsforphar-macologicalintervention.
Increasingly,theuseofhigh-throughput''systemsbiology''approachesdesignedtointerrogateRSV-infectedcellsinanunbiasedmannerhasledtoidenticationofnovelcandidatemoleculesthatcouldpotentiallybetargetedpharmacologically[56,57].
SystemsbiologypermitstheapplicationofinformaticsrelatedtopathwaymodelingrelevanttoRSVinfection.
Bydeningso-calledinteractomes[58]inwhichthevirusaffectscellfunction,investigatorsaimtoachieveacomprehensiveunderstandingofRSV–hostinteractionstoinformthedevelopmentofnovelstrategiesintreatmentandprophy-laxis.
Oneofthecurrentlimitationsofinformaticsapproachesisthatthebiologicalpathwaysbeingmodelingarethemselvesincompletelycharacterized,someaningfulinterpretationswillrequireongoingiterativeactivity[59].
RSVandPediatricAsthma:HaveWeFoundtheMissingLinkWithinafewyearsofthediscoveryofRSV,inaseminalarticle,Wittigetal.
[60]reportedthatasizeablepercentage(32%)ofinfantsandyoungchildrenhospitalizedforRSVbronchiolitisdevelopedsequelaeofrecurrentwheezingandasthmasymptoms.
Thesendingshavebeenreplicatedonanumberofoccasionsoverthelastfewdecades,althoughthereissomeongoingcontroversyoverwhetherpatientscan''outgrow''theirasthmabythetimetheyreachado-lescence,orwhethertheirconditioncanpersistintoadulthood[61–63].
OnepossibilityisthatRSVbronchi-olitisisa''marker''ofchildrenwhoaregeneticallypre-disposedtodevelopasthma,asevidencedbyaclassicstudyinwhichRSV-specicIgEwasdocumentedinchildrenwhohadrecurrentwheezingasasequelaofRSVinfection[64].
Further,inahighlyinuentialarticle,Fricketal.
[65]reportedatemporalrelationshipbetweenviralinfection(particularlyRSVandparainuenzavirus)andtriggeringallergicsensitizationin14childrenborninto''high-risk''families(i.
e.
,whererst-degreerelativeshadaprovenhistoryofallergy).
Thislatterassociationhasalsobeenconrmedinlargerstudies,includingtheCanadianAsthmaPrimaryPreventionStudyofapproximately400''high-risk''childrenandtheirfamilies[66].
Toexplorepotentialmechanismsofpostbronchiolitiswheezing/asthmaandallergy,ourgroupdevelopedagui-neapigmodelofexperimentalRSVlunginfection[67].
Guineapigswerechosenbecausetheywerealreadyamodelofhumanparainuenzavirustype3infection[68],andcouldreadilybesensitizedtoallergen(aerosolizedovalbumin)[69].
AfterinitialworkestablishingthatRSVproteinandgenomecouldpersistintheguineapiglungforupleast60daysafterRSVinoculation[70],subsequentstudiesshowedthatoutbredCamHartleyguineapigscouldharborreplicatingvirusforatleast60–100daysafterinoculation,inassociationwithnonspecicairwayhyper-responsiveness(AHR)andchronicairwayinammation(physiologicalandpathologicalhallmarksofasthma)[24,30].
However,insubsequentwork,wenotedthatviralpersistenceonitsownisnotsufcienttoproduceasthma-likechangesinthelungs,asinbred,''allergy-resistant''strain2guineapigscanalsodeveloppersistentRSVlunginfectionsbuttheseanimalsdonotgetAHRorchronicairwayinammationinassociationwithviruspersistence[71].
ConcerningthepossibilitythatRSVinfectionisa''marker''ofgeneticpredispositiontoallergy,weobservedthatRSVinfectionofCamHartleyguineapigsenhancedallergicsensitizationtoaerosolizedovalbumin,consistentwiththishypothesis[72].
Interestingly,theextentofchronicAHR/airwayinammationandvirus-enhancedovalbuminsensitizationcouldbeamelioratedbyimmu-noprophylaxisofanimalswithCpG-oligodinucleotides(CpG-ODN),apotentTh1stimulus[73].
Takentogether,theseresultssuggestthatthecombinationofRSVandaeroallergenproducesadditiveeffectsonAHRandairway152CurrPediatrRep(2013)1:149–157123inammationinan''allergy-susceptible''hostandthatCpG-ODNimmunoprophylaxisappearstotargettheRSV-associated,nottheallergen-associated,component.
Alargenumberofhumanstudieshavecharacterizeddifferencesinthetypesofcytokinesandotherchemicalmediatorsofinammation—andtheirpotentialgeneticdeterminants—indifferentsubsetsofRSV-infectedpatients(reviewedinZengetal.
[74]).
ThislargebodyofworkprovidesvaluableinsightsintopossibledifferencesinhostresponsestoRSVinfectionandmayberelevanttoexplainingwhysomeinfectedindividualsdevelopmild,upperrespiratorytractinfectionswhereasotherindividualsdevelopbronchiolitisorpneumonia,orgoontodeveloprecurrentwheezing/asthma.
Unfortunately,therehasbeenarecurrentdifcultyinvalidatingthelargenumberofcan-didatemolecules/genesbyreplicationinothercohorts,andtheoverallpictureisfarfromclear[1].
Theuseofasys-temsbiologyapproach(see''ARenaissanceofRSVCellBiology,andtheGrowingRoleofSystemsBiology'')promisestoaccelerateknowledgeofidenticationofcan-didatemoleculesandgenesforvalidation.
Onerecentstudy[75]hasreportedanexcitingdevelopmentthatmaypro-videacluetothepossiblebiological''missinglink''betweensevereRSVinfectionsandthesubsequentdevel-opmentofpediatricasthma.
Inaseriesofexperimentsthatbeganwithunbiasedinterrogationofcellstoidentifycan-didategenes,followedbyvalidationinexperimentalani-malsandanalysisofhumancohorts,investigatorshavecharacterizedgeneticpolymorphismsinthepromoterregiontoNFKBIAashavingbothfunctionalandclinicalsignicanceinthecontextofRSVbronchiolitisandpedi-atricasthma.
NFKBIAisthegenewhichencodesIjBa,themajornegativeregulatorofNF-jB.
NF-jBisanimportantnucleartranscriptionfactorthathasacentralroleinregu-latingmultiplehostimmuneandinammatoryresponseelements.
NFKBIAmayendupastheunifyingconceptforbronchiolitisandallergy,asimplicatedbytheresultsofthisstudy,andthisopensupexcitingnewresearchdirectionsforpatientidentication,stratication,andtargetedtherapy.
RSVandParticulateMatter:AnEvenTighterBondThanPreviouslyThoughtEveryday,humansarebombardedwithnumerousinhaledagentsfromtheindoorandoutdoorenvironment.
Particu-latematterrelatedtoairpollutionisbutoneexampleofsuchanagent.
PreviousworkthatstudiedinteractionsbetweenRSVandparticulatematterfocusedoneffectsonthecellularinammatoryresponseinvitro[76–79]andonalargerscaleondeningtemporalrelationshipsbetweenairpollutionexposureandvirusinfectionsandtheireffectsonrespiratoryhealth[80,81].
Fromthesestudies,itisapparentthatairpollutionshouldbeconsideredasapotentialcon-founderoftheeffectivenessofRSVinterventions.
ArecentstudyhasprovidedanevenmoredramaticrelationshipbetweenRSVandparticulatematter[82].
Inthiswork,''designerparticles''ofspecicchemicalcom-positionwereconjugatedtohumanRSV,andcellcultureswereinoculatedwiththesecomplexes,withstatisticallysignicantresponsesobservedwithrespecttocytokineproductionbycells.
Interestingly,thesamestudyshowedthatRSVconjugatedtotheseparticlescouldremaininfectiousatambientroomtemperatureforatleast2monthsafterpreparation,withouttheneedforcellcul-ture,orfreezingstockstolowtemperature(-80°Corbelow).
TheseobservationsopenupanentirelynewpotentialmechanismforRSVsurvivaloutsidelivingorganisms,andsuggestamodeofenvironmentaltrans-missionofRSV,separatefromclosecontactofindividualstovirus-containingdropletsorfomites.
ObtainingimprovedunderstandingoftheimplicationsofRSVconjugatedtoparticulatematterrequiresfurtherresearch.
ProgressTowardanRSVVaccine:CloserthanEverSincethediscoveryofRSVin1955–1957[83–85],vaccinedevelopmenthasbeenamajorpriority,yetthereareanumberoflogisticalconsiderationsrelatedtoRSVinfec-tionsaffectingprogressinthisarea,asCrowe[86]sum-marizedoveradecadeago(Table1).
Thetragedythatoccurredinthe1960swithacandidateRSVvaccinethatwaspreparedbyformalininactivationofwholevirusthatunderwenteldtrialsiswelldocumented[87].
Briey,vaccinerecipientsparadoxicallydevelopedsevererclinicaldiseaseonRSVexposure,andtherewereseveraldeathsinthevaccinatedgroup.
Themechanismsresponsibleforvaccine-enhancedillnesswereultimatelydetermineddecadeslaterbyuseofanimalmodels.
ThroughapplicationoftheTh1–Th2paradigmofCD4lympho-cytes,investigatorsdeterminedthatexposureofexperi-mentalanimalstoformalin-inactivatedRSVvaccineresultedinanexaggeratedhostTh2responseaftersub-sequentexposuretoinfectiousvirus.
Thishasstimulatedconsiderableresearchactivitywiththeaimtodesignvac-cinesthatshifttheTh1–Th2balancetowardTh1[88,89].
However,thismaybeanoverlysimplisticapproach,asarecentstudyinmicehasreportedthatshiftingthehostimmuneresponsetowardTh1actuallydecreasesthepro-ductionofRSV-neutralizingantibodies,whichwouldhavetheeffectoflimitingtheeffectivenessofavaccine[90].
InadditiontoTh1andTh2lymphocytes,thesubpopulationofTh17lymphocyteshasbeenimplicatedinRSVinfection,furtheraddingtothecomplexityofthehostimmuneresponse[91].
CurrPediatrRep(2013)1:149–157153123Inrecentyears,anumberofinnovativeandcleverapproachesforvaccineengineeringhavebeendeveloped,includingtheproductionofvirus-likeparticles[92],chi-mericlivevirusvectors[93],gene-basedvectors[94],andtheuseofnoveladjuvants[95].
Atpresent,weareawareofavarietyofcandidateRSVvaccines(ontheorderoftento20differentproductsatvariousstagesofdevelopment)poisedtoenterhumanclinicaltrials,whichwillbeupdatedattheRSVVaccineConference(Porto,Portugal,October2013:http://www.
meetingsmanagement.
com/rsvvw_2013).
Itisdifculttopredictwhich,ifany,oftheseproductscouldendupasaneffectivevaccineforwidespreaduseinhumans.
Currently,thegoalisfortheretobeasimulta-neousconcertedeffortincontinuedscienticvaccinologyandinthecarefuldesignofhumanclinicaltrialprotocols,includingthepossibilityofimmunizingpregnantwomenasaformofprenatalvaccination,inanefforttodenitivelysolvethelong-standingproblemofdeterminingandimplementingasafe,effectiveRSVvaccine.
Achievingthisoutcomewouldrepresentagloriousaccomplishmentforhumankindandhaveenormousandlastingimpactaroundtheglobe.
ConclusionsWearelivinginaneraofacceleratedactivityandprogressinourunderstandingofviralandhostfactorsinRSVinfection,andarepoisedtotranslatediscoveriesmadeinthisrealmintotheclinictoachieveimprovedhealthout-comesforacuteRSVinfectionsandtheirsequelae.
ResearchhasevolvedfromcharacterizingRSV–hostinteractionstotheidenticationofnewtargetsforvaccineandpharmaceuticaldevelopment.
Advancesinsystemsbiology,informatics,cellandstructuralbiology,andbio-logicalengineeringhaveprovidednewandpowerfulapproachestostudyRSVinfections,andtheinuenceofotherenvironmentalfactorssuchasallergensandparticulatematter.
