萌发郭杰非你莫属
郭杰非你莫属 时间:2021-05-03 阅读:()
生物资源2020,42(1):43~48BioticResources兰科杓兰属(Cypripedium)植物种子非共生萌发研究进展徐可1,2,王涛1,张毓1*(1.
北京市植物园北京市花卉园艺工程技术研究中心城乡生态环境北京实验室,北京100093;2.
天津师范大学地理与环境科学学院,天津300387)摘要:兰科杓兰属(Cypripedium)植物主要分布于东亚、北美等温带地区和亚热带山地.
杓兰不仅具有极高的观赏价值,而且其经济价值和科研价值也越来越受到人们的重视.
近年来对杓兰属植物人工繁殖的相关研究不断深入,主要集中于种子的非共生萌发等方面.
本文对濒危兰科杓兰属植物进行了简要介绍,并就其种子非共生萌发研究从种子成熟度、预处理和有机添加物的作用、培养基的配置等方面进行综述,为目标杓兰种类的非共生萌发试验方案的制定奠定基础,将有助于温带/高山兰科植物保育研究的发展.
关键词:杓兰属;非共生萌发;离体培养;研究进展中图分类号:Q943文献标识码:A文章编号:20963491(2020)01004306AdvancesinthestudyofasymbioticgerminationofCypripediumseedsXUKe1,2,WANGTao1,ZHANGYu1*(1.
BeijingFloricultureEngineeringTechnologyResearchCentre,BeijingLaboratoryofUrbanandRuralEcologicalEnvironment,BeijingBotanicalGarden,Beijing100093,China;2.
SchoolofGeographicandEnvironmentalSciences,TianjinNormalUniversity,Tianjin300387,China)Abstract:CypripediumplantsaremainlydistributedintemperateAsiaandNorthAmerica.
Theynotonlyhaveveryhighornamentalvalue,butalsotheireconomicvalueandscientificresearchvaluearemoreandmoreconcernedbypeople.
Inrecentyears,researchesonpropagationofCypripediumhavebeendeepened,mainlyfocusonasymbioticgerminationoftheseeds.
ThispaperbrieflyintroducestheendangeredorchidsofCypripedium,andsummarizestheresearchprogressofseedasymbioticgerminationfromaspectsofseedmaturity,pretreatment,theroleoforganicadditives,andtheconfigurationofmedia.
Thestudylaysthefoundationfortheasymbioticgerminationexperimentprogramoftheorchidspecies,andpromotestheresearchanddevelopmentoforchidplantconservation.
Keywords:Cypripedium;asymbioticgermination;invitroculture;researchprogress1杓兰属植物简介杓兰属(Cypripedium)植物属兰科(Orchidaceae)植物大家族,为多年生宿根花卉,因其花唇瓣特化成兜状、杓状或拖鞋状而得名,西方人称为拖鞋兰,即"lady'sslipper",台湾称作"仙履兰".
杓兰独特的外形,高观赏价值以及在植物保育中的旗舰地位,被称为"植物中的大熊猫"[1].
杓兰属植物全世界约有50种,北温带地区主要分布于温带的亚洲和北美洲,南至喜马拉雅地区和中美洲[2,3].
我国是杓兰属植物的世界分布中心,共有36种,其中25种为我国特有种,广泛分布于东北至西南的山地和台湾DOI:10.
14188/j.
ajsh.
2020.
01.
006收稿日期:20191009修回日期:20191120接受日期:20191203作者简介:徐可(1995-),男,硕士,主要从事园林植物研究.
E-mail:xuke19951010@126.
com通讯联系人:张毓(1971-),女,博士,教授级高级工程师,主要从事珍稀濒危植物保护研究.
E-mail:bjyzhang@hotmail.
com基金项目:北京市共建项目专项;北京市公园管理中心项目"菌根真菌对北京杓兰属植物的促生机制研究"引用格式:徐可,王涛,张毓.
兰科杓兰属(Cypripedium)植物种子非共生萌发研究进展[J].
