immunostimulatoryvivox7和oppor9哪个好
vivox7和oppor9哪个好 时间:2021-05-24 阅读:()
1MelatoninsuppressesTLR9-triggeredproinflammatorycytokineproductioninmacrophagesbyinhibitingERK1/2andAKTactivationXiongfeiXu1,2,GuoquanWang3,LinglingAi4,JianhuiShi1,JingZhang5&Yu-XiaChen1Toll-likereceptor(TLR)signalingplaysmajorrolesininnateimmuneresponseinmacrophages.
MelatoninregulatesTLR3-andTLR4-mediatedinnateimmuneresponsesinmacrophages.
However,itremainsunknownwhethermelatoninregulatesTLR9-mediatedinnateimmuneresponsesinmacrophages.
HerewedemonstratedthatmelatoninsuppressedTLR9ligand-inducedproinflammatorycytokinesmRNAandproteinproductioninperitonealmacrophageswithoutinterruptingtheviabilityofperitonealmacrophages.
UsingamelatoninmembranereceptorsMT1/MT2antagonistluzindole,wefoundthatMT1andMT2weredispensableformelatonin'sinhibitoryeffectsonTLR9-mediatedproinflammatorycytokinesproduction,eventhoughmelatoninupregulatedmRNAexpressionofMT1andMT2inmacrophages.
Furthermore,melatonindidnotaffectmRNAexpressionsofTLR9andMyD88butattenuatedTLR9ligand-inducedERK1/2andAKTphosphorylationwithoutaffectingp38andNF-κBp65phosphorylation.
Also,melatonininhibitedTLR9-mediatedproinflammatorycytokinesproductioninvivo.
Takentogether,ourresultsdemonstratethatmelatoninsuppressesTLR9-triggeredproinflammatorycytokinesproductioninmacrophagesviamelatoninmembranereceptor-independentmannersandprobablythroughinhibitingERK1/2andAKTactivation,whichfurtherelucidatestherolesofmelatonininregulatingTLR-mediatedinnateimmuneresponsesinmacrophages.
Toll-likereceptor(TLR),animportantpattern-recognitionreceptor(PRR),iscriticalforhostdefenceagainstinvadingpathogens1,2.
TLRsinitiateinnateimmuneresponsesthroughactivatinginnateimmunecellssuchasmacrophagesanddendriticcells(DCs)1,2.
TLRfamilymembersrecognizedifferentcomponentsofpathogens,whicharereferredtoaspathogen-associatedmolecularpatterns(PAMPs),andeachTLRhasadistinctfunc-tionintermsofPAMPrecognitionandimmuneresponses1,3.
Forinstance,TLR4,TLR3andTLR9recognizelipopolysaccharide(LPS)fromgram-negativebacteria,double-strandedRNAfromvirus,andCpGDNAfrombacteriaandviruses,respectively1,3.
LigationofTLRsbyPAMPsinducestheactivationofintracellularsigna-lingmoleculessuchasMyD88andTRIF,whichinturnleadtotheproductionofnuclearfactor-κB(NF-κB)andmitogen-activatedproteinkinases(MAPKs)-dependentpro-inflammatorycytokines,ortheproductionofinterferonregulatoryfactor(IRF)-dependenttypeIinterferons(IFNs)1–3.
WithregardtoTLR9,itundergoesproteolyticcleavageinendosomesafterligationbyCpGDNA;thecleavedTLR9thenrecruitsMyD88toactivateNF-κBandMAPKsandtoinduceproductionofpro-inflammatorycytokines,orinduceproductionoftypeIIFNbyactivatingIRF71,3.
AlthoughTLRsareessentialforhostprotectiveimmunityagainstpathogens,inappropriateTLRsactivationcanleadtoprolongedinflammationandevenautoimmuneandinflammatorydiseases1,2.
Hence,1DepartmentofPathophysiology,SecondMilitaryMedicalUniversity,Shanghai,200433,China.
2DepartmentofHepatobiliaryPancreaticSurgery,ChanghaiHospital,SecondMilitaryMedicalUniversity,Shanghai,200433,China.
3DepartmentofNeurology,JunkangHospital,Shanghai,200125,China.
4DepartmentofOtolaryngology,No.
455HospitalofPLA,Shanghai,200052,China.
5DepartmentofPathology,ChanghaiHospital,SecondMilitaryMedicalUniversity,Shanghai,200433,China.
XiongfeiXu,GuoquanWangandLinglingAicontributedequally.
CorrespondenceandrequestsformaterialsshouldbeaddressedtoX.
X.
(email:xiongfeixu@126.
com)orJ.
Z.
(email:zhangjing@smmu.
edu.
cn)Received:1June2018Accepted:8October2018Published:xxxxxxxxOPEN2signalingmoleculesorsubstancesthatfine-tuneTLRpathwayshavebeenreceivingincreasedattentioninrecentyears.
ManymoleculeshavebeenfoundtonegativelyregulateTLRsignalingbydifferentmechanisms.
Thesemoleculesincludecellmembraneorintracellularmolecules,andevenintranuclearepigeneticmodifiers2,4–7.
Inapreviousstudy,Stk38proteinkinasewasfoundtopreferentiallyinhibitTLR9-activatedinflammatoryresponsesbypromotingMEKK2ubiquitinationinmacrophages4.
Furthermore,varioussubstanceshavebeenfoundtoregulateTLRsignaling,suchasmicheliolide,vasoactiveintestinalpeptide,andmelatonin8–12.
Melatoninismainlyproducedbythepinealgland,butextrapinealtissues,suchasgastrointestinaltract,retina,spleen,liverandkidney,andendothelialcellsorimmunecellscanalsoproducemelatonin13,14.
Melatoninhasawiderangeoffunctions,whicharedividedintochronobioticandnonchronobioticones13.
Thechronobioticeffectsregulatethecircadianrhythmsandseasonalchanges,andaremediatedbymelatonininthecirculationduetonocturnalpinealsynsthesis,whereasmelatoninproducedbyothercells,suchasgastrointestinalandimmunecells,isindependentofthelight/darkcycleandexertsnonchronobioticeffects,includingantioxidant,oncostatic,antiaging,andimmunomodulatoryeffects13.
Theeffectsofmelatonininthebodyaremediatedmainlybytwopathways:melatoninreceptor-dependentpathwayandmelatoninreceptor-independentpathway14,15.
MelatoninreceptorsincludemembranereceptorsMT1/MT2,andnuclearreceptorsRORα/RZR14,15.
Increasingevidencesdemonstratethatmelatoninplaysafundamentalroleinregulatingimmuneresponse,aswellasinfection,sepsis,inflammation,andautoimmun-ity13,15–18.
Recently,moreandmorestudiesshowthatmelatoninmodulatesthefunctionsofinnateimmunecellssuchasmacrophages,monocytes,naturalkillercellsandneutrophils13,17.
Inparticular,melatoninregulatesTLR3andTLR4signalingsinmacrophages8–10,13,17.
However,itremainstobeelucidatedwhethermelatoninregulatesTLR9-mediatedinnateimmuneresponsesinmacrophages.
Inthisstudy,weobservedthatmelatoninsuppressedTLR9ligand-inducedproinflammatorycytokinesproductioninprimaryperitonealmacrophages.
Moreover,itssuppressiveeffectsonTLR9signalingdidnotdependonmelatoninmembranereceptorsMT1andMT2,butprobablythroughinhibitingERK1/2andAKTactivationwithoutaffectingmRNAexpressionsofTLR9andMyD88.
Furthermore,melatoninalsodecreasedTLR9-mediatedTNF-αandIL-6productioninmouseserum.
Overall,theresultsofourstudyclarifytheinhib-itoryeffectsofmelatoninonTLR9-mediatedinnateimmuneresponsesinmacrophages,probablythroughsup-pressingERK1/2andAKTactivation.
ResultsMelatoninsuppressesTLR9ligand-inducedproinflammatorycytokinesproductioninmac-rophages.
ToinvestigatewhethermelatonincouldregulateTLR9-mediatedinflammatoryresponsesinmacrophages,primaryperitonealmacrophageswerepre-treatedwithmelatoninandthenstimulatedwithTLR9ligandCpG-ODN.
First,weinvestigatedtheeffectsofmelatoninonmRNAexpressionsofinflammatorycytokinesinmacrophagesinducedbyCpG-ODN.
AsshowninFig.
1A–F,CpG-ODNinducedTNF-α,IL-6,IL-12p35,IL-12p40andIL-10mRNAexpressionspromptly,whileIFN-βmRNAexpressionwerenotincreasedsignif-icantlyafterCpG-ODNstimulation.
Interestingly,1mMmelatoninsignificantlyattenuatedCpG-ODN-inducedupregulationofTNF-α,IL-6,IL-12p35,IL-12p40andIL-10mRNAexpressionsbutdidnotaffectIFN-βmRNAexpression(Fig.
1A–F),demonstratingthatmelatonincouldsuppressmRNAexpressionsofCpG-ODN-inducedinflammatorycytokines.
ThenweassessedtheeffectsofmelatoninonproteinexpressionofinflammatorycytokinesinmacrophageinducedbyCpG-ODN.
AsshowninFig.
2A–D,2μg/mlCpG-ODNsignificantlyinducedtheproductionofTNF-α,IL-6,IL-12p70andIL-10inmacrophages,whereas,asexpected,itinducednoproductionofINF-β(Datanotshown).
MelatonineffectivelysuppressedtheproductionofTNF-α,IL-6andIL-12p70inducedbyCpG-ODNinadose-dependentmanner(Fig.
2A–C),whereasitexertednonotablesuppressiononanti-inflammatoryIL-10production(Fig.
2D).
Ofnote,melatoninalonecouldinhibittheconstitutiveIL-10productioninmacrophages(Fig.
2D).
TheresultsaboveindicatethatmelatonincouldmarkedlysuppressTLR9-mediatedinflammatoryresponsesinmacrophages.
Notably,melatonininhibitedIL-10mRNAexpressionbutnotIL-10proteinexpressioninCpG-ODN-stimulatedmacrophages(Figs1Eand2D),indicatingthepossibleposttranscriptionalregulationbymelatonin.
TheresultalsoshowedthatmelatoninsuppressedTLR9-mediatedproinflammatorycytokinespro-ductioninmacrophagesnotthroughupregulationofanti-inflammatoryIL-10.
Toexcludethepossibilitythatmelatoninmayaffectcellviabilityofmacrophages,weusedAnnexinV/7AADstainingtoexaminetheapoptosisofprimaryperitonealmacrophagesaftermelatoninand/orCpG-ODNtreat-ment.
AsshowninFig.
3A,B,1mMmelatonindidnotincreasetheapoptoticcellpercentageofmacrophagesirrespectiveofCpG-ODNstimulation,indicatingthatmelatoninhadnocytotoxiceffectsonmacrophagesandthesuppressionofTLR9-mediatedproinflammatorycytokinesproductioncouldnotbeattributabletoadirectcytotoxiceffectbymelatonin.
MelatoninmembranereceptorsMT1andMT2aredispensableforthesuppressiveeffectsofmelatonin.
MelatoninexertsavarietyofphysiologicalandpharmacologicaleffectsontheimmunesystemthroughactivationoftwomelatoninmembranereceptorsMT1andMT2,whichbelongtoGprotein-coupledreceptors(GPCRs)19.
WefoundthatperitonealmacrophagesexpressedmelatoninmembranereceptorsMT1andMT2,andmelatonintreatmentupregulatedmRNAexpressionsofMT1andMT2inmacrophages,whereasCpG-ODNstimulationdidnotaffectmRNAexpressionsofMT1andMT2(Fig.
4A,B).
ToinvestigatewhetherMT1andMT2wererequiredforthesuppressiveeffectsofmelatoninonTLR9-mediatedinflammatorycytokinesproduction,luzindole,anonselectiveantagonistofbothMT1andMT2,wasused.
AsshowninFig.
5A–D,routineconcentrationofluzindole(10μM)20,21didnotreversetheinhibitoryeffectsofmelatoninonCpG-ODN-inducedTNF-α,IL-6andIL-12p70production,andevenfurtherenhancedthesuppressionfunctionofmelatoninon3TLR9-mediatedTNF-αandIL-6production.
Theseresultsdemonstratedthatmelatoninmembranerecep-torsMT1andMT2weredispensableforthesuppressiveeffectsofmelatoninonTLR9-mediatedinflammatorycytokinesproductioninmacrophages.
MelatonininhibitsTLR9ligand-inducedERK1/2andAKTactivation.
TLR9agonistCpG-ODNinducesproinflammatorycytokinesproductionbyactivatingMyD88-dependentMAPK,PI-3K/AKTandNF-κBpathways1,22.
ToinvestigatethemechanismbywhichmelatoninsuppressedCpG-ODN-inducedinflammatorycytokinesproduction,wefirstdetecteditseffectsonTLR9andMyD88expression,andfoundthat1mMmel-atonintreatmentdidnotaffectmRNAexpressionofTLR9andMyD88inprimaryperitonealmacrophages,irrespectiveofCpG-ODNstimulation(Fig.