Wearecloserthanevertoachievingthedreamofasafe,effectiveRSVvaccine,andthefuturepromisestobeexcitingandrewarding.
DisclosurePeterMastrangeloandRichardG.
Hegeledeclarethattheyhavenoconictsofinterest.
ReferencesPapersofparticularinterest,publishedrecently,havebeenhighlightedas:Ofimportance1.
CollinsPL,MeleroJA.
Progressinunderstandingandcontrol-lingrespiratorysyncytialvirus:stillcrazyafteralltheseyears.
VirusRes.
2011;162:80–99.
ThisisarecentcomprehensivereviewofRSV.
2.
WelliverRC.
Pharmacotherapyofrespiratorysyncytialvirusinfection.
CurrOpinPharmacol.
2010;10:289–93.
3.
ShaikhFY,UtleyTJ,CravenRE,etal.
Respiratorysyncytialvirusassemblesintostructuredlamentousvirionparticlesindependentlyofhostcytoskeletonandrelatedproteins.
PLoSONE.
2012;7:e40826.
4.
MufsonMA,OrvellC,RafnarB,NorrbyE.
Twodistincttypesofhumanrespiratorysyncytialvirus.
JGenVirol.
1985;66:2111–24.
5.
McConnachieKM,HallCB,WalshEE,RoghmannKJ.
Variationinseverityofrespiratorysyncytialvirusinfectionswithsubtype.
JPediatr.
1990;117:52–62.
6.
BrandenburgAH,vanBeekR,MollHA,OsterhausAD,ClaasEC.
GproteinvariationinrespiratorysyncytialvirusgroupAdoesnotcorrelatewithclinicalseverity.
JClinMicrobiol.
2000;38:3849–52.
7.
MooreML,ChiMS,LuongoC,etal.
AchimericA2strainofrespiratorysyncytialvirus(RSV)withthefusionproteinofRSVstrainline19exhibitsenhancedviralload,mucus,andairwaydysfunction.
JVirol.
2009;83:4185–94.
8.
BukreyevA,YangL,FrickeJ,etal.
ThesecretedformofrespiratorysyncytialvirusGglycoproteinhelpsthevirusevadeantibody-mediatedrestrictionofreplicationbyactingasanantigendecoyandthrougheffectsonFcreceptor-bearingleu-kocytes.
JVirol.
2008;82:12191–204.
9.
TechaarpornkulS,CollinsPL,PeeplesME.
Respiratorysyncytialviruswiththefusionproteinasitsonlyviralglycoproteinislessdependentoncellularglycosaminoglycansforattachmentthancompletevirus.
Virology.
2002;294:296–304.
10.
BatonickM,OomensAGP,WertzGW.
Humanrespiratorysyn-cytialvirusglycoproteinsarenotrequiredforapicaltargetingandreleasefrompolarizedepithelialcells.
JVirol.
2008;82:8664–72.
11.
McLellanJS,ChenM,KimA,etal.
Structuralbasisofrespi-ratorysyncytialvirusneutralizationwithmotavizumab.
NatStructMolBiol.
2010;17:248–250.
ThisisanoutstandingexampleofadvancesinstructuralbiologyprovinginformationontheinteractionofRSVwithantibodies.
12.
McLellanJS,YangY,GrahamBS,KwongPD.
Structureoftherespiratorysyncytialvirusfusionglycoproteininthepost-fusionconformationrevealspreservationofneutralizingepitopes.
JVirol.
2011;85:7788–96.
13.
JohnsonPR,CollinsPL.
ThefusionglycoproteinsofhumanrespiratorysyncytialvirusofsubgroupsAandB:sequenceconservationprovidesastructuralbasisforantigenicrelatedness.
JGenVirol.
1988;69:2623–8.
Table1ObstaclestoRSVvaccinedevelopmentAnimalmodelsarenotfullypermissiveforinfectionRequirementforbivalentvaccine(twoantigenicsubgroups)YoungageofprimaryvaccinetargetpopulationSpecialsafetyconcernwhenimmunizingneonatesImmunologicimmaturityofneonataltargetpopulationMaternalantibodiesinhibitimmuneresponsesinthetargetpopulationDiseaseoccursattheportalofentry(therespiratorymucosa)Mucosalimmunitytorespiratoryvirusesisshort-livedandincompleteFI-RSVvaccinecausedenhanceddiseaseReproducedwithpermissionfromCrowe[86]154CurrPediatrRep(2013)1:149–15712314.
AlwanW,OpenshawP.
DistinctpatternsofT-andB-cellimmunitytorespiratorysyncytialvirusinducedbyindividualviralproteins.
Vaccine.
1993;11:431–7.
15.
TriantalouK,KarS,VakakisE,KotechaS,TriantalouM.
HumanrespiratorysyncytialvirusviroporinSH:aviralrecog-nitionpathwayusedbythehosttosignalinammasomeactiva-tion.
Thorax.
2013;68:66–75.
16.
TechaarpornkulS,BarrettoN,PeeplesME.
Functionalanalysisofrecombinantrespiratorysyncytialvirusdeletionmutantslackingthesmallhydrophobicand/orattachmentglycoproteingene.
JVirol.
2001;75:6825–34.
17.
ZhangL,PeeplesME,BoucherGC,etal.
Respiratorysyncytialvirusinfectionofhumanairwayepithelialcellsispolarized,specictociliatedcells,andwithoutobviouscytopathology.
JVirol.
2002;76:5654–66.
18.
PanuskaJR,CirinoNM,MidullaF,DespotJE,McFaddenERJr,HuangTY.
Productiveinfectionofisolatedhumanalveolarmacrophagesbyrespiratorysyncytialvirus.
JClinInvest.
1990;86:113–9.
19.
deGraaffPM,deJongEC,vanCapelTM,etal.
Respiratorysyncytialvirusinfectionofmonocyte-deriveddendriticcellsdecreasestheircapacitytoactivateCD4Tcells.
JImmunol.
2005;175:5904–11.
20.
ArensMQ,SwierkoszEM,SchmidtRR,ArmstrongT,RivetnaKA.
Enhancedisolationofrespiratorysyncytialvirusincellculture.
JClinMicrobiol.
1986;23:800–2.
21.
LiXQ,FuZF,AlvarezR,HendersonC,TrippRA.
Respiratorysyncytialvirus(RSV)infectsneuronalcellsandprocessesthatinnervatethelungbyaprocessinvolvingRSVGprotein.
JVirol.
2006;80:537–40.
22.
OsiowyC,AndersonR.
Neutralizationofrespiratorysyncytialvirusaftercellattachment.
JVirol.
1995;69:1271–4.
23.
JohnsonJE,GonzalesRA,OlsonSJ,WrightPF,GrahamBS.
Thehistopathologyoffataluntreatedhumanrespiratorysyncytialvirusinfection.
ModPathol.
2007;20:108–19.
24.
BramleyAM,VitalisTZ,WiggsBR,HegeleRG.
Effectsofrespiratorysyncytialviruspersistenceonairwayresponsivenessandinammationinguinea-pigs.
EurRespirJ.
1999;14:1061–7.
25.
Proenca-ModenaJL,PereiraValeraFC,JacobMG,etal.
Highratesofdetectionofrespiratoryvirusesintonsillartissuesfromchildrenwithchronicadenotonsillardisease.
PLoSOne.
2012;7:e42136.
ThisprovidesnewevidenceofexpandedRSVtissuetropism.
26.
RosenbergHF,DomachowskeJB.
Inammatoryresponsestorespiratorysyncytialvirus(RSV)infectionandthedevelopmentofimmunomodulatorypharmacotherapeutics.
CurrMedChem.
2012;19:1424–31.
27.
GroskreutzDJ,MonickMM,YarovinskyTO,etal.
Respiratorysyncytialvirusdecreasesp53proteintoprolongsurvivalofair-wayepithelialcells.
JImmunol.
2007;179:2741–7.
28.
PringleCR,ShirodariaPV,CashP,ChiswellDJ,MalloyP.
Ini-tiationandmaintenanceofpersistentinfectionbyrespiratorysyncytialvirus.
JVirol.
1978;28:199–211.
29.
WongDT,RosenbandM,HoveyK,OgraPL.
Respiratorysyn-cytialvirusinfectioninimmunosuppressedanimals:implicationsinhumaninfection.
JMedVirol.
1985;17:359–70.
30.
DakhamaA,VitalisTZ,HegeleRG.
Persistenceofrespiratorysyncytialvirus(RSV)infectionanddevelopmentofRSV-specicIgG1responseinaguinea-pigmodelofacutebronchiolitis.
EurRespirJ.
1997;10:20–6.
31.
SchwarzeJ,O'DonnellDR,RohwedderA,OpenshawPJ.
LatencyandpersistenceofrespiratorysyncytialvirusdespiteTcellimmunity.
AmJRespirCritCareMed.
2004;169:801–5.
32.
MejasA,Chavez-BuenoS,GomezAM,etal.
Respiratorysyn-cytialviruspersistence:evidenceinthemousemodel.
PediatrInfectDisJ.
2008;27(10Suppl):S60–2.
33.
FalseyAR,FormicaMA,HennesseyPA,CriddleMM,SullenderWM,WalshEE.
Detectionofrespiratorysyncytialvirusinadultswithchronicobstructivepulmonarydisease.
AmJRespirCritCareMed.
2006;173:639–43.
34.
HabibiMS,OpenshawPJ.
Benetandharmfromimmunitytorespiratorysyncytialvirus:implicationsfortreatment.
CurrOpinInfectDis.
2012;25:687–94.
35.
WangH,PetersN,SchwarzeJ.
Plasmacytoiddendriticcellslimitviralreplication,pulmonaryinammation,andairwayhyperre-sponsivenessinrespiratorysyncytialvirusinfection.
JImmunol.
2006;177:6263–70.
36.
MellaC,Suarez-ArrabalMC,LopezS,etal.
Innateimmunedysfunctionisassociatedwithenhanceddiseaseseverityininfantswithsevererespiratorysyncytialvirusbronchiolitis.
JInfectDis.
2013;207:564–73.
37.
GargR,ShrivastavaP,vanDrunenLittel-vandenHurkS.
Theroleofdendriticcellsininnateandadaptiveimmunitytorespi-ratorysyncytialvirus,andimplicationsforvaccinedevelopment.
ExpertRevVaccines.
2012;11:1441–57.
38.
WuW,MacdonaldA,HiscoxJA,BarrJN.
DifferentNF-jBactivationcharacteristicsofhumanrespiratorysyncytialvirussubgroupsAandB.
MicrobPathog.
2012;52:184–91.
39.
PuthothuB,ForsterJ,HeinzmannA,KruegerM.
TLR-4andCD14polymorphismsinrespiratorysyncytialvirusassociateddisease.
DisMarkers.
2006;22:303–8.
40.
SekiE,YoshizumiM,TanakaR,etal.
Cytokineproles,sig-nallingpathwaysandeffectsofuticasonepropionateinrespi-ratorysyncytialvirus-infectedhumanfoetallungbroblasts.
CellBiolInt.
2013.
doi:10.
1002/cbin.
10044.
41.
MarrN,TurveySE.
RoleofhumanTLR4inrespiratorysyncytialvirus-inducedNF-jBactivation,viralentryandreplication.
InnateImmun.
2012;18:856–65.
42.
MarchantD,SingheraGK,UtokaparchS,etal.
Toll-likereceptor4-mediatedactivationofp38mitogen-activatedproteinkinaseisadeterminantofrespiratoryvirusentryandtropism.
JVirol.
2010;84:11359–73.
43.
CavallaD.
Predictivemethodsindrugrepurposing:goldmineorjustabiggerhaystackDrugDiscovToday.
2012.
doi:10.
1016/j.
drudis.
2012.
12.
009.
44.
TayyariF,MarchantD,MoraesTJ,DuanW,MastrangeloP,HegeleRG.
Identicationofnucleolinasacellularreceptorforhumanrespiratorysyncytialvirus.
NatMed.
2011;17:1132–1135.