生物资源,2020,42(1):4348.
XuK,WangT,ZhangY.
AdvancesinthestudyofasymbioticgerminationofCypripediumseeds[J].
BioticResources,2020,42(1):4348.
徐可等:兰科杓兰属(Cypripedium)植物种子非共生萌发研究进展高山,尤以四川和云南为多[4].
杓兰种子极为细小,无胚乳,自然条件下需与适宜的真菌共生,形成共生菌根才能萌发[5].
自然环境的破坏和人为过度采挖,加上种子休眠机制等自身因素,野生杓兰资源日趋减少,野外种群自然更新缓慢,已有11个种被列入世界自然保护联盟(internationalunionforconservationofnature,IUCN)濒危名录[6].
亟需采取有效的迁地和就地保育措施,非共生萌发是实现濒危杓兰属植物人工繁育,资源再生的重要途径.
本文将从以下方面对杓兰种子非共生萌发研究进展进行概述.
2杓兰属植物非共生萌发研究为尽可能多地保存遗传多样性的丰富度,以保育为目的的繁殖方式,常常采用有性繁殖方式——种子播种.
兰科植物种子萌发有非共生萌发(无真菌参与)和共生萌发(与真菌共生)两条途径.
其中非共生萌发是指由人工培养基提供营养,不需要任何真菌侵染就可以使杓兰种子萌发的繁殖方式,它能够在短期内获得一定数量的,具有遗传差异的植株群体[7],是目前兰科植物人工繁殖的主流繁育方式.
2.
1种子非共生萌发的影响因素杓兰种子萌发成功与否和萌发率的影响因素很多,一方面受种子质量和发育成熟度影响[8~12];另一方面受培养条件的影响,如培养基的组成、有机添加物的种类和浓度、激素、pH值、活性炭和温度等.
2.
1.
1种子成熟度杓兰是典型的高山植物,种子在严冬来临前成熟,如果成熟后即萌发,幼嫩的个体会在低温中夭折,因此在漫长的进化过程中形成了以休眠来应对的自我保护机制.
杓兰种子特别微小,结构简单,胚乳缺失,仅由一个简单的球形胚和外面包被着的种皮构成[13].
杓兰的种子由于缺乏胚乳供给萌发生长早期所需要的充足养分,萌发时需要由外界提供营养帮助其种子萌发及早期的生长发育[14~16].
研究发现,来自温带和高山地区的杓兰比热带附生兰的萌发困难得多,一般需要特殊的培养基配方和培养条件[15].
为提高无菌萌发效率,确定最佳采种时期,对多种杓兰种子在不同发育阶段的内源激素水平进行了测定,认为高水平的脱落酸(abscisicacid,ABA)是抑制种子萌发的重要因素之一[17,18].
通过观察种子结构发现,杓兰成熟种子具有内外双层(珠被)种皮,且种皮木质化程度较高[19],这可能是杓兰进化中的一种保护性适应,但也成为导致杓兰成熟种子萌发困难的原因之一[4,18,20].
杓兰种子无菌条件下的离体培养进展缓慢,目前只有部分种类能顺利萌发.
大量研究表明,特定发育阶段的杓兰未成熟种子比成熟种子更易萌发[4,17,21].
Hsu等[22]研究表明,对叶杓兰(C.
debile)授粉150天后采集的成熟种子具有最佳萌发率(达75%),相同培养条件下,授粉后30~90天的未成熟种子未见萌发,120天未成熟种子萌发率为72.
7%.
然而Pauw和Remphrey[23]研究发现釉白杓兰(C.
candidum)授粉后8周采收的种子发芽率最高.
Lee等[24]采集授粉后60、90和135天的台湾杓兰(C.
formosanum)绿果荚进行无菌播种试验,结果表明授粉60天以前的种子不能萌发,授粉90天的种子萌发率最高(达70%).
张毓等[25]和邓莲等[26]的研究均表明,大花杓兰(C.
macranthos)授粉后56天的种子萌发率最高,萌发率可达68.