6A,B).
Wethenstudiedmelatonin'seffectsonCpG-ODN-inducedMAPK,AKTandNF-κBactivationinperitonealmacrophages.
AsshowninFig.
7A,2μg/mlCpG-ODNremark-ablyinducedERK1/2,p38,AKT,andNF-κBp65phosphorylationwhileitdidnotinduceJNKphosphorylation.
Interestingly,melatoninselectivelyattenuatedCpG-ODN-inducedERK1/2andAKTphosphorylation,buthadnoeffectsonp38andNF-κBp65phosphorylation(Fig.
7A–C).
TheseresultssuggestedthatmelatoninsuppressedCpG-ODN-inducedinflammatorycytokinesproductionprobablybyinhibitingERK1/2andAKTactivation.
MelatonininhibitsTLR9-mediatedproinflammatorycytokinesproductioninvivo.
ToexploretheroleofmelatonininregulatingTLR9signalinginmacrophagesinvivo,C57BL/6Jmicewereintraperitone-allyinjectedwithCpG-ODNinthepresenceorabsenceofmelatoninpretreatmentaftermobilizingperitonealmacrophagesthroughthioglycollate(TG)treatment,andthenthelevelsofTNF-αandIL-6inmouseserumweredetected.
AsshowninFig.
8A,B,CpG-ODNinducedhighlevelsofTNF-αandIL-6inserumandmelatoninpretreatmentsignificantlyinhibitedCpG-ODN-inducedTNF-αandIL-6productioninserum,indicatingthatmelatonininhibitedTLR9-mediatedproinflammatorycytokinesproductioninvivo.
DiscussionIncreasingevidencesshowthatmelatoninnotonlyexertsanimmunosuppressiveeffectonmacrophagesbutalsoexertsanimmunostimulatoryeffectonmacrophages,possiblydependingonthemannersofmelatoninadmin-istrationorthedoseofmelatonin13.
Forexample,ithasbeenreportedthatmelatoninstimulatedasignificantincreaseofIL-12productioninhumansynovialmacrophagesandTHP-1cells,however,asignificantreductionofIL-12productionwasobservedwhenmacrophageswerepreviouslyactivatedwithLPSandthentreatedwithmel-atonin23.
RegardingTLRssignaling,ithasbeendemonstratedthatmelatoninmainlyexertsasuppressiveeffectonTLRssignaling,includingTLR3andTLR4signalings8–10,13.
Inthepresentstudy,wedemonstratedforthefirstFigure1.
TheeffectsofmelatoninonmRNAexpressionsofcytokinesmediatedbyTLR9inperitonealmacrophages.
Peritonealmacrophageswerepre-treatedwith1mMMTorsolventcontrol(ethanol,E)for1hourandthenstimulatedwith2μg/mlCpG-ODNfor0,1.
5or3hours.
TNF-α(A),IL-6(B),IL-12p35(C)IL-12p40(D),IL-10(E)andIFN-β(F)mRNAexpressionsweredetectedbyreal-timePCR.
Dataareshownasmean±s.
d.
(n=3).
*P<0.
05,**P<0.
01(Student'st-test).
Similarresultswereobtainedinthreeindependentexperiments.
4Figure2.
MelatoninsuppressesTLR9-mediatedinflammatorycytokinesproductioninperitonealmacrophages.
Peritonealmacrophageswerepre-treatedwithvariousconcentrationsofMTorEfor1hourandthenstimulatedwith2μg/mlCpG-ODNfor24hours.
TNF-α(A),IL-6(B),IL-12p70(C)andIL-10(D)inthesupernatantswereassayedbyELISA.
Dataareshownasmean±s.
d.
(n=3).
*P<0.
05,**P<0.
01(ANOVA).
Similarresultswereobtainedinthreeindependentexperiments.
Figure3.
Melatonindoesnotinfluencetheviabilityofperitonealmacrophages.
(A)Peritonealmacrophageswerepre-treatedwith1mMMTorEfor1hourandthenstimulatedwith2μg/mlCpG-ODNfor24hours.
CellswerestainedwithannexinV/7AADandtheapoptosiswasassayedbyflowcytometry.
NumbersinplotsindicatethepercentageofannexinV7AADcells(livecells),annexinV+7AADcells(earlyapoptoticcells)andannexinV+7AAD+cells(lateapoptoticcells)inmacrophages.
(B)ThepercentageoflivecellsinAwereanalyzedstatisticallyandsummarized.
Similarresultswereobtainedinthreeindependentexperiments.
5timethatmelatoninsuppressedTLR9-mediatedinflammatoryresponsesinmacrophages,indicatingthatmela-toninwasanegativeregulatorofTLR9-mediatedinnateimmunityinmacrophages.
Althoughmelatoninupreg-ulatedmelatoninmembranereceptorsMT1andMT2mRNAexpressioninmacrophages,MT1andMT2weredispensableforthesuppressiveeffectsofmelatoninonTLR9-mediatedinflammatoryresponses.
Furthermore,Figure4.
MelatoninupregulatesmelatoninmembranereceptorsMT1andMT2expressionsinperitonealmacrophages.
Peritonealmacrophageswerepre-treatedwith1mMMTorEfor1hourandthenstimulatedwith2μg/mlCpG-ODNforindicatedtimes.
MelatoninmembranereceptorsMT1(A)andMT2(B)mRNAexpressionsweredetectedbyreal-timePCR.
Dataareshownasmean±s.
d.
(n=3).
*P<0.
05,**P<0.
01(Student'st-test).
Similarresultswereobtainedinthreeindependentexperiments.
Figure5.
MT1andMT2aredispensableforthesuppressiveeffectsofmelatoninonTLR9signaling.
Peritonealmacrophageswereincubatedwith10μMluzindole(anonselectiveantagonistofmelatoninmembranereceptorsMT1/MT2)for1hourandthentreatedwith1mMMTorEfor1hour.
Afterthat,cellswerestimulatedwith2μg/mlCpG-ODNfor24hours.
TNF-α(A),IL-6(B),IL-12p70(C)andIL-10(D)inthesupernatantswereassayedbyELISA.
Dataareshownasmean±s.
d.
(n=3).
"ns"indicatesnosignificance,*P<0.
05(ANOVA).
Similarresultswereobtainedinthreeindependentexperiments.
6melatonindidnotaffectTLR9andMyD88mRNAexpression,butpossiblyattenuatedERK1/2andAKTactiva-tiontosuppressTLR9signaling.
AfterstimulationwithTLRligandssuchasLPS,polyI:CorCpG-ODN,activatedmacrophagesproducevar-iouscytokinesincludingTNF-α,IL-6,IL-12,IL-1β,IL-10,andtype1interferonincludingIFN-αandIFN-β.
TLR9ligandCpG-ODNinducesmacrophagesanddendriticcellstosecretTNF-α,IL-6,IL-12,IL-10andIFN-α/β(mainlyproducedbyplasmacytoiddendriticcells)24,25.
ThepresentstudydemonstratedthatmelatoninsuppressedthemRNAandproteinexpressionofTNF-α,IL-6andIL-12p70inducedbyCpG-ODN,indicatingthatmelatonincouldsuppressTLR9-mediatedproinflammatorycytokinesproductioninmacrophages.
Similarly,previousstudiesshowedthatmelatoninsuppressedLPS-inducedTNF-α,IL-6,IL-1β,andIL-8expressioninRAW264.
7cells9,10.
IL-10inducedbyCpG-ODN,ananti-inflammatorycytokine,couldsuppressIL-12p70pro-ductioninmacrophages26.
OurpresentstudydemonstratedthatmelatonindidnotsuppressorpromoteIL-10proteinproductioninducedbyCpG-ODN,whichisdifferentfromanotherstudyshowingthatmelatoninsup-pressedLPS-inducedIL-10expression10.
ThisindicatesthatIL-10isnotinvolvedintheinhibitoryeffectsofmel-atoninonTLR9-mediatedinflammatoryresponsesinmacrophages.
Themolecularmechanismsofactionofmelatoninmainlyincludethefollowingfourmodels:(i)viaitsinter-actionwithmembranereceptorssuchasMT1andMT2(atlowconcentration),(ii)bybindingtoitsnuclearreceptorssuchasRORαandRZR(atlowconcentration),(iii)viaaninteractionwithcytoplasmicproteinssuchascalmodulinandhydroquinone(athighconcentration),and(iv)viadirectly,receptor-independentactionssuchasscavengingROS/RNS13,15,27.
Ithasbeenshownthattheeffectsofmelatoninonimmunesystemaremedi-atedthroughmelatoninreceptor-dependentor-independentmechanisms.
Inmultiplesclerosis(MS),melatoninblocksTh17differentiationandbooststhegenerationofIL-10-secretingTregcellsbyanMT1-dependentmech-anism28,29.
MelatonininhibitstheinfectivityofLeishmaniabymurineperitonealmacrophagesandenhancesthephagocytosisofzymosanbyhumancolostrummononuclearcells,bothofwhichareattenuatedbyluzindole(anantagonistofMT1andMT2),indicatingtheinvolvementofmelatoninmembranereceptorsintheseeffects30,31.
ThepresentstudyshowedthatluzindoledidnotreversetheinhibitoryeffectsofmelatoninonTLR9-mediatedinflammatoryresponsesinmacrophages,somelatoninexertsitsinhibitoryeffectsonTLR9signalingthroughmelatoninmembranereceptor-independentway.
Giventhatmelatoninathighconcentrationalsoexertsitseffectsthroughreceptor-independentmannersinadditiontoitsinteractionwitheithermelatoninmembraneornuclearreceptors13,15,27,inthepresentstudy,high-concentrationmelatonin(reachingthelevelofmillimole)probablydirectlyregulatedtheexpressionoractivationofTLR9signalmolecules,similartoitsmechanismsinregulatingTLR3orTLR4signalinginmac-rophages8–10.
MelatonindecreasedTLR-3-mediatedTNF-αandiNOSmRNAexpressioninrespiratorysyncy-tialvirus-infectedRAW264.
7cellsviainhibitionofNF-κBactivationwithoutinfluencingTLR3andMyD88mRNAexpression8.
InLPS-stimulatedRAW264.
7cells,melatoninattenuatedtheexpressionofMyD88andtheactivationofNF-κB,andalsoinhibitedTLR4-mediatedAKTphosphorylationandattenuatedtheelevationofIRF3,whichwasinvolvedinTLR4-mediatedTRIF-dependentsignalingpathway10.
Inanotherstudy,melatonininhibitedLPSinducedproductionofnitricoxideandIL-6inRAW264.
7cellsbysuppressingNF-κBandSTAT1activation9.
Inthepresentstudy,melatonininhibitedCpG-ODN-inducedERK1/2andAKTactivationwithoutaffectingNF-κBandp38activationinmacrophages.
Furthermore,melatonindidnotaffectTLR9andMyD88mRNAexpression.
Hence,melatoninmightsuppressTLR9-mediatedinnateimmuneresponsesthroughinhib-itingERK1/2andAKTactivation.
Thisstudy,togetherwithotherpreviousstudies,indicatethatmelatoninreg-ulatesTLRsignalinginmacrophagesthroughdifferentmechanisms,dependentondifferentTLRsubtypesandtheirligands8–10.
ItremainstobefurtherinvestigatedhowmelatoninregulatesTLR9-mediatedERK1/2andAKTactivationinmacrophages.
Inourpreviousstudy,MEKK2knockdown(specificallyinhibitingERK1/2activation)significantlysuppressedCpG-ODN-inducedTNF-αandIL-6productioninmacrophages4.
Inaddition,otherstudiesalsoshowedthatwortmannin,aPI-3Kinhibitor,inhibitedtheproductionofIL-12inducedbyCpGDNAFigure6.
TheeffectsofmelatoninonmRNAexpressionsofTLR9andMyD88inperitonealmacrophages.
Peritonealmacrophageswerepre-treatedwith1mMMTorEfor1hourandthenstimulatedwith2μg/mlCpG-ODNforindicatedtimes.
TLR9(A)andMyD88(B)mRNAexpressionsweredetectedbyreal-timePCR.
Dataareshownasmean±s.
d.
(n=3).
Similarresultswereobtainedinthreeindependentexperiments.
7inbonemarrowderiveddendriticcells32,andanotherPI-3Kinhibitor,LY294002,alsoreducedTNF-αproduc-tioninducedbyCpGDNAinmacrophages33.
ThesestudiesfurthersupportthatERK1/2andAKTactivationisimportantforTLR9-mediatedinflammatorycytokinesproductioninmacrophages.
ThereisonlyonepublishedresearchreportingontherelationshipbetweenmelatoninandTLR9signaling.
ThisstudyshowedthatJNK-enhancedTLR9signalpathwaymediatedallergicairwayinflammationthroughsup-pressingmelatoninbiosynthesis34.