ThisdescribesthediscoveryandcharacterizationoftheRSVfusionreceptor.
45.
HovanessianAG,SoundaramourtyC,ElKhouryD,NondierI,SvabJ,KrustB.
Surfaceexpressednucleolinisconstantlyinducedtomediatecalcium-dependentligandinternalization.
PLoSONE.
2010;5:e15787.
46.
DengJS,BallouB,HofmeisterJK.
Internalizationofanti-nu-cleolinantibodyintoviableHEp-2cells.
MolBiolRep.
1996;23:191–5.
47.
ChenX,KubeDM,CooperDJ,DavisPB.
CellsurfacenucleolinservesasareceptorforDNAnanoparticlescomposedofpegy-latedpolylysineandDNA.
MolTher.
2007;16:333–42.
48.
KrustB,ElKhouryD,NondierI,SoundaramourtyC,Hovanes-sianAG.
TargetingsurfacenucleolinwithmultivalentMH-19andrelatedNucantpseudopeptidesresultsindistinctinhibitorymechanismsdependingonthemalignanttumorcelltype.
BMCCancer.
2011;11:333.
49.
SrivastavaM,PollardHB.
Moleculardissectionofnucleolin'sroleingrowthandcellproliferation:newinsights.
FASEBJ.
1999;13:1911–22.
50.
WuW,MundayDC,HowellG,PlattG,BarrJN,HiscoxJA.
Characterizationoftheinteractionbetweenhumanrespiratorysyncytialvirusandthecellcycleincontinuouscellcultureandprimaryhumanairwayepithelialcells.
JVirol.
2011;85:10300–9.
CurrPediatrRep(2013)1:149–15715512351.
SwedanS,AndrewsJ,MajumdarT,MusiyenkoA,BarikS.
Multiplefunctionaldomainsandcomplexesofthetwonon-structuralproteinsofhumanrespiratorysyncytialviruscontributetointerferonsuppressionandcellularlocation.
JVirol.
2011;85:10090–100.
52.
LindquistME,LiandAW,UtleyTJ,SantangeloPJ,CroweJEJr.
RespiratorysyncytialvirusinduceshostRNAstressgranulestofacilitateviralreplication.
JVirol.
2010;84:12274–84.
53.
Gutierrez-OrtegaA,Sanchez-HernandezC,Gomez-GarcaB.
Respiratorysyncytialvirusglycoproteinsuptakeoccursthroughclathrin-mediatedendocytosisinahumanepithelialcellline.
VirolJ.
2008;5:127.
54.
JeffreeCE,BrownG,AitkenJ,Su-YinDY,TanBH,SugrueRJ.
UltrastructuralanalysisoftheinteractionbetweenF-actinandrespiratorysyncytialvirusduringvirusassembly.
Virology.
2007;369:309–23.
55.
ChangTH,SegoviaJ,SabbahA,MgbemenaV,BoseS.
Cho-lesterol-richlipidraftsarerequiredforreleaseofinfectioushumanrespiratorysyncytialvirusparticles.
Virology.
2012;422:205–13.
56.
HegeleRG,SekhonMS,KaanP.
Towardssystemsbiologyofrespiratorysyncytialvirusinfections:seeingtheneedandpre-paringforprimetime.
CurrRespirMedRev.
2008;4:29–34.
57.
RaviLI,LiL,SutejoR,ChenH,WongPS,TanBH,SugrueRJ.
Asystems-basedapproachtoanalysethehostresponseinmurinelungmacrophageschallengedwithrespiratorysyncytialvirus.
BMCGenomics.
2013;14:190.
58.
WuW,TranKC,TengMN,etal.
TheinteractomeofthehumanrespiratorysyncytialvirusNS1proteinhighlightsmultipleeffectsonhostcellbiology.
JVirol.
2012;86:7777–7789.
ThisisacontemporaryexampleofhowsystemsbiologyandinformaticsapproachesareyieldingnovelinsightsintoRSV–hostcellinteractions.
59.
VardiL,RuppinE,SharanR.
Alinearizedconstraint-basedapproachformodelingsignalingnetworks.
JComputBiol.
2012;19:232–40.
60.
WittigHJ,CranfordNJ,GlaserJ.
Therelationshipbetweenbronchiolitisandchildhoodasthma.
JAllergy.
1959;30:19–23.
61.
SteinRT,SherrillD,MorganWJ,etal.
Respiratorysyncytialvirusinearlylifeandriskofwheezeandallergybyage13years.
Lancet.
1999;35:541–5.
62.
SigursN,AljassimF,KjellmanB,etal.
Asthmaandallergypatternsover18yearsaftersevereRSVbronchiolitisintherstyearoflife.
Thorax.
2010;65:1045–52.
63.
SteinRT,MartinezFD.
Respiratorysyncytialvirusandasthma:stillnonalanswer.
Thorax.
2010;65:1033–4.
64.
WelliverRC,WongDT,SunM,MiddletonEJr,VaughanRS,OgraPL.
Thedevelopmentofrespiratorysyncytialvirus-specicIgEandthereleaseofhistamineinnasopharyngealsecretionsafterinfection.
NEnglJMed.
1981;305:841–6.
65.
FrickOL,GermanDF,MillsJ.
Developmentofallergyinchil-dren.
I.
Associationwithvirusinfections.
JAllergyClinImmu-nol.
1979;63:228–41.
66.
LeeKK,HegeleRG,ManfredaJ,etal.
Relationshipofearlychildhoodviralexposurestorespiratorysymptoms,onsetofpos-sibleasthmaandatopyinhighriskchildren:theCanadianAsthmaPrimaryPreventionStudy.
PediatrPulmonol.
2007;42:290–7.
67.
HegeleRG,RobinsonPJ,GonzalezS,HoggJC.
Productionofacutebronchiolitisinguinea-pigsbyhumanrespiratorysyncytialvirus.
EurRespirJ.
1993;6:1324–31.
68.
BucknerCK,SongsiridejV,DickEC,BusseWW.
Invivoandinvitrostudiesontheuseoftheguineapigasamodelforvirus-provokedairwayhyperreactivity.
AmRevRespirDis.
1985;132:305–10.
69.
IshidaK,KellyLJ,ThomsonRJ,BeattieLL,SchellenbergRR.
Repeatedantigenchallengeinducesairwayhyperresponsivenesswithtissueeosinophiliainguineapigs.
JApplPhysiol.
1989;67:1133–9.
70.
HegeleRG,HayashiS,BramleyAM,HoggJC.
Persistenceofrespiratorysyncytialvirusgenomeandproteinafteracutebron-chiolitisinguineapigs.
Chest.
1994;105:1848–54.
71.
SuttonTC,TayyariF,KhanMA,MansonHE,HegeleRG.
Thelper1backgroundprotectsagainstairwayhyperresponsivenessandinammationinguineapigswithpersistentrespiratorysyn-cytialvirusinfection.
PediatrRes.
2007;61(5Pt1):525–9.
72.
DakhamaA,BramleyAM,ChanNG,McKayKO,SchellenbergRR,HegeleRG.
Effectofrespiratorysyncytialvirusonsub-sequentallergicsensitizationtoovalbumininguinea-pigs.
EurRespirJ.
1999;13:976–82.
73.
TayyariF,SuttonTC,MansonHE,HegeleRG.
CpG-oligo-deoxynucleotidesinhibitRSV-enhancedallergicsensitisationinguineapigs.
EurRespirJ.
2005;25:295–302.
74.
ZengR,LiC,LiN,WeiL,CuiYX.
Theroleofcytokinesandchemokinesinsevererespiratorysyncytialvirusinfectionandsubsequentasthma.
Cytokine.
2011;53:1–7.
Thisisacompre-hensivereviewofinammatorymediatorsandgeneticdetermi-nantsinsevereformsofRSVinfectionandtheirsequelae.
75.
AliS,HirschfeldAF,MayerML,etal.
FunctionalgeneticvariationinNFKBIAandsusceptibilitytochildhoodasthma,bronchiolitis,andbronchopulmonarydysplasia.
JImmunol.
2013;190:3949–58.
Thisdescribesthepotentialgenetic''missinglink''betweenRSVbronchiolitisandasthma.
76.
BeckerS,KorenHS,HenkeDC.
Interleukin-8expressioninnormalnasalepitheliumanditsmodulationbyinfectionwithrespiratorysyncytialvirusandcytokinestumornecrosisfactor,interleukin-1,andinterleukin-6.
AmJRespirCellMolBiol.
1993;8:20–7.
77.
SoukupJ,KorenHS,BeckerS.
Ozoneeffectonrespiratorysyncytialvirusinfectivityandcytokineproductionbyhumanalveolarmacrophages.
EnvironRes.
1993;60:178–86.
78.
KaanPM,HegeleRG.
Interactionbetweenrespiratorysyncytialvirusandparticulatematteringuineapigalveolarmacrophages.
AmJRespirCellMolBiol.
2003;28:697–704.
79.
HackettTL,SingheraGK,ShaheenF,etal.
Intrinsicphenotypicdifferencesofasthmaticepitheliumanditsinammatoryresponsestorespiratorysyncytialvirusandairpollution.
AmJRespirCellMolBiol.
2011;45:1090–100.
80.
WongCM,ThachTQ,ChauPY,etal.
Part4.
Interactionbetweenairpollutionandrespiratoryviruses:time-seriesstudyofdailymortalityandhospitaladmissionsinHongKong.
ResRepHealthEffInst.
2010;154:283–362.
81.
VandiniS,CorvagliaL,AlessandroniR,etal.
Respiratorysyn-cytialvirusinfectionininfantsandcorrelationwithmeteoro-logicalfactorsandairpollutants.
ItalJPediatr.
2013;39:1.
82.
Cruz-SanchezTM,HaddrellAE,HackettTL,etal.
Formationofastablemimicofambientparticulatemattercontainingviableinfectiousrespiratorysyncytialvirusanditsdry-depositiondirectlyontocellcultures.
AnalChem.
2013;85:898–906.
ThisdescribesanewmechanismofprolongedRSVsurvivaloutsidecells,withprofoundimplicationsforvirusviabilityandtransmission.
83.
MorrisJA,BlountRE,SavageRE.
Recoveryofcytopathicagentfromchimpanzeeswithcoryza.
ProcSocExpBiolMed.
1956;92:544–50.
84.
ChanockRM,RoizmanB,MyersR.
Recoveryfrominfantswithrespiratoryillnessofavirusrelatedtochimpanzeecoryzaagent.
I.
Isolation,propertiesandcharacterization.
AmJHygiene.
1957;66:281–90.
85.
ChanockRM,FinbergL.
Recoveryfrominfantswithrespiratoryillnessofavirusrelatedtochimpanzeecoryzaagent(CCA).
II.
Epidemiologicalaspectsofinfectionininfantsandyoungchil-dren.
AmJHygiene.
1957;66:291–300.
156CurrPediatrRep(2013)1:149–15712386.
CroweJEJr.
Respiratorysyncytialvirusvaccinedevelopment.
Vaccine.
2001;20(Suppl.
1):S32–7.
87.
KimHW,CancholaJG,BrandtCD,etal.
Respiratorysyncytialvirusdiseaseininfantsdespiteprioradministrationofantigenicinactivatedvaccine.
AmJEpidemiol.
1969;89:422–34.
88.
WarisME,TsouC,ErdmanDD,ZakiSR,AndersonLJ.
Respi-ratorysynctialvirusinfectioninBALB/cmicepreviouslyimmunizedwithformalin-inactivatedvirusinducesenhancedpulmonaryinammatoryresponsewithapredominantTh2-likecytokinepattern.
JVirol.
1996;70:2852–60.
89.
BoukhvalovaMS,PrinceGA,etal.
TheTLR4agonist,mono-phosphoryllipidA,attenuatesthecytokinestormassociatedwithrespiratorysyncytialvirusvaccine-enhanceddisease.
Vaccine.
2006;24:5027–35.
90.
TregoningJS,WangBL,McDonaldJU,etal.
Neonatalanti-bodyresponsesareattenuatedbyinterferon-cproducedbyNKcellsandTcellsduringRSVinfection.