08%,相同培养条件下,授粉后发育时期低于28天和高于84天时种子萌发困难,萌发率为零.
Jiang等[4]研究表明,长瓣杓兰(C.
lentiginosum)在授粉后90~105天采集的未成熟种子具有最佳萌发率(可达57.
8%).
对大花杓兰不同发育时期的种子进行切片观察,认为离体萌发最佳采种期是胚体已发育成球形胚,但内珠被还未发育成内种皮的时期[25].
2.
1.
2培养基成分不同种的杓兰属植物的最适基本培养基各不相同,不同种类及同一种类不同时期在相同培养基上的萌发率也不同.
Peterson和Remphrey[23]等研究发现,不同培养基对釉白杓兰种子萌发的影响存在差异,主要受年份、采收日期和品种的影响.
朴仁哲等[27]将大花杓兰播种在五种培养基中(MS、Hyponex、VWD、Harvais、改良Harvais),结果显示Harvais培养基是大花杓兰萌发的优选培养基.
将未成熟(授粉后105d)的长瓣杓兰种子播种在(1/2,1/4,1/8和1/10)的MS培养基中,结果表明在1/4和1/2MS培养基上种子萌发率最高(分别为54.
1%和52.
8%)[4].
Huh等[28]将大花杓兰播种在2倍MS,MS,1/2MS,1/4MS和1/8MS的培养基中,结果显示,在1/4MS培养基下具有最高萌发率(68.
1%).
浓度过高的MS培养基对大花杓兰种子的萌发具有抑制作用,浓度过低营养不足也会影响种子的萌发和原球茎生成率.
糖类物质作为培养基的能源物质和渗透调节剂,对种子萌发和原球茎的生长影响较大,不同种类和浓度的糖源对杓兰萌发的作用不同.
Harvais[11]··44生物资源对皇后杓兰(C.
reginea)的萌发研究表明,葡萄糖、果糖和蔗糖三种糖源的浓度均为1%的萌发效果好于2%;在同一浓度下,蔗糖优于葡萄糖,葡萄糖优于果糖.
Huh等[29]的研究表明,将大花杓兰种子播种在不同蔗糖浓度(0,10,20,30,40g/L)的MS培养基中,在含有10g/L蔗糖的培养基上,萌发率最高,可达65.
8%.
2.
1.
3有机添加物兰花种子胚乳缺失,缺乏内源性营养物质供种子在萌发过程中使用,因此在常规植物组织培养基中添加适宜的天然有机物或者抽提物可以促进杓兰种子的萌发已得到众多研究证实[26,30~32].
目前常用于杓兰种子萌发的有机添加物包括椰子水、苹果匀浆、马铃薯匀浆液等天然物质.
虽然还没有关于椰子水促进兰花种子萌发的具体机理研究,但作为一种植物液体胚乳,常常是效果最佳、最常用的促进杓兰种子萌发的有机添加物.
Chu等[30]研究表明,将杓兰(C.
calceolusvar.
pubescens)种子播种在添加100ml/L椰子水的1/4MS培养基中,种子萌发的整齐度增强且可以促进原球茎的健康发育.
朴仁哲等[27]发现在改良的Harvais培养基中加入马铃薯匀浆液或苹果匀浆时,可以促进大花杓兰种子萌发.
Huh等[29]研究表明,将大花杓兰播种在不同椰子汁浓度(0,50,100,200ml/L)的MS培养基中,含有100ml/L椰子汁的基础培养基中萌发率(70.
8%)最高.
2.
1.
4激素不同植物激素的作用机理不同,且不同种杓兰萌发所需的外源激素种类和浓度也不同.
Miyoshi和Mii[33]研究表明,大花杓兰种子播种在加入植物激素(1/3MS+1μmol/L6BA)的培养基中,萌发率可达58%~70%,不加植物激素的对照组(1/3MS)萌发率仅17%.
黄家林和胡虹[34]的研究进一步证实,外源细胞分裂素可以作为一种萌发诱导物质直接起作用,明显促进种子萌发.