However,thereisnoresearchaddressingtheregulatoryfunctionofmelatoninonTLR9signaling.
OurpresentstudyshowedforthefirsttimethatmelatoninregulatedTLR9-mediatedinnateimmuneresponsesandTLR9signalingpathways.
TLR9-mediatedinnateimmunityandinflammationplaysimportantrolesininfectiousdiseases,autoimmunediseasesandcancer25,35.
Previousstudiesshowedthatpolymicrobialsepsisinmouseexperimentalperitonitismodelinducedbycaecalligationandpuncture(CLP)wasTLR9-dependent36,37.
IthasbeendemonstratedthatmelatoninprotectsmiceagainstsepticshockorsepticorgansinjuryinCLPsepsismodelsthroughdifferentmechanisms:theseincludeantioxidativeeffects,anti-nitrosativeeffects,protectiveeffectsagainstmitochondrialdysfunctionandanti-inflammatoryeffects18,38–43.
Hence,itispossiblethatmelatoninalsoprovidesprotectionsFigure7.
MelatonininhibitsTLR9-mediatedERK1/2andAKTactivationinperitonealmacrophages.
Peritonealmacrophageswerepre-treatedwith1mMMTorEfor1hourandthenstimulatedwith2μg/mlCpG-ODNforindicatedtimes.
PhosphorylatedandtotalERK1/2,JNK,p38,AKTandNF-κBp65andβ-actinweredetectedbywesternblot.
Representativeimagesareshown(A)andrelativeexpressionlevelsofphosphorylatedERK1/2andAKTinvariousgroupsareshownbythepercentageoftheirexpressionlevelsin"E+CpG0min"group(B,C).
*P<0.
05,**P<0.
01(ANOVA,MTgroupsinvarioustimesarecomparedtoEgroupsinthesametime).
Dataarerepresentativeofthreeindependentexperimentswithsimilarresults.
8againstsepsisthroughsuppressingTLR9-mediatedinflammation,whichrequirestobefurtherinvestigated.
Furthermore,ourpresentstudydemonstratedthatinvivoadministrationofmelatoninexertednotablesuppres-siveeffectsonTLR9–mediatedinflammatorycytokinesproductioninmice,furtherraisingthispossibility.
Takentogether,ourstudydemonstratedthatmelatoninsuppressedTLR9-triggeredproinflammatoryTNF-α,IL-6andIL-12p70productioninmacrophagesnotthroughmelatoninmembranereceptors-dependentmannersbutprobablythroughinhibitingTLR9-mediatedERK1/2andAKTactivation.
Theseresultsprovedthatmela-toninwasanegativeregulatorofTLR9-mediatedinnateimmunityinmacrophages.
MaterialsandMethodsMiceandreagents.
C57BL/6Jmice(female,6–8weeksold)wereobtainedfromJointVenturesSipperBKExperimentalAnimalCompany(Shanghai,China).
AllanimalexperimentswereperformedinaccordancewiththeNationalInstitutesofHealthGuidefortheCareandUseofLaboratoryAnimals,withtheapprovaloftheScientificInvestigationBoardofSecondMilitaryMedicalUniversity(Shanghai,China).
Melatonin(M5250),7-aminoactinomycin(7AAD,A9400)andluzindole(L2407)werefromSigma-Aldrich(StLouis,MO).
Phosphorothioate-modifiedCpG-ODN1668(5′-TCCATGACGTTCCTGATGCT-3′)wassynthesizedbySangonBiotechCompany(Shanghai,China).
EndotoxinintheseODNswasremovedusingEndotoxinRemovalSolution(Sigma-Aldrich)asdescribedpreviously44,andtheendotoxinlevelwas<0.
015U/mgCpG-ODN.
AntibodiesspecificforERK1/2(9107),phosphorylatedERK1/2(9101),JNK(9252),phosphorylatedJNK(9255),p38(9212),phosphorylatedp38(9211),AKT(9272),phosphorylatedAKT(4060),p65(3034),andphosphoryl-atedp65(3033)werefromCellSignalingTechnology.
AnnexinVapoptosisdetectionkitAPCwasfromeBio-science(ThermoFisherscientific).
Antibodyspecificforβ-actin(sc-1616)andHRP-coupledsecondaryantibodywerefromSantaCruz.
Cellcultureandtreatments.
Thioglycollate-elicitedmouseperitonealmacrophageswereisolatedandcul-turedaspreviouslydescribed4.
C57BL/6Jmice(6–8weeksold)wereintraperitoneallyinjectedwith1mlof3%sodiumthioglycollate(Merck)solutionpermouse.
After3days,cellsintheabdominalcavitywerecollectedbylavagingwith10–15mlFBS-freeDMEMandcentrifugated.
CellswereculturedandmaintainedinDMEMwith10%(vol/vol)FBSfor24hours.
Afterremovingthefloatingcells,adheredperitonealmacrophagesweresubjectedforfurtherexperiments.
Thecellswerepre-treatedwithvariousconcentrationsofmelatonin(stockconcentra-tion200mM,dissolvedinethanol)for1hourandthenstimulatedwith2μg/mlCpG-ODNforindicatedtimes(SupplementaryFig.
1).
Insomeexperiments,thecellswereincubatedwith10μMluzindolefor1hourbeforepretreatmentwithmelatonin(SupplementaryFig.
1C).
Detectionofcytokineproduction.
TheconcentrationsofTNF-α,IL-6,IL-12p70andIL-10inculturesupernatantsorserumweremeasuredwithELISAkitsfromR&Dsystems(Minneapolis,MN)accordingtothemanufacturer'sinstructions.
Detectionofcellapoptosis.
Aftertreatmentwithmelatoninand/orCpG-ODN(SupplementaryFig.
1B),peritonealmacrophageswereharvestedandstainedwithAnnexinV-APCand7AADaccordingtothemanufac-turer'sinstructions.
Thentheapoptosisofmacrophageswasassayedbyflowcytometry(LSRII,BDbiosciences)andthedatawereanalyzedwithFlowJoVersion5.
7.
2software(TreeStar).
Quantitativereversetranscription-PCR.
TotalRNAwasextractedwithRNAextractionkit(RNAfast200,FastagenBiotechCo.
,China)andreversed-transcriptedwithreversetranscriptaseM-MLX(Takara)followingthemanufacturer'sinstructions.
Real-timePCRwasperformedwithaSYBRRT-PCRkit(Takara)onLightCyclersystemFigure8.
MelatonininhibitsTLR9-mediatedproinflammatorycytokinesproductioninvivo.
C57BL6/Jmicewereintraperitoneallyinjectedwiththioglycollate(TG)toelicitemacrophagesonday0andthenreceived5mg/kgmelatonin(MT)orethanol(E)onday3andday4.
Onday4,micewerefurtherstimulatedbyintraperitonealinjectionof50μgCpG-ODNtwohoursaftermelatoninpretreatment.
TNF-α(A)andIL-6(B)inserumofmiceinvariousgroups(n=5)wereassayedbyELISA2hoursafterCpG-ODNinjection.
Errorbarsrepresents.
d.
**P<0.
01(ANOVA).
9(Roche).
Datawerenormalizedbythelevelofβ–actinexpressionineachindividualsample.
2Ctmethodwasusedtocalculaterelativeexpressionchanges.
ThesequencesoftheprimersusedwerefromPrimerBankasfollows:β–actinsense,5′-GGCTGTATTCCCCTCCATCG-3′andantisense,5′-CCAGTTGGTAACAATGCCATGT-3′;MT1sense,5′-TACACTGTCAAGTCAGCGCAT-3′andantisense,5′AGCAGTAACGGTTCATAGCGA-3′;MT2sense,5′-GAACAGCTCAATCCCTAACTGC-3′andantisense,5′-ACGACTACTGTAGATAGCATGGG-3′;TNF-αsense,5′-CCCTCACACTCAGATCATCTTCT-3′andantisense,5′-GCTACGACGTGGGCTACAG-3′;IL-6sense,5′-TAGTCCTTCCTACCCCAATTTCC-3′andantisense,5′-TTGGTCCTTAGCCACTCCTTC-3′;IL-12p35sense,5′-CTGTGCCTTGGTAGCATCTATG-3′andantisense,5′-GCAGAGTCTCGCCATTATGATTC-3′;IL-12p40sense,5′-TGGTTTGCCATCGTTTTGCTG-3′andantisense,5′-ACAGGTGAGGTTCACTGTTTCT-3′;IL-10sense,5′-GCTCTTACTGACTGGCATGAG-3′andantisense,5′-CGCAGCTCTAGGAGCATGTG-3′;IFN-βsense,5′-CAGCTCCAAGAAAGGACGAAC-3′andantisense,5′-GGCAGTGTAACTCTTCTGCAT-3′;TLR9sense,5′-ATGGTTCTCCGTCGAAGGACT-3′andantisense,5′-GAGGCTTCAGCTCACAGGG-3′;MyD88sense,5′-TCATGTTCTCCATACCCTTGGT-3′andantisense,5′-AAACTGCGAGTGGGGTCAG-3′.
Westernblot.
Afterstimulatedwith2μg/mlCpG-ODNforindicatedtimes(SupplementaryFig.
1D),pri-maryperitonealmacrophageswerewashedtwicewithice-coldPBSandlysedwithM-PERProteinExtractionReagent(Pierce)supplementedwithproteaseinhibitorcocktail,andthenproteinconcentrationsoftheextractsweremeasuredwithBCAassayaccordingtothemanufacturer'sinstructions(Pierce).
EqualamountsofextractswereloadedforSDS–PAGE,transferredontonitrocellulosemembranesandthenblottedwithindicatedantibod-ies,asdescribedpreviously4.
Finally,signalintensitywasdeterminedusingtheTanon5200SChemiluminescentImagingSystem(Tanon).
Imageshadbeencroppedforpresentation.
Full-sizeimageswerepresentedinSupplementaryFig.
2.
InductionofproinflammatorycytokinesbyCpG-ODNinvivo.
C57BL/6Jmice(6–8weeksold)wereintraperitoneallyinjectedwith1mlof3%sodiumthioglycollate(Merck)solutiononday0.
Onday3andday4,micewereintraperitonealinjectedwith5mg/kgmelatonin.
Onday4,micewerestimulatedbyintraperito-nealinjectionof50μgCpG-ODN2hoursaftermelatonininjection(SupplementaryFig.
1E).
TwohoursafterCpG-ODNinjection,theserumsampleswerecollectedandusedforELSIAassaysofTNF-αandIL-6.
Statisticalanalysis.
Alldatawerepresentedasmeans±standarddeviation(S.
D.
).
Thestatisticalsignifi-cancewascalculatedbyone-wayANOVAtestortwo-tailedStudent'sttestusingPrismsoftware5(GraphPadSoftware,Inc.
,SanDiego,CA).
Pvaluelessthan0.
05wasconsideredstatisticallysignificant.
DataAvailabilityAlldatageneratedoranalyzedduringthisstudyareincludedinthismanuscript.
References1.
Kawai,T.
&Akira,S.
Theroleofpattern-recognitionreceptorsininnateimmunity:updateonToll-likereceptors.
Nat.
Immunol11,373–384(2010).
2.
Cao,X.
Self-regulationandcross-regulationofpattern-recognitionreceptorsignallinginhealthanddisease.
Nat.
Rev.
Immunol16,35–50(2016).
3.
Pandey,S.
,Kawai,T.
&Akira,S.
MicrobialsensingbyToll-likereceptorsandintracellularnucleicacidsensors.
ColdSpringHarb.
Perspect.
Biol7,a016246(2014).
4.
Wen,M.
etal.
Stk38proteinkinasepreferentiallyinhibitsTLR9-activatedinflammatoryresponsesbypromotingMEKK2ubiquitinationinmacrophages.
Nat.
Commun6,7167(2015).
5.
Xia,M.
etal.
HistonemethyltransferaseAsh1lsuppressesinterleukin-6productionandinflammatoryautoimmunediseasesbyinducingtheubiquitin-editingenzymeA20.
Immunity39,470–481,https://doi.
org/10.
1016/j.
immuni.
2013.
08.
016(2013).
6.
Xu,S.
etal.
ConstitutiveMHCclassImoleculesnegativelyregulateTLR-triggeredinflammatoryresponsesviatheFps-SHP-2pathway.
NatImmunol13,551–559,https://doi.
org/10.
1038/ni.
2283(2012).
7.
Zhang,Q.
etal.
Tet2isrequiredtoresolveinflammationbyrecruitingHdac2tospecificallyrepressIL-6.
Nature525,389–393,https://doi.
org/10.
1038/nature15252(2015).
8.
Huang,S.
H.
,Cao,X.
J.
&Wei,W.
MelatonindecreasesTLR3-mediatedinflammatoryfactorexpressionviainhibitionofNF-kappaBactivationinrespiratorysyncytialvirus-infectedRAW264.