ProcNatAcadSciUSA.
2013;110:5576–81.
ThisarticlecastsdoubtondesignofvaccinestoshiftthehostTh1–Th2balancetowardTh1response.
91.
BystromJ,Al-AdhoubiN,Al-BogamiM,JawadAS,MageedRA.
Th17lymphocytesinrespiratorysyncytialvirusinfection.
Viruses.
2013;5:777–91.
92.
QuanF-S,KimY,LeeS,etal.
Viruslikeparticlevaccineinducesprotectionagainstrespiratorysyncytialvirusinfectioninmice.
JInfectDis.
2011;204:987–95.
93.
MokH,ChengX,XuQ,etal.
EvaluationofameaslesvaccinevirusasavectortodeliverrespiratorysyncytialvirusfusionproteinorEpstein-Barrvirusglycoproteingp350.
OpenVirolJ.
2012;6:12–22.
94.
McLellanJS,CorreiaBE,ChenM,etal.
Designandcharacter-izationofepitope-scaffoldimmunogensthatpresentthemot-avizumabepitopefromrespiratorysyncytialvirus.
JMolBiol.
2011;409:853–66.
95.
LindellDM,MorrisSB,WhiteMP,etal.
AnovelinactivatedintranasalrespiratorysyncytialvirusvaccinepromotesviralclearancewithoutTh2associatedvaccine-enhanceddisease.
PLoSONE.
2011;6:321823.
CurrPediatrRep(2013)1:149–157157123
HegelePublishedonline:15May2013SpringerScience+BusinessMediaNewYork2013AbstractRespiratorysyncytialvirus(RSV)isamajorworldwidepathogenforwhichthereisstillnoeffectivevaccineorantiviraltreatmentavailable,andimmunopro-phylaxiswithRSV-specicantibodies(e.
g.
,palivizumab)isusedinlimitedclinicalsettings.
Inthisreview,wedis-cussvirus–hostinteractionsrelevanttoRSVpathobiologyandhowadvancesincellandsystemsbiologyhaveacceleratedknowledgeinthisarea.
WealsohighlightrecentadvancesinunderstandingtherelationshipbetweenRSVbronchiolitisandsequelaeofrecurrentwheezingandasthma,newndingsintoanintriguinginteractionbetweenRSVandairpollution,andexcitingdevelopmentstowardthegoalofrealizingasafeandeffectiveRSVvaccine.
KeywordsRespiratorysyncytialvirusViralpathogenesisVirus–hostinteractionsViruspersistenceAsthmaAllergySystemsbiologyNucleolinAirpollutionVaccinologyIntroductionRespiratorysyncytialvirus(RSV)isasingle-stranded,negative-polarityPneumovirusthatinfectspeopleofallagesworldwide[1].
RSVisafrequentcauseofupperrespiratorytractinfectionsandotitismedia,andalsocausesseriouslowerrespiratorytractinfectionssuchasbronchi-olitisandpneumonia,particularlyininfantsandyoungchildren,immunocompromisedindividuals,andtheelderly.
RSVinfectionsinearlypostnatallifehavealsobeenassociatedwiththeonsetofallergicsensitizationandrecurrentwheezing/asthma.
Despitealmost60yearshav-ingpassedsincethediscoveryofRSV,thereisstillnolicensedvaccineavailable,andprophylaxiswithimmu-noglobulin(palivizumab)isusedonlyforasubsetof''high-risk''children(e.
g.
,pretermbirths,andinfantswithcongenitalheartdisease,congenitalimmunodeciency,and/orbronchopulmonarydysplasia).
AlthoughanumberofpharmacologicalagentshavebeenusedinRSVtherapy,theirefcacyandeffectivenessarepoor[2].
Here,wewillbrieyreviewsomeviralandhostfactorsimplicatedinthepathobiologyofRSVinfections,focusingonhowthediversityofthesefactorsmightberelatedtothevariabilityseeninclinicaldisease.
OurdiscussionwillalsoincludecomparingandcontrastingcharacteristicsofRSVinfectionsinvitroversusinvivo;currentperspectivesontherelationshipbetweenRSVinfections,pediatricallergy,andasthma;recentworkthatdescribesanovelrelationshipbetweenRSVandparticulatematter(airpollution);andprospectsforthedevelopmentandimplementationofasafe,effectiveRSVvaccine.
ViralandHostFactorsinRSVInfection:TowardanIntegratedPerspectiveRSVStructureHumanRSVisalamentousenvelopedvirusinwhichtheviralnucleicacidandstructuralandinternalproteinsaresurroundedbyaproteinaceousoutercoat[3].
AsanP.
MastrangeloR.
G.
Hegele(&)DepartmentofLaboratoryMedicineandPathobiology,UniversityofToronto,6231-1King'sCollegeCircle,Toronto,ONM5S1A8,Canadae-mail:richard.
hegele@utoronto.
caR.
G.
HegeleDepartmentofPaediatricLaboratoryMedicine,TheHospitalforSickChildren,Toronto,ON,Canada123CurrPediatrRep(2013)1:149–157DOI10.
1007/s40124-013-0019-3envelopedvirus,RSV'sgeneralstructurehassimilaritiestothatofinuenzaandparainuenzaviruses,metapneum-oviruses,andcoronaviruses,andcontrastswiththatofothercommonrespiratorypathogenssuchasrhinovirusesandadenoviruses,whicharenonenveloped.
RSVhasthreeenvelopeproteins:G(glycoprotein),F(fusion),andSH(smallhydrophobic)[1].
HeterogeneityintheGglyco-proteindenesthetwomainsubgroupsofRSV,AandB[4].
SomeliteraturesuggeststhatRSVAstrainsproducemoreclinicallysevereinfectionsthanRSVBstrains[5],butthiscontentionhasprovendifculttoreproduceamongdifferentgroupsofinvestigators[6].
OnestrainofRSVA,so-calledline19[7],appearstobeofparticularinterestregardingmucusproductionandsecretionofIL-13,aTh2cytokineassociatedwithallergic-typehostresponses(seebelow),andtherebycontributinginparttotheheteroge-neityandseverityofclinicalmanifestations.
Inaddition,RSVGhasasecretedformthatbindshostantibodiesandtherebyservesasanimmunological''decoy''thatfacili-tatescontinuedviralreplication[8].
RSVGisalsocon-sideredtobeakeyattachmentproteinforthevirustobindtothecellsurfacetoinitiateinfection;however,withthediscoveryandengineeringofRSVstrainsdecientinG(so-calledRSVDG)thatareinfectious,albeitatwithmuchlowerefciencythanvirusexpressingG,RSVGisneithersufcientnornecessarytocauseinfection[9].
Bycontrast,RSVFiscrucialforfusionoftheviralenvelopetothehostcellsurface,sinceRSVDFcannotentercellsonitsownbutratherrequiresahelpervirustoinitiateinfection[10].
Onfreevirus,RSVFexistsina''prefusion''formthatundergoesanelegantlycharacterizedconforma-tionalchangeduringthefusionprocess[11,12].
IncontrasttoRSVG,RSVFishighlyconservedbetweenthetwomajorsubgroupsofthevirus[13].
RSVFandRSVGmayhaveanoverall''yin-yang''effectonhostadaptiveimmuneresponsesconcerningTh1–Th2balance.
Inbrief,Th1responsesinvolveasubsetofCD4lymphocytesproducingcytokinessuchasinterferon(IFN)-candinterleukin(IL)-2consideredimportantindelayedhypersensitivityandanti-viralimmunity,whereasTh2responsesinvolveproductionofinammatorymediatorssuchasIL-4,IL-5,andIL-13byadifferentsubsetofCD4lymphocytesandareimplicatedinallergy.
RSVFappearstostimulateTh1cells,whereasRSVGappearstostimulateTh2responses,andclinicalmanifestationsarebelievedtoreectwhethertheoverallresponseisshiftedtowardTh1orTh2,orremainsinbalancewithregardtocytokineproduction[14].
TheRSVSHprotein,byanalogytosimilarproteinsexpressedinotherviruses,mayhavearoleinenhancingthepermeabilityofthecellsurfacetomoleculesandions[15].
TheimportanceofthisputativefunctionofRSVSHisunclear,sincegeneticallyengineeredstrainsofRSVinwhichSHisdeletedappeartobeessentiallyasefcientaswild-typevirusincausingcellularinfections[16].
ThereisincreasinginterestinSHasapotentialviralantigentargetinvaccinedevelopment(seelater).
OtherRSVcompo-nents,includingnucleocapsid(N),polymerase(L),non-structuralcomponents(NS1,NS2),phosphoprotein(P),andmatrix(M,M2-1,M2-2)havebeenimplicatedinstimulatinghostantiviralresponses(e.
g.
,IFN-aandIFN-b)througheffectsinhostcellsignaltransductionpathwaysandimportantnucleartranscriptionfactors(e.
g.
,NF-jB,AP-1).
FurtherdetailsoftheroleofnonenvelopedproteinsinRSVreplicationandhostresponsesareprovidedinanexcellentreviewbyCollinsandMelero[1].
RSVInfectionsInVitroVersusInVivoOneofthebiggestenigmassurroundingRSVpathobiologyisthestrongtropismthevirusshowsforinfectingthehumanrespiratorytractinvivo;invitro,RSVestablishesproductiveinfectionofvirtuallyallmammaliancelltypesstudied,includingawidevarietyofprimaryandcontinuouscellslinesofepithelialcells[17],macrophages[18],otherleukocytes[19],broblasts[20],andevenneurons[21].
Infact,ndingacelltypethatisresistanttoRSVinfectioninvitroischallenging.
Aswillbediscussedin''NucleolinasaFusionReceptorforRSV:FromDiscoverytoDrugTargeting''inthecontextofthediscoveryofacellularfusionreceptorforRSV,Sf9cellsfromthemothSpo-dopterafrugiperdaaredifculttoinfectwithRSV[22].
InadenitiveautopsystudyofthelungpathologyofuntreatedacuteRSVinfections,Johnsonetal.
[23]repor-tedthatRSVantigenswereobservedwithinairwayandalveolarepithelialcells,aswellaspulmonarymacro-phages.
Similarobservationshavebeenmadeinexperi-mentalanimals.
InaguineapigmodelofexperimentalRSVinfection,ourgrouphasdocumentedtheexpressionofviralantigensviaimmunohistochemistrywithinlungepithelialcells,alveolarmacrophages,interstitialcells(presumablydendriticcells),andcellswithinbronchus-associatedlymphoidtissue,foratleast100daysafterRSVinoculation[24].
ArecentstudyhasreportedthatRSVcanbedetectedinapproximately15%ofhumantonsilsofchildrenwithchronicadenotonsillardisease[25].
GiventheabundantliteratureconcerningtheeffectsofRSVinfectiononthesecretionofvariousinammatorymedia-tors,responsesofcellsignalingpathways,andotherhostresponsesobservedinvitro,onemustexercisecautioninextrapolatingthesignicanceofobservationsmadeinvitrototheinvivostate,andwhethertheyrepresentopportu-nitiesforpotentialclinicalinterventions.
OneonlyneedstoreectonthehistoricallydisappointingresultsseenwithattemptstotreatRSVinfectionsofexperimentalanimalsorpatientswithantiviraloranti-inammatoryagentstoappreciatethispoint[2,26].
150CurrPediatrRep(2013)1:149–157123Incontrasttoothervirusessuchasinuenzavirusandadenovirus,whereextensivecytotoxicityandcelldeathisthenormwithinfectioninvitro,RSVinfectiondoesnotelicitmuchinthewayofcelldamage[17].
Thereisevi-dencethatRSVcanactuallydecreasetherateofapoptosisinvitro[27].
Takenfurther,RSVhasclearlybeenshowntocausepersistent,noncytolyticinfectionofepithelialcellsinvitro[28].
RSVpersistence,characterizedbycontinu-ous,low-levelreplicationofinfectiousvirus,hasbeendescribedinvivo,rstinamodelinwhichcottonratswereimmunosuppressedwithcyclophosphamide[29],andsub-sequentlyinimmunocompetentguineapigs[30]andinmice[31,32].