在黄花杓兰(C.
flavum)未成熟种子萌发中,外源细胞分裂素起明显的促进萌发作用,而且低浓度的KT(kinetin)和6BA(6benzylaminopurine)即可使黄花杓兰[34]、斑花杓兰(C.
guttatum)、大花杓兰[20,26,27]、长瓣杓兰[4]种子无菌萌发率成倍提高.
将未成熟(授粉后105d)的大花杓兰种子播种在补充有1μmol/L的2IP、6BA以及TDZ的培养基中,只有2IP对种子萌发具有促进作用[4].
对多种杓兰种子在不同发育阶段的内源激素水平进行测定,认为高水平的ABA是抑制种子萌发的重要因素之一[17,18],因此越来越多的研究认为,植物激素促进种子萌发的机理是激素拮抗ABA的抑制作用而促使种子萌发.
由于较高浓度的植物激素对杓兰幼苗的分化有一定的负面影响,因此,在不添加激素能达到萌发目的的条件下,尽量避免使用或者少用植物激素来促进种子的萌发[35].
在必需激素帮助种子萌发的情况下,萌发以后也要尽快转移到不含激素的培养基中.
2.
1.
5培养方式液体培养可以稀释兰花种皮中产生的抑制物质而促进萌发[8].
Chu和Mudge[30]对杓兰(C.
calceolusvar.
pubescens)的研究发现,液体培养能代替低温处理提高种子的萌发率,同时增加种子萌发的同步性.
1/4MS液体培养基比固体培养基更能促进对叶杓兰成熟种子萌发[22].
至于哪种培养方式更适合杓兰种子的萌发,还需进一步研究.
2.
1.
6pH值大多数杓兰植物非共生萌发的研究表明,培养基pH值通常设置在5.
5~5.
7[4,30,36,37].
但是Oliva和Arditti[38]的研究表明,加州杓兰(C.
californicum)和山地杓兰(C.
montanum)在pH值为7.
0~7.
5的范围内萌发率最高.
皇后杓兰种子在pH4.
5的培养基中萌发和生长状态优于pH5.
5的培养基[12].
2.
1.
7活性炭Curtis[39]尝试在兰花培养基中加入活性炭,随后越来越多杓兰相关研究证明,在培养基中加入活性炭可以提高一些杓兰种类的萌发率[32,36,37].
但是,不同的杓兰种类也有差异.
比如Chu等[30]研究表明,活性炭对杓兰(C.
calceolusvar.
pubescens)种子萌发没有显著影响.
活性炭不仅可以吸收酚类等培养过程中产生的代谢物质,而且在一定程度上营造部分黑暗环境,并维持pH值相对稳定[40].
此外,活性炭可以改善通气并添加微量元素[41],这有利于种子萌发和幼苗生长.
同时,活性炭也会吸附某些化合物,包括某些维生素和植物激素,降低他们的浓度,这可能对种子非共生萌发和幼苗的生长产生负面的影响[42].
因此针对不同的植物种类和发育阶段,活性炭的使用及浓度需经过试验进行优选.
2.
2预处理对成熟种子非共生萌发的影响杓兰的种子在成熟后可以有长达数年的休眠期[16],在人工繁育中,这些保护机制则成为需要逾越的障碍.
打破杓兰种子休眠的方法有很多,前人也进行了大量的相关研究.
2.
2.
1适度破坏种皮成熟的杓兰植物种子可以通过腐蚀种皮来提高··45徐可等:兰科杓兰属(Cypripedium)植物种子非共生萌发研究进展萌发率,比如用NaClO或Ca(ClO)对种子进行预处理.
Lauzer等[43]的研究表明,在播种前对基叶杓兰(C.
acaule)种子进行NaClO预处理是影响种子萌发的重要因素.
NaClO预处理的时间由40min提高到2h,其萌发率也由3.
9%提高到10.
4%.
Miyoshi等[33]研究表明,用0.
5%NaClO处理大花杓兰1h,或用3.
2%的Ca(ClO)处理7h,萌发率可达到70%.