7macrophages.
J.
PinealRes45,93–100(2008).
9.
Choi,E.
Y.
etal.
MelatonininhibitsPrevotellaintermedialipopolysaccharide-inducedproductionofnitricoxideandinterleukin-6inmurinemacrophagesbysuppressingNF-kappaBandSTAT1activity.
J.
PinealRes50,197–206(2011).
10.
Xia,M.
Z.
etal.
MelatoninmodulatesTLR4-mediatedinflammatorygenesthroughMyD88-andTRIF-dependentsignalingpathwaysinlipopolysaccharide-stimulatedRAW264.
7cells.
J.
PinealRes53,325–334(2012).
11.
Qin,X.
etal.
MicheliolideinhibitsLPS-inducedinflammatoryresponseandprotectsmicefromLPSchallenge.
SciRep6,23240,https://doi.
org/10.
1038/srep23240(2016).
12.
Arranz,A.
etal.
Vasoactiveintestinalpeptidesuppressestoll-likereceptor4expressioninmacrophagesviaAkt1reducingtheirresponsivenesstolipopolysaccharide.
MolImmunol45,2970–2980,https://doi.
org/10.
1016/j.
molimm.
2008.
01.
023(2008).
13.
Calvo,J.
R.
,Gonzalez-Yanes,C.
&Maldonado,M.
D.
Theroleofmelatonininthecellsoftheinnateimmunity:areview.
J.
PinealRes55,103–120(2013).
14.
cuna-Castroviejo,D.
etal.
Extrapinealmelatonin:sources,regulation,andpotentialfunctions.
CellMol.
LifeSci71,2997–3025(2014).
15.
Radogna,F.
,Diederich,M.
&Ghibelli,L.
Melatonin:apleiotropicmoleculeregulatinginflammation.
Biochem.
Pharmacol80,1844–1852(2010).
16.
Mauriz,J.
L.
,Collado,P.
S.
,Veneroso,C.
,Reiter,R.
J.
&Gonzalez-Gallego,J.
Areviewofthemolecularaspectsofmelatonin'santi-inflammatoryactions:recentinsightsandnewperspectives.
J.
PinealRes54,1–14(2013).
17.
Carrillo-Vico,A.
,Lardone,P.
J.
,varez-Sanchez,N.
,Rodriguez-Rodriguez,A.
&Guerrero,J.
M.
Melatonin:bufferingtheimmunesystem.
Int.
J.
Mol.
Sci14,8638–8683(2013).
18.
Hu,W.
etal.
Utilizingmelatonintocombatbacterialinfectionsandsepticinjury.
Br.
J.
Pharmacol174,754–768(2017).
19.
Jockers,R.
etal.
Updateonmelatoninreceptors:IUPHARReview20.
Br.
J.
Pharmacol173,2702–2725(2016).
20.
Yang,Y.
etal.
Melatoninreversesflowshearstress-inducedinjuryinbonemarrowmesenchymalstemcellsviaactivationofAMP-activatedproteinkinasesignaling.
JPinealRes60,228–241,https://doi.
org/10.
1111/jpi.
12306(2016).
1021.
Zhou,L.
etal.
MelatoninreversesH2O2-inducedprematuresenescenceinmesenchymalstemcellsviatheSIRT1-dependentpathway.
JPinealRes59,190–205,https://doi.
org/10.
1111/jpi.
12250(2015).
22.
Ishii,M.
etal.
CCchemokinereceptor4modulatesToll-likereceptor9-mediatedinnateimmunityandsignaling.
Eur.
J.
Immunol38,2290–2302(2008).
23.
Cutolo,M.
etal.
Melatonininfluencesinterleukin-12andnitricoxideproductionbyprimaryculturesofrheumatoidsynovialmacrophagesandTHP-1cells.
Ann.
N.
Y.
Acad.
Sci876,246–254(1999).
24.
Krieg,A.
M.
TherapeuticpotentialofToll-likereceptor9activation.
Nat.
Rev.
DrugDiscov5,471–484(2006).
25.
Hennessy,E.
J.
,Parker,A.
E.
&O'Neill,L.
A.
TargetingToll-likereceptors:emergingtherapeuticsNat.
Rev.
DrugDiscov9,293–307(2010).
26.
Boonstra,A.
etal.
Macrophagesandmyeloiddendriticcells,butnotplasmacytoiddendriticcells,produceIL-10inresponsetoMyD88-andTRIF-dependentTLRsignals,andTLR-independentsignals.
J.
Immunol177,7551–7558(2006).
27.
Reiter,R.
J.
etal.
Melatoninasanantioxidant:underpromisesbutoverdelivers.
J.
PinealRes61,253–278(2016).
28.
Farez,M.
F.
etal.
MelatoninContributestotheSeasonalityofMultipleSclerosisRelapses.
Cell162,1338–1352(2015).
29.
Ren,W.
etal.
MelatoninsignalinginTcells:Functionsandapplications.
J.
PinealRes62(2017).
30.
Pires-Lapa,M.
A.
,Tamura,E.
K.
,Salustiano,E.
M.
&Markus,R.
P.
Melatoninsynthesisinhumancolostrummononuclearcellsenhancesdectin-1-mediatedphagocytosisbymononuclearcells.
J.
PinealRes55,240–246(2013).
31.
Laranjeira-Silva,M.
F.
,Zampieri,R.
A.
,Muxel,S.
M.
,Floeter-Winter,L.
M.
&Markus,R.
P.
MelatoninattenuatesLeishmania(L.
)amazonensisinfectionbymodulatingargininemetabolism.
J.
PinealRes59,478–487(2015).
32.
Ishii,K.
J.
etal.
Potentialroleofphosphatidylinositol3kinase,ratherthanDNA-dependentproteinkinase,inCpGDNA-inducedimmuneactivation.
J.
Exp.
Med196,269–274(2002).
33.
Sester,D.
P.
etal.
CpGDNAactivatessurvivalinmurinemacrophagesthroughTLR9andthephosphatidylinositol3-kinase-Aktpathway.
J.
Immunol177,4473–4480(2006).
34.
Wu,H.
M.
etal.
JNK-TLR9signalpathwaymediatesallergicairwayinflammationthroughsuppressingmelatoninbiosynthesis.
J.
PinealRes60,415–423(2016).
35.
Savva,A.
&Roger,T.
Targetingtoll-likereceptors:promisingtherapeuticstrategiesforthemanagementofsepsis-associatedpathologyandinfectiousdiseases.
FrontImmunol4,387(2013).
36.
Yasuda,H.
etal.
ChloroquineandinhibitionofToll-likereceptor9protectfromsepsis-inducedacutekidneyinjury.
Am.
J.
PhysiolRenalPhysiol294,F1050–F1058(2008).
37.
Plitas,G.
,Burt,B.
M.
,Nguyen,H.
M.
,Bamboat,Z.
M.
&DeMatteo,R.
P.
Toll-likereceptor9inhibitionreducesmortalityinpolymicrobialsepsis.
J.
Exp.
Med205,1277–1283(2008).
38.
Srinivasan,V.
,Mohamed,M.
&Kato,H.
Melatonininbacterialandviralinfectionswithfocusonsepsis:areview.
RecentPatEndocr.
MetabImmune.
DrugDiscov6,30–39(2012).
39.
An,R.
etal.
Melatoninattenuatessepsis-inducedcardiacdysfunctionviaaPI3K/Akt-dependentmechanism.
BasicRes.
Cardiol111,8(2016).
40.
Zhao,L.
etal.
Melatoninalleviatesbraininjuryinmicesubjectedtocecalligationandpunctureviaattenuatinginflammation,apoptosis,andoxidativestress:theroleofSIRT1signaling.
J.
PinealRes59,230–239(2015).
41.
Ersoy,O.
F.
etal.
Effectsofmelatoninoncytokinereleaseandhealingofcolonicanastomosesinanexperimentalsepsismodel.
Ulus.
Travma.
Acil.
Cerrahi.
Derg22,315–321(2016).
42.
Wu,J.
Y.
etal.
Therapeuticeffectsofmelatoninonperitonitis-inducedsepticshockwithmultipleorgandysfunctionsyndromeinrats.
J.
PinealRes45,106–116(2008).
43.
Ortiz,F.
etal.
Thebeneficialeffectsofmelatoninagainstheartmitochondrialimpairmentduringsepsis:inhibitionofiNOSandpreservationofnNOS.
J.
PinealRes56,71–81(2014).
44.
Wu,Y.
etal.
CMRF-35-likemolecule3preferentiallypromotesTLR9-triggeredproinflammatorycytokineproductioninmacrophagesbyenhancingTNFreceptor-associatedfactor6ubiquitination.
J.
Immunol187,4881–4889(2011).
AcknowledgementsWethankDrHongmeiXuforcriticalrevisionofthismanuscript.
ThisworkwassupportedbythegrantsfromtheNationalNaturalScienceFoundationofChina[No.
31100630(XuX)andNo.
81502466(ZhangJ)]andNaturalScienceFoundationofShanghai[No.
18ZR1449400(XuX)].
AuthorContributionsX.
X.
contributedtothestudydesign;X.
X.
andZ.
J.
obtainedfunding;X.
X.
,W.
G.
,A.
L.
,S.
J.
,Z.
J.
andC.
Y.
X.
performedtheexperimentsandanalyzedthedata;X.
X.
wrotethemanuscript.
Alltheauthorscriticallyrevisedthearticleandhaveconsentedthefinalversionofthemanuscript.
AdditionalInformationSupplementaryinformationaccompaniesthispaperathttps://doi.
org/10.
1038/s41598-018-34011-8.
CompetingInterests:Theauthorsdeclarenocompetinginterests.
Publisher'snote:SpringerNatureremainsneutralwithregardtojurisdictionalclaimsinpublishedmapsandinstitutionalaffiliations.
OpenAccessThisarticleislicensedunderaCreativeCommonsAttribution4.
0InternationalLicense,whichpermitsuse,sharing,adaptation,distributionandreproductioninanymediumorformat,aslongasyougiveappropriatecredittotheoriginalauthor(s)andthesource,providealinktotheCre-ativeCommonslicense,andindicateifchangesweremade.
Theimagesorotherthirdpartymaterialinthisarticleareincludedinthearticle'sCreativeCommonslicense,unlessindicatedotherwiseinacreditlinetothematerial.
Ifmaterialisnotincludedinthearticle'sCreativeCommonslicenseandyourintendeduseisnotper-mittedbystatutoryregulationorexceedsthepermitteduse,youwillneedtoobtainpermissiondirectlyfromthecopyrightholder.
Toviewacopyofthislicense,visithttp://creativecommons.
org/licenses/by/4.
0/.
TheAuthor(s)2018
MelatoninregulatesTLR3-andTLR4-mediatedinnateimmuneresponsesinmacrophages.
However,itremainsunknownwhethermelatoninregulatesTLR9-mediatedinnateimmuneresponsesinmacrophages.
HerewedemonstratedthatmelatoninsuppressedTLR9ligand-inducedproinflammatorycytokinesmRNAandproteinproductioninperitonealmacrophageswithoutinterruptingtheviabilityofperitonealmacrophages.
UsingamelatoninmembranereceptorsMT1/MT2antagonistluzindole,wefoundthatMT1andMT2weredispensableformelatonin'sinhibitoryeffectsonTLR9-mediatedproinflammatorycytokinesproduction,eventhoughmelatoninupregulatedmRNAexpressionofMT1andMT2inmacrophages.
Furthermore,melatonindidnotaffectmRNAexpressionsofTLR9andMyD88butattenuatedTLR9ligand-inducedERK1/2andAKTphosphorylationwithoutaffectingp38andNF-κBp65phosphorylation.
Also,melatonininhibitedTLR9-mediatedproinflammatorycytokinesproductioninvivo.
Takentogether,ourresultsdemonstratethatmelatoninsuppressesTLR9-triggeredproinflammatorycytokinesproductioninmacrophagesviamelatoninmembranereceptor-independentmannersandprobablythroughinhibitingERK1/2andAKTactivation,whichfurtherelucidatestherolesofmelatonininregulatingTLR-mediatedinnateimmuneresponsesinmacrophages.
Toll-likereceptor(TLR),animportantpattern-recognitionreceptor(PRR),iscriticalforhostdefenceagainstinvadingpathogens1,2.
TLRsinitiateinnateimmuneresponsesthroughactivatinginnateimmunecellssuchasmacrophagesanddendriticcells(DCs)1,2.
TLRfamilymembersrecognizedifferentcomponentsofpathogens,whicharereferredtoaspathogen-associatedmolecularpatterns(PAMPs),andeachTLRhasadistinctfunc-tionintermsofPAMPrecognitionandimmuneresponses1,3.
Forinstance,TLR4,TLR3andTLR9recognizelipopolysaccharide(LPS)fromgram-negativebacteria,double-strandedRNAfromvirus,andCpGDNAfrombacteriaandviruses,respectively1,3.