ThereisalsoevidenceconsistentwithRSVpersistenceinhumans,bestcharacterizedinthesettingofadultpatientswithchronicobstructivepulmonarydisease[33].
ApossibleroleforRSVpersistenceinthepathobi-ologyofpostbronchiolitiswheezing/asthmaisdescribedin''RSVandPediatricAsthma:HaveWeFoundtheMissingLink''RSV–HostInteractions:MovingEverMoreProximallyFormanyyears,investigationofRSV–hostinteractionsfocusedoncharacterizingtheinammatorymediatorresponsesofinfectedcells,animals,andhumansubjectswiththehopethatanunderstandingofthevirus-specichostresponsestoRSVinfectionscouldinformpossibleinterventionsdesignedtomodulaterelevantinammatorymediatorsandtherebyleadtoimprovedclinicaloutcomes[34].
Theapparentheterogeneityinvirus-specicimmu-nityobservedinhostadaptiveimmuneresponsestoRSV(bothcell-mediatedandhumoral),dependingonthepop-ulationstudied,complicatestheabilitytomakeclinicallyrelevantadvances.
ExtensiveinquirieshavefocusedoninnateimmunemechanismsintheearlyhostresponsetoRSV[35,36].
Onerationaleforthisapproachisthatbyunderstandingtheearly,''proximal''eventsthatoccurinRSVinfections,itmaybeeasiertodevelopeffectiveclinicalinterventionsthatcouldbeusedeffectivelyamongdiversehostswhohavedifferentadaptiveimmuneresponsestothevirus[15,37].
Thisrationalehasbeenrecentlyquestioned,sinceapparentdifferencesininnateimmuneresponseshavebeenobservedbetweenRSVAandRSVBinfections[38].
Nevertheless,researchintotheroleofinnateimmunemechanismsinRSVpathobiologyledtoanumberofpro-foundinsights,oneofwhichwastheabilityofRSVtoincreasethelevelofpattern-recognitionreceptors(e.
g.
,Toll-likereceptors)expressedonthecellsurface[39].
Theseobservationswereinuentialtotheeldforanumberofreasons:forexample,thenatureofcellsurfacemoleculesinvolvedininnateimmunitythatcanbeaffectedbyRSVwaspoorlyunderstood,andpattern-recognitionreceptors(e.
g.
,Toll-likereceptor4andothers)stimulatecellsig-nalingpathwaysthatareamenabletopharmacologicalmanipulation[40],eveniftheyarenotactualreceptorsforthevirus[41].
OnecellsignalingmoleculerelatedtoToll-likereceptorinteractionwithRSVisp38mitogen-activatedproteinkinase,asrevealedbyascreenofculturedcellswithapanelofpharmacologicalkinaseinhibitors.
Inhibitionofp38mitogen-activatedproteinkinasewithSB203580(acompoundoriginallydevelopedfortreatmentofarthritisandcancer)wasassociatedwithmarkedinhibitionofRSVreplicationinculturedcells,withoutinducingsignicantdrug-relatedcytotoxicity[42].
Thistypeofexperimentisanexampleofso-calleddrugrepurposing,inwhichacom-pounddevelopedforanunrelatedclinicalindicationcanbegivenonthebasisofpathobiologydenedinanotherclinicalsetting[43].
Itremainstobeseenwhethersuchanapproachtodrugrepurposingwillbeusefulintheman-agementofRSVinfectionsinvivoinanimalmodels,andultimatelyinhumans.
NucleolinasaFusionReceptorforRSV:FromDiscoverytoDrugTargetingOurgroupidentiedcellsurfacenucleolinasamoleculethatfulllsthecriteriaofafunctionalreceptorforRSV[44].
Oneofthekeychallengestoovercomeinthisdis-coverywasndingacelltyperesistanttoRSVinfectioninvitrowhichcouldbecomepermissiveafterforcedexpressionofthehumannucleolingeneandectopicexpressionofhumannucleolinproteinonthecellsurface.
ThiswasachievedbyuseofinsectSf9cells[22].
Vali-dationofnucleolinasacellularreceptorinvivowasaccomplishedinamousemodelbysmallinterferingRNAknockdownofnucleolinpriortoRSVchallenge,whichwasassociatedwithasignicantdecreaseinRSVtitersasmeasuredbyquantitativeplaqueassaysoflunghomoge-nates.
WeshowedthatRSVF,notRSVG,interactswithnucleolin,andwecoinedtheterm''fusionreceptor''todenotethisfact.
NucleolinisanespeciallychallengingpotentialdrugtargetforRSV.
Nucleolinhasaveryshorthalf-lifewhenexpressedonthecellsurface,andconstantlyundergoesrecyclingbetweenthecytoplasmicandnuclearcompart-mentsofthecell[45].
Thus,approachesthatinterferewithcellsurfacenucleolinsuchasuseofnucleolin-specicblockingantibodies,resultintheantibody–nucleolincomplexbeinginternalizedintothecellandnew(''fresh'')unboundnucleolinmovingtothesurface[46].
Thisphe-nomenoncouldexplainwhyinourantibodyneutralizationexperimentsattemptingtovalidatenucleolinasanRSVreceptorwewereabletoachieveonlyontheorderapproximately70–80%decreasedinfectionofcellsthathadbeenpreincubatedwithnucleolin-specicantibodyCurrPediatrRep(2013)1:149–157151123priortoRSVchallenge.
Secondly,nucleolinisanotori-ously''sticky''moleculethatcanbindallsortsofmole-cules,includingproteinsexpressedbyotherviruses(e.
g.
,parainuenzavirustype3,HIV),certaindrugs(e.
g.
,mid-kine,lactoferrin,pseudopeptideHB-19),andevenDNAmoieties(reviewedin[47]).
HowtotranslatethissituationintoascenariothatisRSV-specicneedstobeaddressed.
Moreover,nucleolindoesnotexistinisolationonthecellsurface;rather,itispartofamultiproteincomplexofmolecularmassofapproximately500kDa[48].
Toaddfurthercontext,intranuclearnucleolinisessentialtocellsurvivalandreplication:inhibitingnucleolininanunde-siredcellularcompartmentcouldelicitseriousoff-targeteffects[49].
Overall,theidenticationofnucleolinasafusionreceptorforRSVposesanumberofprofoundchallengestobeovercomebeforeitmayserveasabonadedrugtarget.
ARenaissanceofRSVCellBiology,andtheGrowingRoleofSystemsBiologyOneareaofincreasingresearchactivityistheintracellularcompartmentalizationofRSVconstituentsduringthevirallifecycle.
CellbiologistsinvestigatingRSVreplicationkineticsdemonstratedthatthevirusdoesnotnecessarilyfollowastereotypedreplicativecycle,butrathercanbepreferentiallydirectedintointracellularcompartmentsthatfavororinhibitreplication[50,51].
Forexample,cyto-plasmicstressgranules[52],caveolae[53],hostcytoskel-eton[54],andlipidrafts[55]havebeenreportedtointeractwithRSV,andthesearepotentialnoveltargetsforphar-macologicalintervention.
Increasingly,theuseofhigh-throughput''systemsbiology''approachesdesignedtointerrogateRSV-infectedcellsinanunbiasedmannerhasledtoidenticationofnovelcandidatemoleculesthatcouldpotentiallybetargetedpharmacologically[56,57].
SystemsbiologypermitstheapplicationofinformaticsrelatedtopathwaymodelingrelevanttoRSVinfection.
Bydeningso-calledinteractomes[58]inwhichthevirusaffectscellfunction,investigatorsaimtoachieveacomprehensiveunderstandingofRSV–hostinteractionstoinformthedevelopmentofnovelstrategiesintreatmentandprophy-laxis.
Oneofthecurrentlimitationsofinformaticsapproachesisthatthebiologicalpathwaysbeingmodelingarethemselvesincompletelycharacterized,someaningfulinterpretationswillrequireongoingiterativeactivity[59].
RSVandPediatricAsthma:HaveWeFoundtheMissingLinkWithinafewyearsofthediscoveryofRSV,inaseminalarticle,Wittigetal.
[60]reportedthatasizeablepercentage(32%)ofinfantsandyoungchildrenhospitalizedforRSVbronchiolitisdevelopedsequelaeofrecurrentwheezingandasthmasymptoms.
Thesendingshavebeenreplicatedonanumberofoccasionsoverthelastfewdecades,althoughthereissomeongoingcontroversyoverwhetherpatientscan''outgrow''theirasthmabythetimetheyreachado-lescence,orwhethertheirconditioncanpersistintoadulthood[61–63].
OnepossibilityisthatRSVbronchi-olitisisa''marker''ofchildrenwhoaregeneticallypre-disposedtodevelopasthma,asevidencedbyaclassicstudyinwhichRSV-specicIgEwasdocumentedinchildrenwhohadrecurrentwheezingasasequelaofRSVinfection[64].
Further,inahighlyinuentialarticle,Fricketal.
[65]reportedatemporalrelationshipbetweenviralinfection(particularlyRSVandparainuenzavirus)andtriggeringallergicsensitizationin14childrenborninto''high-risk''families(i.
e.
,whererst-degreerelativeshadaprovenhistoryofallergy).
Thislatterassociationhasalsobeenconrmedinlargerstudies,includingtheCanadianAsthmaPrimaryPreventionStudyofapproximately400''high-risk''childrenandtheirfamilies[66].
Toexplorepotentialmechanismsofpostbronchiolitiswheezing/asthmaandallergy,ourgroupdevelopedagui-neapigmodelofexperimentalRSVlunginfection[67].
Guineapigswerechosenbecausetheywerealreadyamodelofhumanparainuenzavirustype3infection[68],andcouldreadilybesensitizedtoallergen(aerosolizedovalbumin)[69].
AfterinitialworkestablishingthatRSVproteinandgenomecouldpersistintheguineapiglungforupleast60daysafterRSVinoculation[70],subsequentstudiesshowedthatoutbredCamHartleyguineapigscouldharborreplicatingvirusforatleast60–100daysafterinoculation,inassociationwithnonspecicairwayhyper-responsiveness(AHR)andchronicairwayinammation(physiologicalandpathologicalhallmarksofasthma)[24,30].
However,insubsequentwork,wenotedthatviralpersistenceonitsownisnotsufcienttoproduceasthma-likechangesinthelungs,asinbred,''allergy-resistant''strain2guineapigscanalsodeveloppersistentRSVlunginfectionsbuttheseanimalsdonotgetAHRorchronicairwayinammationinassociationwithviruspersistence[71].
ConcerningthepossibilitythatRSVinfectionisa''marker''ofgeneticpredispositiontoallergy,weobservedthatRSVinfectionofCamHartleyguineapigsenhancedallergicsensitizationtoaerosolizedovalbumin,consistentwiththishypothesis[72].
Interestingly,theextentofchronicAHR/airwayinammationandvirus-enhancedovalbuminsensitizationcouldbeamelioratedbyimmu-noprophylaxisofanimalswithCpG-oligodinucleotides(CpG-ODN),apotentTh1stimulus[73].
Takentogether,theseresultssuggestthatthecombinationofRSVandaeroallergenproducesadditiveeffectsonAHRandairway152CurrPediatrRep(2013)1:149–157123inammationinan''allergy-susceptible''hostandthatCpG-ODNimmunoprophylaxisappearstotargettheRSV-associated,nottheallergen-associated,component.
Alargenumberofhumanstudieshavecharacterizeddifferencesinthetypesofcytokinesandotherchemicalmediatorsofinammation—andtheirpotentialgeneticdeterminants—indifferentsubsetsofRSV-infectedpatients(reviewedinZengetal.
[74]).
ThislargebodyofworkprovidesvaluableinsightsintopossibledifferencesinhostresponsestoRSVinfectionandmayberelevanttoexplainingwhysomeinfectedindividualsdevelopmild,upperrespiratorytractinfectionswhereasotherindividualsdevelopbronchiolitisorpneumonia,orgoontodeveloprecurrentwheezing/asthma.