但是,当处理时间达到5h时,种子则不再萌发,原因可能是NaClO长时间接触种子,使种子内部的种胚失去了活性.
Bae等[36]用1%的NaClO处理成熟的斑花杓兰和大花杓兰种子30min,萌发率分别为77.
3%和27.
8%.
将长瓣杓兰成熟种子在播种前用0.
5%或1%NaOCl预处理15、30、45或60min,结果表明45min时萌发率最高(56.
9±6.
4%,60.
3±6.
1%)[4].
2.
2.
2低温处理研究发现,低温处理是打破成熟种子休眠的有效方式之一.
对皇后杓兰的研究表明,成熟种子必须通过2个月以上的低温处理才能打破休眠萌发[10].
同样,对杓兰(C.
calceolusvar.
pubescens)的研究表明,将种子放在5℃低温处理8周,萌发率从44.
8%提高到91.
7%[30].
低温处理可以影响大花杓兰和扇脉杓兰(C.
japonicum)种子的萌发[44].
将杓兰(C.
macranthosvar.
rebunense)成熟种子在4°C条件下处理3个月后,通过添加NAA和细胞分裂素的改良Hyponexpeptone培养基进行萌发培养成功获得杓兰原球茎[45].
但同时也有研究表明,低温处理并不能提高杓兰种子的萌发率,有时甚至起抑制作用.
不同条件的冷处理对釉白杓兰萌发的影响是可变的[15].
对基叶杓兰研究表明,灭菌播种后的种子在萌发培养基上做低温处理6个月,结果萌发率并未提高[43,46],说明杓兰属不同种类萌发习性差异大,不同的原生种萌发要求不同,这也是杓兰繁育习性的一个特点.
3展望对于杓兰属植物种子非共生萌发,研究者要解决的问题是如何使种子高效萌发,围绕这一目标,需要探究的是种子萌发过程中需要哪些营养物质、何种培养条件,分析不同种类、生境的杓兰属植物种子萌发的需求异同点,并从中总结规律,优化杓兰种子繁育技术体系.
以杓兰属植物为代表的温带(高山)兰科植物是繁殖和栽培都十分困难的类群,通过对杓兰属植物非共生萌发繁育体系的研究,有利于为高山兰科植物种子萌发提供指导依据,推动珍稀濒危高山兰科植物的保育研究工作.
参考文献[1]罗毅波,贾建生,王春玲.
中国兰科植物保育的现状和展望[J].
生物多样性,2003,11(1):7077.
LuoYB,JiaJS,WangCL.
AgeneralreviewoftheconservationstatusofChineseorchids[J].
ChinBiodivers,2003,11(1):7077.
[2]CribbP,SandisonMS.
ApreliminaryassessmentoftheconservationstatusofCypripediumspeciesinthewild[J].
BotJLinnSoc,1998,126:183190.
[3]WuZY,RavenPH,HongDY.
eds.
FloraofChina.
Vol.
25(Orchidaceae)[M].
Beijing:SciencePressandSt.
Louis:MissouriBotanicalGardenPress,2009:2233.
[4]JiangH,ChenMC,LeeYI.
InvitrogerminationandlowtemperatureseedstorageofCypripediumlentiginosumP.
J.
Cribb&S.
C.
Chen,arareandendangeredlady'sslipperorchid[J].
SciHortic,2017,225:471479.
[5]安曼云.
云南杓兰菌根真菌组成及共生关系研究[J].
广西植物,2017,37(6):763767.
AnMY.
CompositionofmycorrhizalfungiandsymbioticrelationshipofCypripediumyunnanense[J].
Guihaia,2017,37(6):763767.
[6]刘思思,陈娟,郭顺星.
兰科植物种子萌发的研究进展[J].
种子,2015,34:4350.
LiuSS,ChenJ,GuoSX.
Reviewongerminationoforchidseeds[J].
Seed,2015,34:4350.
[7]朱泉,田甜,杨澍,等.
兰科植物种子的非共生萌发研究进展[J].