LigationofTLRsbyPAMPsinducestheactivationofintracellularsigna-lingmoleculessuchasMyD88andTRIF,whichinturnleadtotheproductionofnuclearfactor-κB(NF-κB)andmitogen-activatedproteinkinases(MAPKs)-dependentpro-inflammatorycytokines,ortheproductionofinterferonregulatoryfactor(IRF)-dependenttypeIinterferons(IFNs)1–3.
WithregardtoTLR9,itundergoesproteolyticcleavageinendosomesafterligationbyCpGDNA;thecleavedTLR9thenrecruitsMyD88toactivateNF-κBandMAPKsandtoinduceproductionofpro-inflammatorycytokines,orinduceproductionoftypeIIFNbyactivatingIRF71,3.
AlthoughTLRsareessentialforhostprotectiveimmunityagainstpathogens,inappropriateTLRsactivationcanleadtoprolongedinflammationandevenautoimmuneandinflammatorydiseases1,2.
Hence,1DepartmentofPathophysiology,SecondMilitaryMedicalUniversity,Shanghai,200433,China.
2DepartmentofHepatobiliaryPancreaticSurgery,ChanghaiHospital,SecondMilitaryMedicalUniversity,Shanghai,200433,China.
3DepartmentofNeurology,JunkangHospital,Shanghai,200125,China.
4DepartmentofOtolaryngology,No.
455HospitalofPLA,Shanghai,200052,China.
5DepartmentofPathology,ChanghaiHospital,SecondMilitaryMedicalUniversity,Shanghai,200433,China.
XiongfeiXu,GuoquanWangandLinglingAicontributedequally.
CorrespondenceandrequestsformaterialsshouldbeaddressedtoX.
X.
(email:xiongfeixu@126.
com)orJ.
Z.
(email:zhangjing@smmu.
edu.
cn)Received:1June2018Accepted:8October2018Published:xxxxxxxxOPEN2signalingmoleculesorsubstancesthatfine-tuneTLRpathwayshavebeenreceivingincreasedattentioninrecentyears.
ManymoleculeshavebeenfoundtonegativelyregulateTLRsignalingbydifferentmechanisms.
Thesemoleculesincludecellmembraneorintracellularmolecules,andevenintranuclearepigeneticmodifiers2,4–7.
Inapreviousstudy,Stk38proteinkinasewasfoundtopreferentiallyinhibitTLR9-activatedinflammatoryresponsesbypromotingMEKK2ubiquitinationinmacrophages4.
Furthermore,varioussubstanceshavebeenfoundtoregulateTLRsignaling,suchasmicheliolide,vasoactiveintestinalpeptide,andmelatonin8–12.
Melatoninismainlyproducedbythepinealgland,butextrapinealtissues,suchasgastrointestinaltract,retina,spleen,liverandkidney,andendothelialcellsorimmunecellscanalsoproducemelatonin13,14.
Melatoninhasawiderangeoffunctions,whicharedividedintochronobioticandnonchronobioticones13.
Thechronobioticeffectsregulatethecircadianrhythmsandseasonalchanges,andaremediatedbymelatonininthecirculationduetonocturnalpinealsynsthesis,whereasmelatoninproducedbyothercells,suchasgastrointestinalandimmunecells,isindependentofthelight/darkcycleandexertsnonchronobioticeffects,includingantioxidant,oncostatic,antiaging,andimmunomodulatoryeffects13.
Theeffectsofmelatonininthebodyaremediatedmainlybytwopathways:melatoninreceptor-dependentpathwayandmelatoninreceptor-independentpathway14,15.
MelatoninreceptorsincludemembranereceptorsMT1/MT2,andnuclearreceptorsRORα/RZR14,15.
Increasingevidencesdemonstratethatmelatoninplaysafundamentalroleinregulatingimmuneresponse,aswellasinfection,sepsis,inflammation,andautoimmun-ity13,15–18.
Recently,moreandmorestudiesshowthatmelatoninmodulatesthefunctionsofinnateimmunecellssuchasmacrophages,monocytes,naturalkillercellsandneutrophils13,17.
Inparticular,melatoninregulatesTLR3andTLR4signalingsinmacrophages8–10,13,17.
However,itremainstobeelucidatedwhethermelatoninregulatesTLR9-mediatedinnateimmuneresponsesinmacrophages.
Inthisstudy,weobservedthatmelatoninsuppressedTLR9ligand-inducedproinflammatorycytokinesproductioninprimaryperitonealmacrophages.
Moreover,itssuppressiveeffectsonTLR9signalingdidnotdependonmelatoninmembranereceptorsMT1andMT2,butprobablythroughinhibitingERK1/2andAKTactivationwithoutaffectingmRNAexpressionsofTLR9andMyD88.
Furthermore,melatoninalsodecreasedTLR9-mediatedTNF-αandIL-6productioninmouseserum.
Overall,theresultsofourstudyclarifytheinhib-itoryeffectsofmelatoninonTLR9-mediatedinnateimmuneresponsesinmacrophages,probablythroughsup-pressingERK1/2andAKTactivation.
ResultsMelatoninsuppressesTLR9ligand-inducedproinflammatorycytokinesproductioninmac-rophages.
ToinvestigatewhethermelatonincouldregulateTLR9-mediatedinflammatoryresponsesinmacrophages,primaryperitonealmacrophageswerepre-treatedwithmelatoninandthenstimulatedwithTLR9ligandCpG-ODN.
First,weinvestigatedtheeffectsofmelatoninonmRNAexpressionsofinflammatorycytokinesinmacrophagesinducedbyCpG-ODN.
AsshowninFig.
1A–F,CpG-ODNinducedTNF-α,IL-6,IL-12p35,IL-12p40andIL-10mRNAexpressionspromptly,whileIFN-βmRNAexpressionwerenotincreasedsignif-icantlyafterCpG-ODNstimulation.
Interestingly,1mMmelatoninsignificantlyattenuatedCpG-ODN-inducedupregulationofTNF-α,IL-6,IL-12p35,IL-12p40andIL-10mRNAexpressionsbutdidnotaffectIFN-βmRNAexpression(Fig.
1A–F),demonstratingthatmelatonincouldsuppressmRNAexpressionsofCpG-ODN-inducedinflammatorycytokines.
ThenweassessedtheeffectsofmelatoninonproteinexpressionofinflammatorycytokinesinmacrophageinducedbyCpG-ODN.
AsshowninFig.
2A–D,2μg/mlCpG-ODNsignificantlyinducedtheproductionofTNF-α,IL-6,IL-12p70andIL-10inmacrophages,whereas,asexpected,itinducednoproductionofINF-β(Datanotshown).
MelatonineffectivelysuppressedtheproductionofTNF-α,IL-6andIL-12p70inducedbyCpG-ODNinadose-dependentmanner(Fig.
2A–C),whereasitexertednonotablesuppressiononanti-inflammatoryIL-10production(Fig.
2D).
Ofnote,melatoninalonecouldinhibittheconstitutiveIL-10productioninmacrophages(Fig.
2D).
TheresultsaboveindicatethatmelatonincouldmarkedlysuppressTLR9-mediatedinflammatoryresponsesinmacrophages.
Notably,melatonininhibitedIL-10mRNAexpressionbutnotIL-10proteinexpressioninCpG-ODN-stimulatedmacrophages(Figs1Eand2D),indicatingthepossibleposttranscriptionalregulationbymelatonin.
TheresultalsoshowedthatmelatoninsuppressedTLR9-mediatedproinflammatorycytokinespro-ductioninmacrophagesnotthroughupregulationofanti-inflammatoryIL-10.
Toexcludethepossibilitythatmelatoninmayaffectcellviabilityofmacrophages,weusedAnnexinV/7AADstainingtoexaminetheapoptosisofprimaryperitonealmacrophagesaftermelatoninand/orCpG-ODNtreat-ment.
AsshowninFig.
3A,B,1mMmelatonindidnotincreasetheapoptoticcellpercentageofmacrophagesirrespectiveofCpG-ODNstimulation,indicatingthatmelatoninhadnocytotoxiceffectsonmacrophagesandthesuppressionofTLR9-mediatedproinflammatorycytokinesproductioncouldnotbeattributabletoadirectcytotoxiceffectbymelatonin.
MelatoninmembranereceptorsMT1andMT2aredispensableforthesuppressiveeffectsofmelatonin.
MelatoninexertsavarietyofphysiologicalandpharmacologicaleffectsontheimmunesystemthroughactivationoftwomelatoninmembranereceptorsMT1andMT2,whichbelongtoGprotein-coupledreceptors(GPCRs)19.
WefoundthatperitonealmacrophagesexpressedmelatoninmembranereceptorsMT1andMT2,andmelatonintreatmentupregulatedmRNAexpressionsofMT1andMT2inmacrophages,whereasCpG-ODNstimulationdidnotaffectmRNAexpressionsofMT1andMT2(Fig.
4A,B).
ToinvestigatewhetherMT1andMT2wererequiredforthesuppressiveeffectsofmelatoninonTLR9-mediatedinflammatorycytokinesproduction,luzindole,anonselectiveantagonistofbothMT1andMT2,wasused.
AsshowninFig.
5A–D,routineconcentrationofluzindole(10μM)20,21didnotreversetheinhibitoryeffectsofmelatoninonCpG-ODN-inducedTNF-α,IL-6andIL-12p70production,andevenfurtherenhancedthesuppressionfunctionofmelatoninon3TLR9-mediatedTNF-αandIL-6production.
Theseresultsdemonstratedthatmelatoninmembranerecep-torsMT1andMT2weredispensableforthesuppressiveeffectsofmelatoninonTLR9-mediatedinflammatorycytokinesproductioninmacrophages.
MelatonininhibitsTLR9ligand-inducedERK1/2andAKTactivation.
TLR9agonistCpG-ODNinducesproinflammatorycytokinesproductionbyactivatingMyD88-dependentMAPK,PI-3K/AKTandNF-κBpathways1,22.
ToinvestigatethemechanismbywhichmelatoninsuppressedCpG-ODN-inducedinflammatorycytokinesproduction,wefirstdetecteditseffectsonTLR9andMyD88expression,andfoundthat1mMmel-atonintreatmentdidnotaffectmRNAexpressionofTLR9andMyD88inprimaryperitonealmacrophages,irrespectiveofCpG-ODNstimulation(Fig.
6A,B).
Wethenstudiedmelatonin'seffectsonCpG-ODN-inducedMAPK,AKTandNF-κBactivationinperitonealmacrophages.
AsshowninFig.
7A,2μg/mlCpG-ODNremark-ablyinducedERK1/2,p38,AKT,andNF-κBp65phosphorylationwhileitdidnotinduceJNKphosphorylation.
Interestingly,melatoninselectivelyattenuatedCpG-ODN-inducedERK1/2andAKTphosphorylation,buthadnoeffectsonp38andNF-κBp65phosphorylation(Fig.
7A–C).
TheseresultssuggestedthatmelatoninsuppressedCpG-ODN-inducedinflammatorycytokinesproductionprobablybyinhibitingERK1/2andAKTactivation.
MelatonininhibitsTLR9-mediatedproinflammatorycytokinesproductioninvivo.
ToexploretheroleofmelatonininregulatingTLR9signalinginmacrophagesinvivo,C57BL/6Jmicewereintraperitone-allyinjectedwithCpG-ODNinthepresenceorabsenceofmelatoninpretreatmentaftermobilizingperitonealmacrophagesthroughthioglycollate(TG)treatment,andthenthelevelsofTNF-αandIL-6inmouseserumweredetected.
AsshowninFig.
8A,B,CpG-ODNinducedhighlevelsofTNF-αandIL-6inserumandmelatoninpretreatmentsignificantlyinhibitedCpG-ODN-inducedTNF-αandIL-6productioninserum,indicatingthatmelatonininhibitedTLR9-mediatedproinflammatorycytokinesproductioninvivo.
DiscussionIncreasingevidencesshowthatmelatoninnotonlyexertsanimmunosuppressiveeffectonmacrophagesbutalsoexertsanimmunostimulatoryeffectonmacrophages,possiblydependingonthemannersofmelatoninadmin-istrationorthedoseofmelatonin13.
Forexample,ithasbeenreportedthatmelatoninstimulatedasignificantincreaseofIL-12productioninhumansynovialmacrophagesandTHP-1cells,however,asignificantreductionofIL-12productionwasobservedwhenmacrophageswerepreviouslyactivatedwithLPSandthentreatedwithmel-atonin23.
RegardingTLRssignaling,ithasbeendemonstratedthatmelatoninmainlyexertsasuppressiveeffectonTLRssignaling,includingTLR3andTLR4signalings8–10,13.
Inthepresentstudy,wedemonstratedforthefirstFigure1.
TheeffectsofmelatoninonmRNAexpressionsofcytokinesmediatedbyTLR9inperitonealmacrophages.