Unfortunately,therehasbeenarecurrentdifcultyinvalidatingthelargenumberofcan-didatemolecules/genesbyreplicationinothercohorts,andtheoverallpictureisfarfromclear[1].
Theuseofasys-temsbiologyapproach(see''ARenaissanceofRSVCellBiology,andtheGrowingRoleofSystemsBiology'')promisestoaccelerateknowledgeofidenticationofcan-didatemoleculesandgenesforvalidation.
Onerecentstudy[75]hasreportedanexcitingdevelopmentthatmaypro-videacluetothepossiblebiological''missinglink''betweensevereRSVinfectionsandthesubsequentdevel-opmentofpediatricasthma.
Inaseriesofexperimentsthatbeganwithunbiasedinterrogationofcellstoidentifycan-didategenes,followedbyvalidationinexperimentalani-malsandanalysisofhumancohorts,investigatorshavecharacterizedgeneticpolymorphismsinthepromoterregiontoNFKBIAashavingbothfunctionalandclinicalsignicanceinthecontextofRSVbronchiolitisandpedi-atricasthma.
NFKBIAisthegenewhichencodesIjBa,themajornegativeregulatorofNF-jB.
NF-jBisanimportantnucleartranscriptionfactorthathasacentralroleinregu-latingmultiplehostimmuneandinammatoryresponseelements.
NFKBIAmayendupastheunifyingconceptforbronchiolitisandallergy,asimplicatedbytheresultsofthisstudy,andthisopensupexcitingnewresearchdirectionsforpatientidentication,stratication,andtargetedtherapy.
RSVandParticulateMatter:AnEvenTighterBondThanPreviouslyThoughtEveryday,humansarebombardedwithnumerousinhaledagentsfromtheindoorandoutdoorenvironment.
Particu-latematterrelatedtoairpollutionisbutoneexampleofsuchanagent.
PreviousworkthatstudiedinteractionsbetweenRSVandparticulatematterfocusedoneffectsonthecellularinammatoryresponseinvitro[76–79]andonalargerscaleondeningtemporalrelationshipsbetweenairpollutionexposureandvirusinfectionsandtheireffectsonrespiratoryhealth[80,81].
Fromthesestudies,itisapparentthatairpollutionshouldbeconsideredasapotentialcon-founderoftheeffectivenessofRSVinterventions.
ArecentstudyhasprovidedanevenmoredramaticrelationshipbetweenRSVandparticulatematter[82].
Inthiswork,''designerparticles''ofspecicchemicalcom-positionwereconjugatedtohumanRSV,andcellcultureswereinoculatedwiththesecomplexes,withstatisticallysignicantresponsesobservedwithrespecttocytokineproductionbycells.
Interestingly,thesamestudyshowedthatRSVconjugatedtotheseparticlescouldremaininfectiousatambientroomtemperatureforatleast2monthsafterpreparation,withouttheneedforcellcul-ture,orfreezingstockstolowtemperature(-80°Corbelow).
TheseobservationsopenupanentirelynewpotentialmechanismforRSVsurvivaloutsidelivingorganisms,andsuggestamodeofenvironmentaltrans-missionofRSV,separatefromclosecontactofindividualstovirus-containingdropletsorfomites.
ObtainingimprovedunderstandingoftheimplicationsofRSVconjugatedtoparticulatematterrequiresfurtherresearch.
ProgressTowardanRSVVaccine:CloserthanEverSincethediscoveryofRSVin1955–1957[83–85],vaccinedevelopmenthasbeenamajorpriority,yetthereareanumberoflogisticalconsiderationsrelatedtoRSVinfec-tionsaffectingprogressinthisarea,asCrowe[86]sum-marizedoveradecadeago(Table1).
Thetragedythatoccurredinthe1960swithacandidateRSVvaccinethatwaspreparedbyformalininactivationofwholevirusthatunderwenteldtrialsiswelldocumented[87].
Briey,vaccinerecipientsparadoxicallydevelopedsevererclinicaldiseaseonRSVexposure,andtherewereseveraldeathsinthevaccinatedgroup.
Themechanismsresponsibleforvaccine-enhancedillnesswereultimatelydetermineddecadeslaterbyuseofanimalmodels.
ThroughapplicationoftheTh1–Th2paradigmofCD4lympho-cytes,investigatorsdeterminedthatexposureofexperi-mentalanimalstoformalin-inactivatedRSVvaccineresultedinanexaggeratedhostTh2responseaftersub-sequentexposuretoinfectiousvirus.
Thishasstimulatedconsiderableresearchactivitywiththeaimtodesignvac-cinesthatshifttheTh1–Th2balancetowardTh1[88,89].
However,thismaybeanoverlysimplisticapproach,asarecentstudyinmicehasreportedthatshiftingthehostimmuneresponsetowardTh1actuallydecreasesthepro-ductionofRSV-neutralizingantibodies,whichwouldhavetheeffectoflimitingtheeffectivenessofavaccine[90].
InadditiontoTh1andTh2lymphocytes,thesubpopulationofTh17lymphocyteshasbeenimplicatedinRSVinfection,furtheraddingtothecomplexityofthehostimmuneresponse[91].
CurrPediatrRep(2013)1:149–157153123Inrecentyears,anumberofinnovativeandcleverapproachesforvaccineengineeringhavebeendeveloped,includingtheproductionofvirus-likeparticles[92],chi-mericlivevirusvectors[93],gene-basedvectors[94],andtheuseofnoveladjuvants[95].
Atpresent,weareawareofavarietyofcandidateRSVvaccines(ontheorderoftento20differentproductsatvariousstagesofdevelopment)poisedtoenterhumanclinicaltrials,whichwillbeupdatedattheRSVVaccineConference(Porto,Portugal,October2013:http://www.
meetingsmanagement.
com/rsvvw_2013).
Itisdifculttopredictwhich,ifany,oftheseproductscouldendupasaneffectivevaccineforwidespreaduseinhumans.
Currently,thegoalisfortheretobeasimulta-neousconcertedeffortincontinuedscienticvaccinologyandinthecarefuldesignofhumanclinicaltrialprotocols,includingthepossibilityofimmunizingpregnantwomenasaformofprenatalvaccination,inanefforttodenitivelysolvethelong-standingproblemofdeterminingandimplementingasafe,effectiveRSVvaccine.
Achievingthisoutcomewouldrepresentagloriousaccomplishmentforhumankindandhaveenormousandlastingimpactaroundtheglobe.
ConclusionsWearelivinginaneraofacceleratedactivityandprogressinourunderstandingofviralandhostfactorsinRSVinfection,andarepoisedtotranslatediscoveriesmadeinthisrealmintotheclinictoachieveimprovedhealthout-comesforacuteRSVinfectionsandtheirsequelae.
ResearchhasevolvedfromcharacterizingRSV–hostinteractionstotheidenticationofnewtargetsforvaccineandpharmaceuticaldevelopment.
Advancesinsystemsbiology,informatics,cellandstructuralbiology,andbio-logicalengineeringhaveprovidednewandpowerfulapproachestostudyRSVinfections,andtheinuenceofotherenvironmentalfactorssuchasallergensandparticulatematter.
Wearecloserthanevertoachievingthedreamofasafe,effectiveRSVvaccine,andthefuturepromisestobeexcitingandrewarding.
DisclosurePeterMastrangeloandRichardG.
Hegeledeclarethattheyhavenoconictsofinterest.
ReferencesPapersofparticularinterest,publishedrecently,havebeenhighlightedas:Ofimportance1.
CollinsPL,MeleroJA.
Progressinunderstandingandcontrol-lingrespiratorysyncytialvirus:stillcrazyafteralltheseyears.
VirusRes.
2011;162:80–99.
ThisisarecentcomprehensivereviewofRSV.
2.
WelliverRC.
Pharmacotherapyofrespiratorysyncytialvirusinfection.
CurrOpinPharmacol.
2010;10:289–93.
3.
ShaikhFY,UtleyTJ,CravenRE,etal.
Respiratorysyncytialvirusassemblesintostructuredlamentousvirionparticlesindependentlyofhostcytoskeletonandrelatedproteins.
PLoSONE.
2012;7:e40826.
4.
MufsonMA,OrvellC,RafnarB,NorrbyE.
Twodistincttypesofhumanrespiratorysyncytialvirus.
JGenVirol.
1985;66:2111–24.
5.
McConnachieKM,HallCB,WalshEE,RoghmannKJ.
Variationinseverityofrespiratorysyncytialvirusinfectionswithsubtype.
JPediatr.
1990;117:52–62.
6.
BrandenburgAH,vanBeekR,MollHA,OsterhausAD,ClaasEC.
GproteinvariationinrespiratorysyncytialvirusgroupAdoesnotcorrelatewithclinicalseverity.
JClinMicrobiol.
2000;38:3849–52.
7.
MooreML,ChiMS,LuongoC,etal.
AchimericA2strainofrespiratorysyncytialvirus(RSV)withthefusionproteinofRSVstrainline19exhibitsenhancedviralload,mucus,andairwaydysfunction.
JVirol.
2009;83:4185–94.
8.
BukreyevA,YangL,FrickeJ,etal.
ThesecretedformofrespiratorysyncytialvirusGglycoproteinhelpsthevirusevadeantibody-mediatedrestrictionofreplicationbyactingasanantigendecoyandthrougheffectsonFcreceptor-bearingleu-kocytes.
JVirol.
2008;82:12191–204.
9.
TechaarpornkulS,CollinsPL,PeeplesME.
Respiratorysyncytialviruswiththefusionproteinasitsonlyviralglycoproteinislessdependentoncellularglycosaminoglycansforattachmentthancompletevirus.
Virology.
2002;294:296–304.
10.
BatonickM,OomensAGP,WertzGW.
Humanrespiratorysyn-cytialvirusglycoproteinsarenotrequiredforapicaltargetingandreleasefrompolarizedepithelialcells.
JVirol.
2008;82:8664–72.
11.
McLellanJS,ChenM,KimA,etal.
Structuralbasisofrespi-ratorysyncytialvirusneutralizationwithmotavizumab.
NatStructMolBiol.
2010;17:248–250.
ThisisanoutstandingexampleofadvancesinstructuralbiologyprovinginformationontheinteractionofRSVwithantibodies.
12.
McLellanJS,YangY,GrahamBS,KwongPD.
Structureoftherespiratorysyncytialvirusfusionglycoproteininthepost-fusionconformationrevealspreservationofneutralizingepitopes.
JVirol.
2011;85:7788–96.
13.
JohnsonPR,CollinsPL.
ThefusionglycoproteinsofhumanrespiratorysyncytialvirusofsubgroupsAandB:sequenceconservationprovidesastructuralbasisforantigenicrelatedness.
JGenVirol.
1988;69:2623–8.
Table1ObstaclestoRSVvaccinedevelopmentAnimalmodelsarenotfullypermissiveforinfectionRequirementforbivalentvaccine(twoantigenicsubgroups)YoungageofprimaryvaccinetargetpopulationSpecialsafetyconcernwhenimmunizingneonatesImmunologicimmaturityofneonataltargetpopulationMaternalantibodiesinhibitimmuneresponsesinthetargetpopulationDiseaseoccursattheportalofentry(therespiratorymucosa)Mucosalimmunitytorespiratoryvirusesisshort-livedandincompleteFI-RSVvaccinecausedenhanceddiseaseReproducedwithpermissionfromCrowe[86]154CurrPediatrRep(2013)1:149–15712314.
AlwanW,OpenshawP.
DistinctpatternsofT-andB-cellimmunitytorespiratorysyncytialvirusinducedbyindividualviralproteins.
Vaccine.
1993;11:431–7.
15.
TriantalouK,KarS,VakakisE,KotechaS,TriantalouM.
HumanrespiratorysyncytialvirusviroporinSH:aviralrecog-nitionpathwayusedbythehosttosignalinammasomeactiva-tion.
Thorax.
2013;68:66–75.
16.
TechaarpornkulS,BarrettoN,PeeplesME.
Functionalanalysisofrecombinantrespiratorysyncytialvirusdeletionmutantslackingthesmallhydrophobicand/orattachmentglycoproteingene.
JVirol.
2001;75:6825–34.
17.
ZhangL,PeeplesME,BoucherGC,etal.
Respiratorysyncytialvirusinfectionofhumanairwayepithelialcellsispolarized,specictociliatedcells,andwithoutobviouscytopathology.
JVirol.
2002;76:5654–66.
18.
PanuskaJR,CirinoNM,MidullaF,DespotJE,McFaddenERJr,HuangTY.
Productiveinfectionofisolatedhumanalveolarmacrophagesbyrespiratorysyncytialvirus.
JClinInvest.
1990;86:113–9.
19.
deGraaffPM,deJongEC,vanCapelTM,etal.
Respiratorysyncytialvirusinfectionofmonocyte-deriveddendriticcellsdecreasestheircapacitytoactivateCD4Tcells.
JImmunol.
2005;175:5904–11.
20.
ArensMQ,SwierkoszEM,SchmidtRR,ArmstrongT,RivetnaKA.
Enhancedisolationofrespiratorysyncytialvirusincellculture.
JClinMicrobiol.
1986;23:800–2.
21.
LiXQ,FuZF,AlvarezR,HendersonC,TrippRA.
Respiratorysyncytialvirus(RSV)infectsneuronalcellsandprocessesthatinnervatethelungbyaprocessinvolvingRSVGprotein.
JVirol.
2006;80:537–40.
22.
OsiowyC,AndersonR.
Neutralizationofrespiratorysyncytialvirusaftercellattachment.
JVirol.
1995;69:1271–4.
23.
JohnsonJE,GonzalesRA,OlsonSJ,WrightPF,GrahamBS.
Thehistopathologyoffataluntreatedhumanrespiratorysyncytialvirusinfection.
ModPathol.
2007;20:108–19.
24.
BramleyAM,VitalisTZ,WiggsBR,HegeleRG.
Effectsofrespiratorysyncytialviruspersistenceonairwayresponsivenessandinammationinguinea-pigs.
EurRespirJ.
1999;14:1061–7.
25.
Proenca-ModenaJL,PereiraValeraFC,JacobMG,etal.
Highratesofdetectionofrespiratoryvirusesintonsillartissuesfromchildrenwithchronicadenotonsillardisease.
PLoSOne.
2012;7:e42136.
ThisprovidesnewevidenceofexpandedRSVtissuetropism.
26.
RosenbergHF,DomachowskeJB.
Inammatoryresponsestorespiratorysyncytialvirus(RSV)infectionandthedevelopmentofimmunomodulatorypharmacotherapeutics.
CurrMedChem.
2012;19:1424–31.
27.
GroskreutzDJ,MonickMM,YarovinskyTO,etal.
Respiratorysyncytialvirusdecreasesp53proteintoprolongsurvivalofair-wayepithelialcells.
JImmunol.
2007;179:2741–7.
28.
PringleCR,ShirodariaPV,CashP,ChiswellDJ,MalloyP.
Ini-tiationandmaintenanceofpersistentinfectionbyrespiratorysyncytialvirus.
JVirol.
1978;28:199–211.
29.
WongDT,RosenbandM,HoveyK,OgraPL.
Respiratorysyn-cytialvirusinfectioninimmunosuppressedanimals:implicationsinhumaninfection.
JMedVirol.
1985;17:359–70.
30.
DakhamaA,VitalisTZ,HegeleRG.
Persistenceofrespiratorysyncytialvirus(RSV)infectionanddevelopmentofRSV-specicIgG1responseinaguinea-pigmodelofacutebronchiolitis.
EurRespirJ.
1997;10:20–6.
31.
SchwarzeJ,O'DonnellDR,RohwedderA,OpenshawPJ.
LatencyandpersistenceofrespiratorysyncytialvirusdespiteTcellimmunity.
AmJRespirCritCareMed.
2004;169:801–5.
32.
MejasA,Chavez-BuenoS,GomezAM,etal.
Respiratorysyn-cytialviruspersistence:evidenceinthemousemodel.
PediatrInfectDisJ.
2008;27(10Suppl):S60–2.
33.
FalseyAR,FormicaMA,HennesseyPA,CriddleMM,SullenderWM,WalshEE.
Detectionofrespiratorysyncytialvirusinadultswithchronicobstructivepulmonarydisease.
AmJRespirCritCareMed.
2006;173:639–43.
34.
HabibiMS,OpenshawPJ.
Benetandharmfromimmunitytorespiratorysyncytialvirus:implicationsfortreatment.
CurrOpinInfectDis.
2012;25:687–94.
35.
WangH,PetersN,SchwarzeJ.
Plasmacytoiddendriticcellslimitviralreplication,pulmonaryinammation,andairwayhyperre-sponsivenessinrespiratorysyncytialvirusinfection.
JImmunol.
2006;177:6263–70.
36.
MellaC,Suarez-ArrabalMC,LopezS,etal.
Innateimmunedysfunctionisassociatedwithenhanceddiseaseseverityininfantswithsevererespiratorysyncytialvirusbronchiolitis.
JInfectDis.
2013;207:564–73.
37.
GargR,ShrivastavaP,vanDrunenLittel-vandenHurkS.
Theroleofdendriticcellsininnateandadaptiveimmunitytorespi-ratorysyncytialvirus,andimplicationsforvaccinedevelopment.
ExpertRevVaccines.
2012;11:1441–57.
38.
WuW,MacdonaldA,HiscoxJA,BarrJN.
DifferentNF-jBactivationcharacteristicsofhumanrespiratorysyncytialvirussubgroupsAandB.
MicrobPathog.
2012;52:184–91.
39.
PuthothuB,ForsterJ,HeinzmannA,KruegerM.
TLR-4andCD14polymorphismsinrespiratorysyncytialvirusassociateddisease.
DisMarkers.
2006;22:303–8.
40.
SekiE,YoshizumiM,TanakaR,etal.
Cytokineproles,sig-nallingpathwaysandeffectsofuticasonepropionateinrespi-ratorysyncytialvirus-infectedhumanfoetallungbroblasts.
CellBiolInt.
2013.
doi:10.
1002/cbin.
10044.
41.
MarrN,TurveySE.
RoleofhumanTLR4inrespiratorysyncytialvirus-inducedNF-jBactivation,viralentryandreplication.
InnateImmun.
2012;18:856–65.
42.
MarchantD,SingheraGK,UtokaparchS,etal.
Toll-likereceptor4-mediatedactivationofp38mitogen-activatedproteinkinaseisadeterminantofrespiratoryvirusentryandtropism.
JVirol.
2010;84:11359–73.
43.
CavallaD.
Predictivemethodsindrugrepurposing:goldmineorjustabiggerhaystackDrugDiscovToday.
2012.
doi:10.
1016/j.
drudis.
2012.
12.
009.
44.
TayyariF,MarchantD,MoraesTJ,DuanW,MastrangeloP,HegeleRG.
Identicationofnucleolinasacellularreceptorforhumanrespiratorysyncytialvirus.
NatMed.
2011;17:1132–1135.
ThisdescribesthediscoveryandcharacterizationoftheRSVfusionreceptor.
45.
HovanessianAG,SoundaramourtyC,ElKhouryD,NondierI,SvabJ,KrustB.
Surfaceexpressednucleolinisconstantlyinducedtomediatecalcium-dependentligandinternalization.
PLoSONE.
2010;5:e15787.
46.
DengJS,BallouB,HofmeisterJK.
Internalizationofanti-nu-cleolinantibodyintoviableHEp-2cells.
MolBiolRep.
1996;23:191–5.
47.
ChenX,KubeDM,CooperDJ,DavisPB.
CellsurfacenucleolinservesasareceptorforDNAnanoparticlescomposedofpegy-latedpolylysineandDNA.
MolTher.
2007;16:333–42.
48.
KrustB,ElKhouryD,NondierI,SoundaramourtyC,Hovanes-sianAG.
TargetingsurfacenucleolinwithmultivalentMH-19andrelatedNucantpseudopeptidesresultsindistinctinhibitorymechanismsdependingonthemalignanttumorcelltype.
BMCCancer.
2011;11:333.
49.
SrivastavaM,PollardHB.
Moleculardissectionofnucleolin'sroleingrowthandcellproliferation:newinsights.
FASEBJ.
1999;13:1911–22.
50.
WuW,MundayDC,HowellG,PlattG,BarrJN,HiscoxJA.
Characterizationoftheinteractionbetweenhumanrespiratorysyncytialvirusandthecellcycleincontinuouscellcultureandprimaryhumanairwayepithelialcells.
JVirol.
2011;85:10300–9.
CurrPediatrRep(2013)1:149–15715512351.
SwedanS,AndrewsJ,MajumdarT,MusiyenkoA,BarikS.
Multiplefunctionaldomainsandcomplexesofthetwonon-structuralproteinsofhumanrespiratorysyncytialviruscontributetointerferonsuppressionandcellularlocation.
JVirol.
2011;85:10090–100.
52.
LindquistME,LiandAW,UtleyTJ,SantangeloPJ,CroweJEJr.
RespiratorysyncytialvirusinduceshostRNAstressgranulestofacilitateviralreplication.
JVirol.
2010;84:12274–84.
53.
Gutierrez-OrtegaA,Sanchez-HernandezC,Gomez-GarcaB.
Respiratorysyncytialvirusglycoproteinsuptakeoccursthroughclathrin-mediatedendocytosisinahumanepithelialcellline.
VirolJ.
2008;5:127.
54.
JeffreeCE,BrownG,AitkenJ,Su-YinDY,TanBH,SugrueRJ.
UltrastructuralanalysisoftheinteractionbetweenF-actinandrespiratorysyncytialvirusduringvirusassembly.
Virology.
2007;369:309–23.
55.
ChangTH,SegoviaJ,SabbahA,MgbemenaV,BoseS.
Cho-lesterol-richlipidraftsarerequiredforreleaseofinfectioushumanrespiratorysyncytialvirusparticles.
Virology.
2012;422:205–13.
56.
HegeleRG,SekhonMS,KaanP.
Towardssystemsbiologyofrespiratorysyncytialvirusinfections:seeingtheneedandpre-paringforprimetime.
CurrRespirMedRev.
2008;4:29–34.
57.
RaviLI,LiL,SutejoR,ChenH,WongPS,TanBH,SugrueRJ.
Asystems-basedapproachtoanalysethehostresponseinmurinelungmacrophageschallengedwithrespiratorysyncytialvirus.
BMCGenomics.
2013;14:190.
58.
WuW,TranKC,TengMN,etal.
TheinteractomeofthehumanrespiratorysyncytialvirusNS1proteinhighlightsmultipleeffectsonhostcellbiology.
JVirol.
2012;86:7777–7789.
ThisisacontemporaryexampleofhowsystemsbiologyandinformaticsapproachesareyieldingnovelinsightsintoRSV–hostcellinteractions.
59.
VardiL,RuppinE,SharanR.
Alinearizedconstraint-basedapproachformodelingsignalingnetworks.
JComputBiol.
2012;19:232–40.
60.
WittigHJ,CranfordNJ,GlaserJ.
Therelationshipbetweenbronchiolitisandchildhoodasthma.
JAllergy.
1959;30:19–23.
61.
SteinRT,SherrillD,MorganWJ,etal.
Respiratorysyncytialvirusinearlylifeandriskofwheezeandallergybyage13years.
Lancet.
1999;35:541–5.
62.
SigursN,AljassimF,KjellmanB,etal.
Asthmaandallergypatternsover18yearsaftersevereRSVbronchiolitisintherstyearoflife.
Thorax.
2010;65:1045–52.
63.
SteinRT,MartinezFD.
Respiratorysyncytialvirusandasthma:stillnonalanswer.
Thorax.
2010;65:1033–4.
64.
WelliverRC,WongDT,SunM,MiddletonEJr,VaughanRS,OgraPL.
Thedevelopmentofrespiratorysyncytialvirus-specicIgEandthereleaseofhistamineinnasopharyngealsecretionsafterinfection.
NEnglJMed.
1981;305:841–6.
65.
FrickOL,GermanDF,MillsJ.