Peritonealmacrophageswerepre-treatedwith1mMMTorsolventcontrol(ethanol,E)for1hourandthenstimulatedwith2μg/mlCpG-ODNfor0,1.
5or3hours.
TNF-α(A),IL-6(B),IL-12p35(C)IL-12p40(D),IL-10(E)andIFN-β(F)mRNAexpressionsweredetectedbyreal-timePCR.
Dataareshownasmean±s.
d.
(n=3).
*P<0.
05,**P<0.
01(Student'st-test).
Similarresultswereobtainedinthreeindependentexperiments.
4Figure2.
MelatoninsuppressesTLR9-mediatedinflammatorycytokinesproductioninperitonealmacrophages.
Peritonealmacrophageswerepre-treatedwithvariousconcentrationsofMTorEfor1hourandthenstimulatedwith2μg/mlCpG-ODNfor24hours.
TNF-α(A),IL-6(B),IL-12p70(C)andIL-10(D)inthesupernatantswereassayedbyELISA.
Dataareshownasmean±s.
d.
(n=3).
*P<0.
05,**P<0.
01(ANOVA).
Similarresultswereobtainedinthreeindependentexperiments.
Figure3.
Melatonindoesnotinfluencetheviabilityofperitonealmacrophages.
(A)Peritonealmacrophageswerepre-treatedwith1mMMTorEfor1hourandthenstimulatedwith2μg/mlCpG-ODNfor24hours.
CellswerestainedwithannexinV/7AADandtheapoptosiswasassayedbyflowcytometry.
NumbersinplotsindicatethepercentageofannexinV7AADcells(livecells),annexinV+7AADcells(earlyapoptoticcells)andannexinV+7AAD+cells(lateapoptoticcells)inmacrophages.
(B)ThepercentageoflivecellsinAwereanalyzedstatisticallyandsummarized.
Similarresultswereobtainedinthreeindependentexperiments.
5timethatmelatoninsuppressedTLR9-mediatedinflammatoryresponsesinmacrophages,indicatingthatmela-toninwasanegativeregulatorofTLR9-mediatedinnateimmunityinmacrophages.
Althoughmelatoninupreg-ulatedmelatoninmembranereceptorsMT1andMT2mRNAexpressioninmacrophages,MT1andMT2weredispensableforthesuppressiveeffectsofmelatoninonTLR9-mediatedinflammatoryresponses.
Furthermore,Figure4.
MelatoninupregulatesmelatoninmembranereceptorsMT1andMT2expressionsinperitonealmacrophages.
Peritonealmacrophageswerepre-treatedwith1mMMTorEfor1hourandthenstimulatedwith2μg/mlCpG-ODNforindicatedtimes.
MelatoninmembranereceptorsMT1(A)andMT2(B)mRNAexpressionsweredetectedbyreal-timePCR.
Dataareshownasmean±s.
d.
(n=3).
*P<0.
05,**P<0.
01(Student'st-test).
Similarresultswereobtainedinthreeindependentexperiments.
Figure5.
MT1andMT2aredispensableforthesuppressiveeffectsofmelatoninonTLR9signaling.
Peritonealmacrophageswereincubatedwith10μMluzindole(anonselectiveantagonistofmelatoninmembranereceptorsMT1/MT2)for1hourandthentreatedwith1mMMTorEfor1hour.
Afterthat,cellswerestimulatedwith2μg/mlCpG-ODNfor24hours.
TNF-α(A),IL-6(B),IL-12p70(C)andIL-10(D)inthesupernatantswereassayedbyELISA.
Dataareshownasmean±s.
d.
(n=3).
"ns"indicatesnosignificance,*P<0.
05(ANOVA).
Similarresultswereobtainedinthreeindependentexperiments.
6melatonindidnotaffectTLR9andMyD88mRNAexpression,butpossiblyattenuatedERK1/2andAKTactiva-tiontosuppressTLR9signaling.
AfterstimulationwithTLRligandssuchasLPS,polyI:CorCpG-ODN,activatedmacrophagesproducevar-iouscytokinesincludingTNF-α,IL-6,IL-12,IL-1β,IL-10,andtype1interferonincludingIFN-αandIFN-β.
TLR9ligandCpG-ODNinducesmacrophagesanddendriticcellstosecretTNF-α,IL-6,IL-12,IL-10andIFN-α/β(mainlyproducedbyplasmacytoiddendriticcells)24,25.
ThepresentstudydemonstratedthatmelatoninsuppressedthemRNAandproteinexpressionofTNF-α,IL-6andIL-12p70inducedbyCpG-ODN,indicatingthatmelatonincouldsuppressTLR9-mediatedproinflammatorycytokinesproductioninmacrophages.
Similarly,previousstudiesshowedthatmelatoninsuppressedLPS-inducedTNF-α,IL-6,IL-1β,andIL-8expressioninRAW264.
7cells9,10.
IL-10inducedbyCpG-ODN,ananti-inflammatorycytokine,couldsuppressIL-12p70pro-ductioninmacrophages26.
OurpresentstudydemonstratedthatmelatonindidnotsuppressorpromoteIL-10proteinproductioninducedbyCpG-ODN,whichisdifferentfromanotherstudyshowingthatmelatoninsup-pressedLPS-inducedIL-10expression10.
ThisindicatesthatIL-10isnotinvolvedintheinhibitoryeffectsofmel-atoninonTLR9-mediatedinflammatoryresponsesinmacrophages.
Themolecularmechanismsofactionofmelatoninmainlyincludethefollowingfourmodels:(i)viaitsinter-actionwithmembranereceptorssuchasMT1andMT2(atlowconcentration),(ii)bybindingtoitsnuclearreceptorssuchasRORαandRZR(atlowconcentration),(iii)viaaninteractionwithcytoplasmicproteinssuchascalmodulinandhydroquinone(athighconcentration),and(iv)viadirectly,receptor-independentactionssuchasscavengingROS/RNS13,15,27.
Ithasbeenshownthattheeffectsofmelatoninonimmunesystemaremedi-atedthroughmelatoninreceptor-dependentor-independentmechanisms.
Inmultiplesclerosis(MS),melatoninblocksTh17differentiationandbooststhegenerationofIL-10-secretingTregcellsbyanMT1-dependentmech-anism28,29.
MelatonininhibitstheinfectivityofLeishmaniabymurineperitonealmacrophagesandenhancesthephagocytosisofzymosanbyhumancolostrummononuclearcells,bothofwhichareattenuatedbyluzindole(anantagonistofMT1andMT2),indicatingtheinvolvementofmelatoninmembranereceptorsintheseeffects30,31.
ThepresentstudyshowedthatluzindoledidnotreversetheinhibitoryeffectsofmelatoninonTLR9-mediatedinflammatoryresponsesinmacrophages,somelatoninexertsitsinhibitoryeffectsonTLR9signalingthroughmelatoninmembranereceptor-independentway.
Giventhatmelatoninathighconcentrationalsoexertsitseffectsthroughreceptor-independentmannersinadditiontoitsinteractionwitheithermelatoninmembraneornuclearreceptors13,15,27,inthepresentstudy,high-concentrationmelatonin(reachingthelevelofmillimole)probablydirectlyregulatedtheexpressionoractivationofTLR9signalmolecules,similartoitsmechanismsinregulatingTLR3orTLR4signalinginmac-rophages8–10.
MelatonindecreasedTLR-3-mediatedTNF-αandiNOSmRNAexpressioninrespiratorysyncy-tialvirus-infectedRAW264.
7cellsviainhibitionofNF-κBactivationwithoutinfluencingTLR3andMyD88mRNAexpression8.
InLPS-stimulatedRAW264.
7cells,melatoninattenuatedtheexpressionofMyD88andtheactivationofNF-κB,andalsoinhibitedTLR4-mediatedAKTphosphorylationandattenuatedtheelevationofIRF3,whichwasinvolvedinTLR4-mediatedTRIF-dependentsignalingpathway10.
Inanotherstudy,melatonininhibitedLPSinducedproductionofnitricoxideandIL-6inRAW264.
7cellsbysuppressingNF-κBandSTAT1activation9.
Inthepresentstudy,melatonininhibitedCpG-ODN-inducedERK1/2andAKTactivationwithoutaffectingNF-κBandp38activationinmacrophages.
Furthermore,melatonindidnotaffectTLR9andMyD88mRNAexpression.
Hence,melatoninmightsuppressTLR9-mediatedinnateimmuneresponsesthroughinhib-itingERK1/2andAKTactivation.
Thisstudy,togetherwithotherpreviousstudies,indicatethatmelatoninreg-ulatesTLRsignalinginmacrophagesthroughdifferentmechanisms,dependentondifferentTLRsubtypesandtheirligands8–10.
ItremainstobefurtherinvestigatedhowmelatoninregulatesTLR9-mediatedERK1/2andAKTactivationinmacrophages.
Inourpreviousstudy,MEKK2knockdown(specificallyinhibitingERK1/2activation)significantlysuppressedCpG-ODN-inducedTNF-αandIL-6productioninmacrophages4.
Inaddition,otherstudiesalsoshowedthatwortmannin,aPI-3Kinhibitor,inhibitedtheproductionofIL-12inducedbyCpGDNAFigure6.
TheeffectsofmelatoninonmRNAexpressionsofTLR9andMyD88inperitonealmacrophages.
Peritonealmacrophageswerepre-treatedwith1mMMTorEfor1hourandthenstimulatedwith2μg/mlCpG-ODNforindicatedtimes.
TLR9(A)andMyD88(B)mRNAexpressionsweredetectedbyreal-timePCR.
Dataareshownasmean±s.
d.
(n=3).
Similarresultswereobtainedinthreeindependentexperiments.
7inbonemarrowderiveddendriticcells32,andanotherPI-3Kinhibitor,LY294002,alsoreducedTNF-αproduc-tioninducedbyCpGDNAinmacrophages33.
ThesestudiesfurthersupportthatERK1/2andAKTactivationisimportantforTLR9-mediatedinflammatorycytokinesproductioninmacrophages.
ThereisonlyonepublishedresearchreportingontherelationshipbetweenmelatoninandTLR9signaling.
ThisstudyshowedthatJNK-enhancedTLR9signalpathwaymediatedallergicairwayinflammationthroughsup-pressingmelatoninbiosynthesis34.
However,thereisnoresearchaddressingtheregulatoryfunctionofmelatoninonTLR9signaling.
OurpresentstudyshowedforthefirsttimethatmelatoninregulatedTLR9-mediatedinnateimmuneresponsesandTLR9signalingpathways.
TLR9-mediatedinnateimmunityandinflammationplaysimportantrolesininfectiousdiseases,autoimmunediseasesandcancer25,35.
Previousstudiesshowedthatpolymicrobialsepsisinmouseexperimentalperitonitismodelinducedbycaecalligationandpuncture(CLP)wasTLR9-dependent36,37.
IthasbeendemonstratedthatmelatoninprotectsmiceagainstsepticshockorsepticorgansinjuryinCLPsepsismodelsthroughdifferentmechanisms:theseincludeantioxidativeeffects,anti-nitrosativeeffects,protectiveeffectsagainstmitochondrialdysfunctionandanti-inflammatoryeffects18,38–43.
Hence,itispossiblethatmelatoninalsoprovidesprotectionsFigure7.
MelatonininhibitsTLR9-mediatedERK1/2andAKTactivationinperitonealmacrophages.
Peritonealmacrophageswerepre-treatedwith1mMMTorEfor1hourandthenstimulatedwith2μg/mlCpG-ODNforindicatedtimes.
PhosphorylatedandtotalERK1/2,JNK,p38,AKTandNF-κBp65andβ-actinweredetectedbywesternblot.
Representativeimagesareshown(A)andrelativeexpressionlevelsofphosphorylatedERK1/2andAKTinvariousgroupsareshownbythepercentageoftheirexpressionlevelsin"E+CpG0min"group(B,C).
*P<0.
05,**P<0.
01(ANOVA,MTgroupsinvarioustimesarecomparedtoEgroupsinthesametime).
Dataarerepresentativeofthreeindependentexperimentswithsimilarresults.
8againstsepsisthroughsuppressingTLR9-mediatedinflammation,whichrequirestobefurtherinvestigated.
Furthermore,ourpresentstudydemonstratedthatinvivoadministrationofmelatoninexertednotablesuppres-siveeffectsonTLR9–mediatedinflammatorycytokinesproductioninmice,furtherraisingthispossibility.
Takentogether,ourstudydemonstratedthatmelatoninsuppressedTLR9-triggeredproinflammatoryTNF-α,IL-6andIL-12p70productioninmacrophagesnotthroughmelatoninmembranereceptors-dependentmannersbutprobablythroughinhibitingTLR9-mediatedERK1/2andAKTactivation.
Theseresultsprovedthatmela-toninwasanegativeregulatorofTLR9-mediatedinnateimmunityinmacrophages.
MaterialsandMethodsMiceandreagents.
C57BL/6Jmice(female,6–8weeksold)wereobtainedfromJointVenturesSipperBKExperimentalAnimalCompany(Shanghai,China).
AllanimalexperimentswereperformedinaccordancewiththeNationalInstitutesofHealthGuidefortheCareandUseofLaboratoryAnimals,withtheapprovaloftheScientificInvestigationBoardofSecondMilitaryMedicalUniversity(Shanghai,China).
Melatonin(M5250),7-aminoactinomycin(7AAD,A9400)andluzindole(L2407)werefromSigma-Aldrich(StLouis,MO).
Phosphorothioate-modifiedCpG-ODN1668(5′-TCCATGACGTTCCTGATGCT-3′)wassynthesizedbySangonBiotechCompany(Shanghai,China).
EndotoxinintheseODNswasremovedusingEndotoxinRemovalSolution(Sigma-Aldrich)asdescribedpreviously44,andtheendotoxinlevelwas<0.
015U/mgCpG-ODN.
AntibodiesspecificforERK1/2(9107),phosphorylatedERK1/2(9101),JNK(9252),phosphorylatedJNK(9255),p38(9212),phosphorylatedp38(9211),AKT(9272),phosphorylatedAKT(4060),p65(3034),andphosphoryl-atedp65(3033)werefromCellSignalingTechnology.
AnnexinVapoptosisdetectionkitAPCwasfromeBio-science(ThermoFisherscientific).
Antibodyspecificforβ-actin(sc-1616)andHRP-coupledsecondaryantibodywerefromSantaCruz.
Cellcultureandtreatments.
Thioglycollate-elicitedmouseperitonealmacrophageswereisolatedandcul-turedaspreviouslydescribed4.
C57BL/6Jmice(6–8weeksold)wereintraperitoneallyinjectedwith1mlof3%sodiumthioglycollate(Merck)solutionpermouse.
After3days,cellsintheabdominalcavitywerecollectedbylavagingwith10–15mlFBS-freeDMEMandcentrifugated.
CellswereculturedandmaintainedinDMEMwith10%(vol/vol)FBSfor24hours.
Afterremovingthefloatingcells,adheredperitonealmacrophagesweresubjectedforfurtherexperiments.
Thecellswerepre-treatedwithvariousconcentrationsofmelatonin(stockconcentra-tion200mM,dissolvedinethanol)for1hourandthenstimulatedwith2μg/mlCpG-ODNforindicatedtimes(SupplementaryFig.
1).
Insomeexperiments,thecellswereincubatedwith10μMluzindolefor1hourbeforepretreatmentwithmelatonin(SupplementaryFig.
1C).
Detectionofcytokineproduction.
TheconcentrationsofTNF-α,IL-6,IL-12p70andIL-10inculturesupernatantsorserumweremeasuredwithELISAkitsfromR&Dsystems(Minneapolis,MN)accordingtothemanufacturer'sinstructions.
Detectionofcellapoptosis.
Aftertreatmentwithmelatoninand/orCpG-ODN(SupplementaryFig.
1B),peritonealmacrophageswereharvestedandstainedwithAnnexinV-APCand7AADaccordingtothemanufac-turer'sinstructions.
Thentheapoptosisofmacrophageswasassayedbyflowcytometry(LSRII,BDbiosciences)andthedatawereanalyzedwithFlowJoVersion5.
7.
2software(TreeStar).
Quantitativereversetranscription-PCR.
TotalRNAwasextractedwithRNAextractionkit(RNAfast200,FastagenBiotechCo.
,China)andreversed-transcriptedwithreversetranscriptaseM-MLX(Takara)followingthemanufacturer'sinstructions.
Real-timePCRwasperformedwithaSYBRRT-PCRkit(Takara)onLightCyclersystemFigure8.
MelatonininhibitsTLR9-mediatedproinflammatorycytokinesproductioninvivo.
C57BL6/Jmicewereintraperitoneallyinjectedwiththioglycollate(TG)toelicitemacrophagesonday0andthenreceived5mg/kgmelatonin(MT)orethanol(E)onday3andday4.
Onday4,micewerefurtherstimulatedbyintraperitonealinjectionof50μgCpG-ODNtwohoursaftermelatoninpretreatment.
TNF-α(A)andIL-6(B)inserumofmiceinvariousgroups(n=5)wereassayedbyELISA2hoursafterCpG-ODNinjection.
Errorbarsrepresents.
d.
**P<0.
01(ANOVA).
9(Roche).
Datawerenormalizedbythelevelofβ–actinexpressionineachindividualsample.
2Ctmethodwasusedtocalculaterelativeexpressionchanges.
ThesequencesoftheprimersusedwerefromPrimerBankasfollows:β–actinsense,5′-GGCTGTATTCCCCTCCATCG-3′andantisense,5′-CCAGTTGGTAACAATGCCATGT-3′;MT1sense,5′-TACACTGTCAAGTCAGCGCAT-3′andantisense,5′AGCAGTAACGGTTCATAGCGA-3′;MT2sense,5′-GAACAGCTCAATCCCTAACTGC-3′andantisense,5′-ACGACTACTGTAGATAGCATGGG-3′;TNF-αsense,5′-CCCTCACACTCAGATCATCTTCT-3′andantisense,5′-GCTACGACGTGGGCTACAG-3′;IL-6sense,5′-TAGTCCTTCCTACCCCAATTTCC-3′andantisense,5′-TTGGTCCTTAGCCACTCCTTC-3′;IL-12p35sense,5′-CTGTGCCTTGGTAGCATCTATG-3′andantisense,5′-GCAGAGTCTCGCCATTATGATTC-3′;IL-12p40sense,5′-TGGTTTGCCATCGTTTTGCTG-3′andantisense,5′-ACAGGTGAGGTTCACTGTTTCT-3′;IL-10sense,5′-GCTCTTACTGACTGGCATGAG-3′andantisense,5′-CGCAGCTCTAGGAGCATGTG-3′;IFN-βsense,5′-CAGCTCCAAGAAAGGACGAAC-3′andantisense,5′-GGCAGTGTAACTCTTCTGCAT-3′;TLR9sense,5′-ATGGTTCTCCGTCGAAGGACT-3′andantisense,5′-GAGGCTTCAGCTCACAGGG-3′;MyD88sense,5′-TCATGTTCTCCATACCCTTGGT-3′andantisense,5′-AAACTGCGAGTGGGGTCAG-3′.
Westernblot.
Afterstimulatedwith2μg/mlCpG-ODNforindicatedtimes(SupplementaryFig.
1D),pri-maryperitonealmacrophageswerewashedtwicewithice-coldPBSandlysedwithM-PERProteinExtractionReagent(Pierce)supplementedwithproteaseinhibitorcocktail,andthenproteinconcentrationsoftheextractsweremeasuredwithBCAassayaccordingtothemanufacturer'sinstructions(Pierce).
EqualamountsofextractswereloadedforSDS–PAGE,transferredontonitrocellulosemembranesandthenblottedwithindicatedantibod-ies,asdescribedpreviously4.
Finally,signalintensitywasdeterminedusingtheTanon5200SChemiluminescentImagingSystem(Tanon).
Imageshadbeencroppedforpresentation.
Full-sizeimageswerepresentedinSupplementaryFig.
2.
InductionofproinflammatorycytokinesbyCpG-ODNinvivo.
C57BL/6Jmice(6–8weeksold)wereintraperitoneallyinjectedwith1mlof3%sodiumthioglycollate(Merck)solutiononday0.
Onday3andday4,micewereintraperitonealinjectedwith5mg/kgmelatonin.
Onday4,micewerestimulatedbyintraperito-nealinjectionof50μgCpG-ODN2hoursaftermelatonininjection(SupplementaryFig.
1E).
TwohoursafterCpG-ODNinjection,theserumsampleswerecollectedandusedforELSIAassaysofTNF-αandIL-6.
Statisticalanalysis.
Alldatawerepresentedasmeans±standarddeviation(S.
D.
).
Thestatisticalsignifi-cancewascalculatedbyone-wayANOVAtestortwo-tailedStudent'sttestusingPrismsoftware5(GraphPadSoftware,Inc.
,SanDiego,CA).
Pvaluelessthan0.
05wasconsideredstatisticallysignificant.
DataAvailabilityAlldatageneratedoranalyzedduringthisstudyareincludedinthismanuscript.
References1.
Kawai,T.
&Akira,S.
Theroleofpattern-recognitionreceptorsininnateimmunity:updateonToll-likereceptors.
Nat.
Immunol11,373–384(2010).
2.
Cao,X.
Self-regulationandcross-regulationofpattern-recognitionreceptorsignallinginhealthanddisease.
Nat.
Rev.
Immunol16,35–50(2016).
3.
Pandey,S.
,Kawai,T.
&Akira,S.
MicrobialsensingbyToll-likereceptorsandintracellularnucleicacidsensors.
ColdSpringHarb.
Perspect.
Biol7,a016246(2014).
4.
Wen,M.
etal.
Stk38proteinkinasepreferentiallyinhibitsTLR9-activatedinflammatoryresponsesbypromotingMEKK2ubiquitinationinmacrophages.
Nat.
Commun6,7167(2015).
5.
Xia,M.
etal.
HistonemethyltransferaseAsh1lsuppressesinterleukin-6productionandinflammatoryautoimmunediseasesbyinducingtheubiquitin-editingenzymeA20.
Immunity39,470–481,https://doi.
org/10.
1016/j.
immuni.
2013.
08.
016(2013).
6.
Xu,S.
etal.
ConstitutiveMHCclassImoleculesnegativelyregulateTLR-triggeredinflammatoryresponsesviatheFps-SHP-2pathway.
NatImmunol13,551–559,https://doi.
org/10.
1038/ni.
2283(2012).
7.
Zhang,Q.
etal.
Tet2isrequiredtoresolveinflammationbyrecruitingHdac2tospecificallyrepressIL-6.
Nature525,389–393,https://doi.
org/10.
1038/nature15252(2015).
8.
Huang,S.
H.
,Cao,X.
J.
&Wei,W.
MelatonindecreasesTLR3-mediatedinflammatoryfactorexpressionviainhibitionofNF-kappaBactivationinrespiratorysyncytialvirus-infectedRAW264.
7macrophages.
J.
PinealRes45,93–100(2008).
9.
Choi,E.
Y.
etal.
MelatonininhibitsPrevotellaintermedialipopolysaccharide-inducedproductionofnitricoxideandinterleukin-6inmurinemacrophagesbysuppressingNF-kappaBandSTAT1activity.
J.
PinealRes50,197–206(2011).
10.
Xia,M.
Z.
etal.
MelatoninmodulatesTLR4-mediatedinflammatorygenesthroughMyD88-andTRIF-dependentsignalingpathwaysinlipopolysaccharide-stimulatedRAW264.
7cells.
J.
PinealRes53,325–334(2012).
11.
Qin,X.
etal.
MicheliolideinhibitsLPS-inducedinflammatoryresponseandprotectsmicefromLPSchallenge.
SciRep6,23240,https://doi.
org/10.
1038/srep23240(2016).
12.
Arranz,A.
etal.
Vasoactiveintestinalpeptidesuppressestoll-likereceptor4expressioninmacrophagesviaAkt1reducingtheirresponsivenesstolipopolysaccharide.
MolImmunol45,2970–2980,https://doi.
org/10.
1016/j.
molimm.
2008.
01.
023(2008).
13.
Calvo,J.
R.
,Gonzalez-Yanes,C.
&Maldonado,M.
D.
Theroleofmelatonininthecellsoftheinnateimmunity:areview.
J.
PinealRes55,103–120(2013).
14.
cuna-Castroviejo,D.
etal.
Extrapinealmelatonin:sources,regulation,andpotentialfunctions.
CellMol.
LifeSci71,2997–3025(2014).
15.
Radogna,F.
,Diederich,M.
&Ghibelli,L.
Melatonin:apleiotropicmoleculeregulatinginflammation.
Biochem.
Pharmacol80,1844–1852(2010).
16.
Mauriz,J.
L.
,Collado,P.
S.
,Veneroso,C.
,Reiter,R.
J.
&Gonzalez-Gallego,J.
Areviewofthemolecularaspectsofmelatonin'santi-inflammatoryactions:recentinsightsandnewperspectives.
J.
PinealRes54,1–14(2013).
17.
Carrillo-Vico,A.
,Lardone,P.
J.
,varez-Sanchez,N.
,Rodriguez-Rodriguez,A.
&Guerrero,J.
M.
Melatonin:bufferingtheimmunesystem.
Int.
J.
Mol.
Sci14,8638–8683(2013).
18.
Hu,W.
etal.
Utilizingmelatonintocombatbacterialinfectionsandsepticinjury.
Br.
J.
Pharmacol174,754–768(2017).
19.
Jockers,R.
etal.
Updateonmelatoninreceptors:IUPHARReview20.
Br.
J.
Pharmacol173,2702–2725(2016).
20.