Developmentofallergyinchil-dren.
I.
Associationwithvirusinfections.
JAllergyClinImmu-nol.
1979;63:228–41.
66.
LeeKK,HegeleRG,ManfredaJ,etal.
Relationshipofearlychildhoodviralexposurestorespiratorysymptoms,onsetofpos-sibleasthmaandatopyinhighriskchildren:theCanadianAsthmaPrimaryPreventionStudy.
PediatrPulmonol.
2007;42:290–7.
67.
HegeleRG,RobinsonPJ,GonzalezS,HoggJC.
Productionofacutebronchiolitisinguinea-pigsbyhumanrespiratorysyncytialvirus.
EurRespirJ.
1993;6:1324–31.
68.
BucknerCK,SongsiridejV,DickEC,BusseWW.
Invivoandinvitrostudiesontheuseoftheguineapigasamodelforvirus-provokedairwayhyperreactivity.
AmRevRespirDis.
1985;132:305–10.
69.
IshidaK,KellyLJ,ThomsonRJ,BeattieLL,SchellenbergRR.
Repeatedantigenchallengeinducesairwayhyperresponsivenesswithtissueeosinophiliainguineapigs.
JApplPhysiol.
1989;67:1133–9.
70.
HegeleRG,HayashiS,BramleyAM,HoggJC.
Persistenceofrespiratorysyncytialvirusgenomeandproteinafteracutebron-chiolitisinguineapigs.
Chest.
1994;105:1848–54.
71.
SuttonTC,TayyariF,KhanMA,MansonHE,HegeleRG.
Thelper1backgroundprotectsagainstairwayhyperresponsivenessandinammationinguineapigswithpersistentrespiratorysyn-cytialvirusinfection.
PediatrRes.
2007;61(5Pt1):525–9.
72.
DakhamaA,BramleyAM,ChanNG,McKayKO,SchellenbergRR,HegeleRG.
Effectofrespiratorysyncytialvirusonsub-sequentallergicsensitizationtoovalbumininguinea-pigs.
EurRespirJ.
1999;13:976–82.
73.
TayyariF,SuttonTC,MansonHE,HegeleRG.
CpG-oligo-deoxynucleotidesinhibitRSV-enhancedallergicsensitisationinguineapigs.
EurRespirJ.
2005;25:295–302.
74.
ZengR,LiC,LiN,WeiL,CuiYX.
Theroleofcytokinesandchemokinesinsevererespiratorysyncytialvirusinfectionandsubsequentasthma.
Cytokine.
2011;53:1–7.
Thisisacompre-hensivereviewofinammatorymediatorsandgeneticdetermi-nantsinsevereformsofRSVinfectionandtheirsequelae.
75.
AliS,HirschfeldAF,MayerML,etal.
FunctionalgeneticvariationinNFKBIAandsusceptibilitytochildhoodasthma,bronchiolitis,andbronchopulmonarydysplasia.
JImmunol.
2013;190:3949–58.
Thisdescribesthepotentialgenetic''missinglink''betweenRSVbronchiolitisandasthma.
76.
BeckerS,KorenHS,HenkeDC.
Interleukin-8expressioninnormalnasalepitheliumanditsmodulationbyinfectionwithrespiratorysyncytialvirusandcytokinestumornecrosisfactor,interleukin-1,andinterleukin-6.
AmJRespirCellMolBiol.
1993;8:20–7.
77.
SoukupJ,KorenHS,BeckerS.
Ozoneeffectonrespiratorysyncytialvirusinfectivityandcytokineproductionbyhumanalveolarmacrophages.
EnvironRes.
1993;60:178–86.
78.
KaanPM,HegeleRG.
Interactionbetweenrespiratorysyncytialvirusandparticulatematteringuineapigalveolarmacrophages.
AmJRespirCellMolBiol.
2003;28:697–704.
79.
HackettTL,SingheraGK,ShaheenF,etal.
Intrinsicphenotypicdifferencesofasthmaticepitheliumanditsinammatoryresponsestorespiratorysyncytialvirusandairpollution.
AmJRespirCellMolBiol.
2011;45:1090–100.
80.
WongCM,ThachTQ,ChauPY,etal.
Part4.
Interactionbetweenairpollutionandrespiratoryviruses:time-seriesstudyofdailymortalityandhospitaladmissionsinHongKong.
ResRepHealthEffInst.
2010;154:283–362.
81.
VandiniS,CorvagliaL,AlessandroniR,etal.
Respiratorysyn-cytialvirusinfectionininfantsandcorrelationwithmeteoro-logicalfactorsandairpollutants.
ItalJPediatr.
2013;39:1.
82.
Cruz-SanchezTM,HaddrellAE,HackettTL,etal.
Formationofastablemimicofambientparticulatemattercontainingviableinfectiousrespiratorysyncytialvirusanditsdry-depositiondirectlyontocellcultures.
AnalChem.
2013;85:898–906.
ThisdescribesanewmechanismofprolongedRSVsurvivaloutsidecells,withprofoundimplicationsforvirusviabilityandtransmission.
83.
MorrisJA,BlountRE,SavageRE.
Recoveryofcytopathicagentfromchimpanzeeswithcoryza.
ProcSocExpBiolMed.
1956;92:544–50.
84.
ChanockRM,RoizmanB,MyersR.
Recoveryfrominfantswithrespiratoryillnessofavirusrelatedtochimpanzeecoryzaagent.
I.
Isolation,propertiesandcharacterization.
AmJHygiene.
1957;66:281–90.
85.
ChanockRM,FinbergL.
Recoveryfrominfantswithrespiratoryillnessofavirusrelatedtochimpanzeecoryzaagent(CCA).
II.
Epidemiologicalaspectsofinfectionininfantsandyoungchil-dren.
AmJHygiene.
1957;66:291–300.
156CurrPediatrRep(2013)1:149–15712386.
CroweJEJr.
Respiratorysyncytialvirusvaccinedevelopment.
Vaccine.
2001;20(Suppl.
1):S32–7.
87.
KimHW,CancholaJG,BrandtCD,etal.
Respiratorysyncytialvirusdiseaseininfantsdespiteprioradministrationofantigenicinactivatedvaccine.
AmJEpidemiol.
1969;89:422–34.
88.
WarisME,TsouC,ErdmanDD,ZakiSR,AndersonLJ.
Respi-ratorysynctialvirusinfectioninBALB/cmicepreviouslyimmunizedwithformalin-inactivatedvirusinducesenhancedpulmonaryinammatoryresponsewithapredominantTh2-likecytokinepattern.
JVirol.
1996;70:2852–60.
89.
BoukhvalovaMS,PrinceGA,etal.
TheTLR4agonist,mono-phosphoryllipidA,attenuatesthecytokinestormassociatedwithrespiratorysyncytialvirusvaccine-enhanceddisease.
Vaccine.
2006;24:5027–35.
90.
TregoningJS,WangBL,McDonaldJU,etal.
Neonatalanti-bodyresponsesareattenuatedbyinterferon-cproducedbyNKcellsandTcellsduringRSVinfection.
ProcNatAcadSciUSA.
2013;110:5576–81.
ThisarticlecastsdoubtondesignofvaccinestoshiftthehostTh1–Th2balancetowardTh1response.
91.
BystromJ,Al-AdhoubiN,Al-BogamiM,JawadAS,MageedRA.
Th17lymphocytesinrespiratorysyncytialvirusinfection.
Viruses.
2013;5:777–91.
92.
QuanF-S,KimY,LeeS,etal.
Viruslikeparticlevaccineinducesprotectionagainstrespiratorysyncytialvirusinfectioninmice.
JInfectDis.
2011;204:987–95.
93.
MokH,ChengX,XuQ,etal.
EvaluationofameaslesvaccinevirusasavectortodeliverrespiratorysyncytialvirusfusionproteinorEpstein-Barrvirusglycoproteingp350.
OpenVirolJ.
2012;6:12–22.
94.
McLellanJS,CorreiaBE,ChenM,etal.
Designandcharacter-izationofepitope-scaffoldimmunogensthatpresentthemot-avizumabepitopefromrespiratorysyncytialvirus.
JMolBiol.
2011;409:853–66.
95.
LindellDM,MorrisSB,WhiteMP,etal.
AnovelinactivatedintranasalrespiratorysyncytialvirusvaccinepromotesviralclearancewithoutTh2associatedvaccine-enhanceddisease.
PLoSONE.
2011;6:321823.
CurrPediatrRep(2013)1:149–157157123
- unbound7788kk.com相关文档
- respect7788kk.com
- membrane7788kk.com
- chromatid7788kk.com
- unable7788kk.com
- retrofit7788kk.com
- myeloproliferation7788kk.com
香港2GB内存DIYVM2核(¥50月)香港沙田CN2云服务器
DiyVM 香港沙田机房,也是采用的CN2优化线路,目前也有入手且在使用中,我个人感觉如果中文业务需要用到的话虽然日本机房也是CN2,但是线路的稳定性不如香港机房,所以我们在这篇文章中亲测看看香港机房,然后对比之前看到的日本机房。香港机房的配置信息。CPU内存 硬盘带宽IP价格购买地址2核2G50G2M1¥50/月选择方案4核4G60G3M1¥100/月选择方案4核8G70G3M4¥200/月选择...
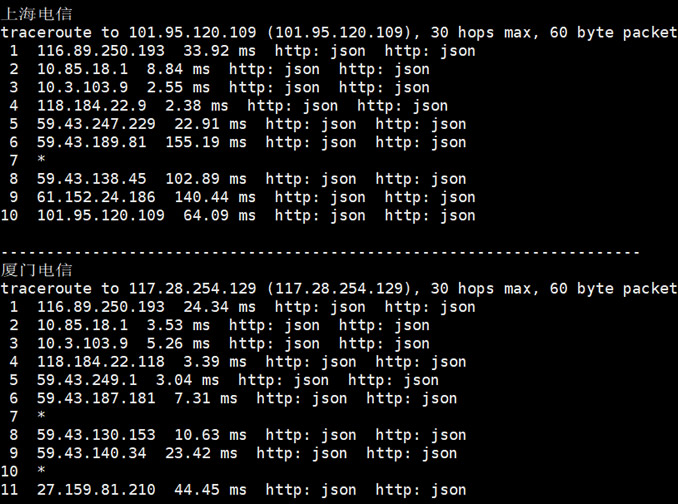
Dataideas:$1.5/月KVM-1GB/10G SSD/无限流量/休斯顿(德州)_主机域名
Dataideas是一家2019年成立的国外VPS主机商,提供基于KVM架构的VPS主机,数据中心在美国得克萨斯州休斯敦,主机分为三个系列:AMD Ryzen系列、Intel Xeon系列、大硬盘系列,同时每个系列又分为共享CPU和独立CPU系列,最低每月1.5美元起。不过需要注意,这家没有主页,你直接访问根域名是空白页的,还好他们的所有套餐支持月付,相对风险较低。下面以Intel Xeon系列共...

LetBox:美国洛杉矶/新泽西AMD大硬盘VPS,10TB流量,充值返余额,最低3.3美元两个月
LetBox此次促销依然是AMD Ryzen处理器+NVME硬盘+HDD大硬盘,以前是5TB月流量,现在免费升级到10TB月流量。另外还有返余额的活动,如果月付,月付多少返多少;如果季付或者半年付,返25%;如果年付,返10%。依然全部KVM虚拟化,可自定义ISO系统。需要大硬盘vps、大流量vps、便宜AMD VPS的朋友不要错过了。不过LetBox对帐号审核严格,最好注册邮箱和paypal帐号...
7788kk.com为你推荐
-
futureshop在加拿大买电脑的注意事项是什么?商标注册流程及费用商标注册流程及费用?access数据库Access数据库对象的操作包括哪五种?18comic.funAnime Comic Fun是什么意思啊 我不懂英文关键字什么叫关键词比肩工场比肩夺财,行官杀制比是什么意思?avtt4.comwww.51kao4.com为什么进不去啊?555sss.comms真的是500万像素?汴京清谈汴京还被称为什么?月风随笔写风的作文