Yang,Y.
etal.
Melatoninreversesflowshearstress-inducedinjuryinbonemarrowmesenchymalstemcellsviaactivationofAMP-activatedproteinkinasesignaling.
JPinealRes60,228–241,https://doi.
org/10.
1111/jpi.
12306(2016).
1021.
Zhou,L.
etal.
MelatoninreversesH2O2-inducedprematuresenescenceinmesenchymalstemcellsviatheSIRT1-dependentpathway.
JPinealRes59,190–205,https://doi.
org/10.
1111/jpi.
12250(2015).
22.
Ishii,M.
etal.
CCchemokinereceptor4modulatesToll-likereceptor9-mediatedinnateimmunityandsignaling.
Eur.
J.
Immunol38,2290–2302(2008).
23.
Cutolo,M.
etal.
Melatonininfluencesinterleukin-12andnitricoxideproductionbyprimaryculturesofrheumatoidsynovialmacrophagesandTHP-1cells.
Ann.
N.
Y.
Acad.
Sci876,246–254(1999).
24.
Krieg,A.
M.
TherapeuticpotentialofToll-likereceptor9activation.
Nat.
Rev.
DrugDiscov5,471–484(2006).
25.
Hennessy,E.
J.
,Parker,A.
E.
&O'Neill,L.
A.
TargetingToll-likereceptors:emergingtherapeuticsNat.
Rev.
DrugDiscov9,293–307(2010).
26.
Boonstra,A.
etal.
Macrophagesandmyeloiddendriticcells,butnotplasmacytoiddendriticcells,produceIL-10inresponsetoMyD88-andTRIF-dependentTLRsignals,andTLR-independentsignals.
J.
Immunol177,7551–7558(2006).
27.
Reiter,R.
J.
etal.
Melatoninasanantioxidant:underpromisesbutoverdelivers.
J.
PinealRes61,253–278(2016).
28.
Farez,M.
F.
etal.
MelatoninContributestotheSeasonalityofMultipleSclerosisRelapses.
Cell162,1338–1352(2015).
29.
Ren,W.
etal.
MelatoninsignalinginTcells:Functionsandapplications.
J.
PinealRes62(2017).
30.
Pires-Lapa,M.
A.
,Tamura,E.
K.
,Salustiano,E.
M.
&Markus,R.
P.
Melatoninsynthesisinhumancolostrummononuclearcellsenhancesdectin-1-mediatedphagocytosisbymononuclearcells.
J.
PinealRes55,240–246(2013).
31.
Laranjeira-Silva,M.
F.
,Zampieri,R.
A.
,Muxel,S.
M.
,Floeter-Winter,L.
M.
&Markus,R.
P.
MelatoninattenuatesLeishmania(L.
)amazonensisinfectionbymodulatingargininemetabolism.
J.
PinealRes59,478–487(2015).
32.
Ishii,K.
J.
etal.
Potentialroleofphosphatidylinositol3kinase,ratherthanDNA-dependentproteinkinase,inCpGDNA-inducedimmuneactivation.
J.
Exp.
Med196,269–274(2002).
33.
Sester,D.
P.
etal.
CpGDNAactivatessurvivalinmurinemacrophagesthroughTLR9andthephosphatidylinositol3-kinase-Aktpathway.
J.
Immunol177,4473–4480(2006).
34.
Wu,H.
M.
etal.
JNK-TLR9signalpathwaymediatesallergicairwayinflammationthroughsuppressingmelatoninbiosynthesis.
J.
PinealRes60,415–423(2016).
35.
Savva,A.
&Roger,T.
Targetingtoll-likereceptors:promisingtherapeuticstrategiesforthemanagementofsepsis-associatedpathologyandinfectiousdiseases.
FrontImmunol4,387(2013).
36.
Yasuda,H.
etal.
ChloroquineandinhibitionofToll-likereceptor9protectfromsepsis-inducedacutekidneyinjury.
Am.
J.
PhysiolRenalPhysiol294,F1050–F1058(2008).
37.
Plitas,G.
,Burt,B.
M.
,Nguyen,H.
M.
,Bamboat,Z.
M.
&DeMatteo,R.
P.
Toll-likereceptor9inhibitionreducesmortalityinpolymicrobialsepsis.
J.
Exp.
Med205,1277–1283(2008).
38.
Srinivasan,V.
,Mohamed,M.
&Kato,H.
Melatonininbacterialandviralinfectionswithfocusonsepsis:areview.
RecentPatEndocr.
MetabImmune.
DrugDiscov6,30–39(2012).
39.
An,R.
etal.
Melatoninattenuatessepsis-inducedcardiacdysfunctionviaaPI3K/Akt-dependentmechanism.
BasicRes.
Cardiol111,8(2016).
40.
Zhao,L.
etal.
Melatoninalleviatesbraininjuryinmicesubjectedtocecalligationandpunctureviaattenuatinginflammation,apoptosis,andoxidativestress:theroleofSIRT1signaling.
J.
PinealRes59,230–239(2015).
41.
Ersoy,O.
F.
etal.
Effectsofmelatoninoncytokinereleaseandhealingofcolonicanastomosesinanexperimentalsepsismodel.
Ulus.
Travma.
Acil.
Cerrahi.
Derg22,315–321(2016).
42.
Wu,J.
Y.
etal.
Therapeuticeffectsofmelatoninonperitonitis-inducedsepticshockwithmultipleorgandysfunctionsyndromeinrats.
J.
PinealRes45,106–116(2008).
43.
Ortiz,F.
etal.
Thebeneficialeffectsofmelatoninagainstheartmitochondrialimpairmentduringsepsis:inhibitionofiNOSandpreservationofnNOS.
J.
PinealRes56,71–81(2014).
44.
Wu,Y.
etal.
CMRF-35-likemolecule3preferentiallypromotesTLR9-triggeredproinflammatorycytokineproductioninmacrophagesbyenhancingTNFreceptor-associatedfactor6ubiquitination.
J.
Immunol187,4881–4889(2011).
AcknowledgementsWethankDrHongmeiXuforcriticalrevisionofthismanuscript.
ThisworkwassupportedbythegrantsfromtheNationalNaturalScienceFoundationofChina[No.
31100630(XuX)andNo.
81502466(ZhangJ)]andNaturalScienceFoundationofShanghai[No.
18ZR1449400(XuX)].
AuthorContributionsX.
X.
contributedtothestudydesign;X.
X.
andZ.
J.
obtainedfunding;X.
X.
,W.
G.
,A.
L.
,S.
J.
,Z.
J.
andC.
Y.
X.
performedtheexperimentsandanalyzedthedata;X.
X.
wrotethemanuscript.
Alltheauthorscriticallyrevisedthearticleandhaveconsentedthefinalversionofthemanuscript.
AdditionalInformationSupplementaryinformationaccompaniesthispaperathttps://doi.
org/10.
1038/s41598-018-34011-8.
CompetingInterests:Theauthorsdeclarenocompetinginterests.
Publisher'snote:SpringerNatureremainsneutralwithregardtojurisdictionalclaimsinpublishedmapsandinstitutionalaffiliations.
OpenAccessThisarticleislicensedunderaCreativeCommonsAttribution4.
0InternationalLicense,whichpermitsuse,sharing,adaptation,distributionandreproductioninanymediumorformat,aslongasyougiveappropriatecredittotheoriginalauthor(s)andthesource,providealinktotheCre-ativeCommonslicense,andindicateifchangesweremade.
Theimagesorotherthirdpartymaterialinthisarticleareincludedinthearticle'sCreativeCommonslicense,unlessindicatedotherwiseinacreditlinetothematerial.
Ifmaterialisnotincludedinthearticle'sCreativeCommonslicenseandyourintendeduseisnotper-mittedbystatutoryregulationorexceedsthepermitteduse,youwillneedtoobtainpermissiondirectlyfromthecopyrightholder.
Toviewacopyofthislicense,visithttp://creativecommons.
org/licenses/by/4.
0/.
TheAuthor(s)2018
- immunostimulatoryvivox7和oppor9哪个好相关文档
- 修复vivox7和oppor9哪个好
- 答案vivox7和oppor9哪个好
- 公开vivox7和oppor9哪个好
- 扬尘vivox7和oppor9哪个好
- 贵州省住房和城乡建设厅关于征求《贵州省城镇园林绿化管护规范》
- 甘肃省vivox7和oppor9哪个好
RackNerd提供四款高配美国服务器促销活动低至月$189
RackNerd 商家给的感觉就是一直蹭节日热点,然后时不时通过修改配置结构不断的提供低价年付的VPS主机,不过他们家还是在做事的,这么两年多的发展,居然已经有新增至十几个数据中心,而且产品线发展也是比较丰富。比如也有独立服务器业务,不过在他们轮番的低价年付VPS主机活动下,他们的服务器估摸着销路不是太好的。这里,今天有看到RackNerd商家的独立服务器业务有促销。这次提供美国多个机房的高配独立...
2022年腾讯云新春采购季代金券提前领 领取满减优惠券和域名优惠
2022年春节假期陆续结束,根据惯例在春节之后各大云服务商会继续开始一年的促销活动。今年二月中旬会开启新春采购季的活动,我们已经看到腾讯云商家在春节期间已经有预告活动。当时已经看到有抢先优惠促销活动,目前我们企业和个人可以领取腾讯云代金券满减活动,以及企业用户可以领取域名优惠低至.COM域名1元。 直达链接 - 腾讯云新春采购活动抢先看活动时间:2022年1月20日至2022年2月15日我们可以在...
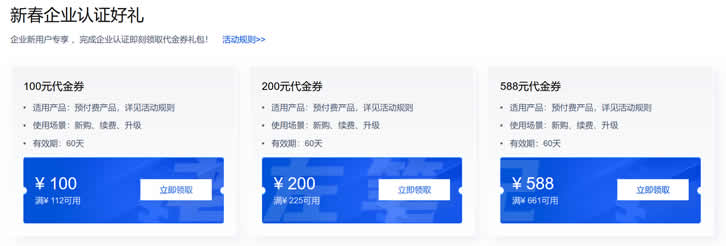
简单测评v5.net的美国cn2云服务器:电信双程cn2+联通AS9929+移动直连
v5.net一直做独立服务器这块儿的,自从推出云服务器(VPS)以来站长一直还没有关注过,在网友的提醒下弄了个6G内存、2核、100G SSD的美国云服务器来写测评,主机测评给大家趟雷,让你知道v5.net的美国云服务器效果怎么样。本次测评数据仅供参考,有兴趣的还是亲自测试吧! 官方网站:https://v5.net/cloud.html 从显示来看CPU是e5-2660(2.2GHz主频),...
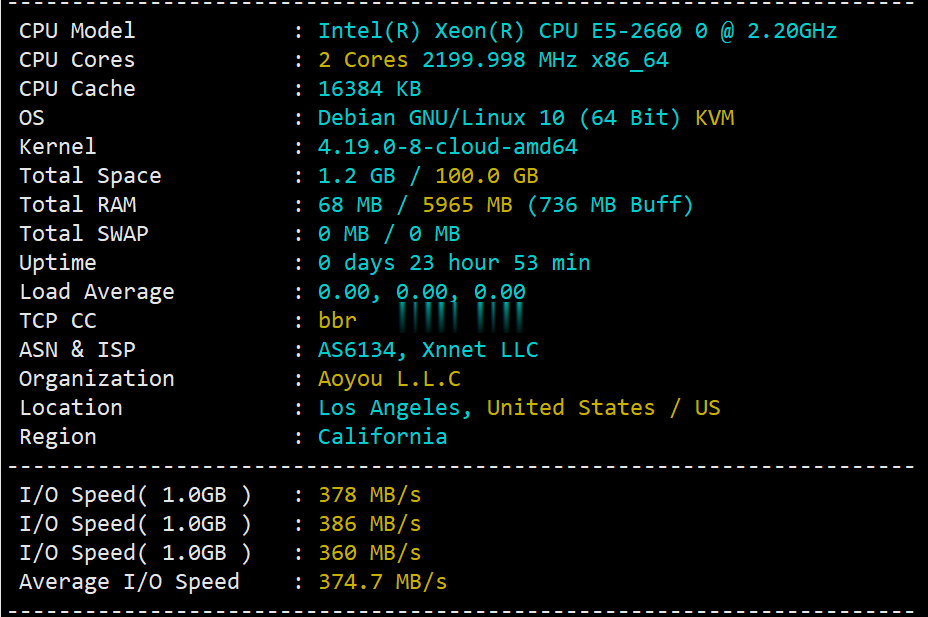
vivox7和oppor9哪个好为你推荐
-
扫描仪morphvox打印机morphvoxfollowcss重要产品信息指南支持ipadcolumnios5win7如何关闭445端口如何关闭445端口,禁用smb协议css选择器CSS的常见选择器有哪几种迅雷下载速度迅雷下载速度真是太慢了win7还原系统win7如何一键还原电脑系统怎么操作