江苏农业科学,2009,37(4):205208.
ZhuQ,TianT,YangS,etal.
Reviewonnonasymbioticgerminationoforchidseeds[J].
JiangsuAgriSci,2009,37(4):205208.
[8]ArdittiJ.
Factorsaffectingthegerminationoforchidseeds[J].
BotRev,1967,33(1):197.
[9]ArdittiJ.
Micropropagationoforchids,VolumeI–II.
2nded[M].
Oxford:BlackwellPublishingLtd,2008.
[10]BallardWW.
SterilepropagationofCypripediumreginaefromseeds[J].
AmerOrchidSocBull,1987,56:935946.
[11]HarvaisG.
GrowthrequirementsanddevelopmentofCypripediumreginaeinaxenicculture[J].
CanJBot,1973,51(2):327332.
[12]HarvaisG.
AnimprovedculturemediumforgrowingtheorchidCypripediumreginaeaxenically[J].
CanJBot,1982,60(12):25472555.
[13]胡适宜.
被子植物胚胎学[M].
北京:人民教育出版社,1982:103113.
HuSY.
Embryologyofangiosperms[M].
Beijing:Peo··46生物资源ple'sEducationPress,1982:103113.
[14]ArdittiJ,PridgeonA.
Orchidbiology:reiewsandperspectives[M].
London:KIuwerAcademicPublisher,1997:3273.
[15]PetersonRL,UetakeY,ZelmerC.
Fungalsymbioseswithorchidprotocorms[J].
Symbiosis1998,25(1):2955.
[16]ArdittiJ,ErnstR.
Physiologyofgerminatingorchidseeds.
In:ArdittiJ,edOrchidbiology:reviewsandperspectivesIII[M].
Ithaca,NewYork:CornellUniversityPress,1984:176222.
[17]LeeYI,ChungM,YeungEC,etal.
Dynamicdistributionandtheroleofabscisicacidduringseeddevelopmentofalady'sslipperorchid,Cypripediumformosanum[J].
AnnBot,2015,116(3):403411.
[18]YanXN,TianM,LiuF,etal.
HormonalandmorphologicalchangesduringseeddevelopmentofCypripediumjaponicum[J].
Protoplasma,2017,25460:23152322.
[19]BarsbergST,LeeYI,RasmussenHN.
DevelopmentofCligninwithG/SligninandlipidsinorchidseedcoatsanunexpecteddiversityexposedbyATRFTIRspectroscopy[J].
SeedSciRes,2018,28(1):4151.
[20]ZhangY,LeeYI,DengL,etal.
AsymbioticgerminationofimmatureseedsandtheseedlingdevelopmentofCypripediummacranthosSw.
anendangeredlady'sslipperorchid[J].
SciHortic,2013,164:130136.
[21]WagnerJ,HanselA.
InvitroseedgerminationofCypripediumcalceolusL.
atvariousembryogenicstages[J].
AngewBot,1994,68:59.
[22]HsuRCC,LeeYI.
SeeddevelopmentofCypripediumdebilerchb.
f.
inrelationtoasymbioticgermination[J].
Horts,2012,47(10):14951498.
[23]DePauwMA,RemphreyWR.
InvitrogerminationofthreeCypripediumspeciesinrelationtotimeofseedcollection,media,andcoldtreatment[J].
CanJBot,1993,71(6):879885.
[24]LeeYI,LeeN,YeungEC,etal.
EmbryodevelopmentofCypripediumformosanuminrelationtoseedgerminationinvitro[J].
JAmSocHorticSci,2005,130(5):747753.
[25]张毓,张启翔,赵世伟,等.
濒危植物大花杓兰胚与珠被发育的研究[J].
园艺学报,2010,37(1):7276.
ZhangY,ZhangQX,ZhaoSW,etal.
EmbryoandintegumentdevelopmentoftheendangeredspeciesCypripediummacranthosSw.
[J].
ActaHorticulturaeSinica,2010,37(1):7276.
[26]邓莲,张毓,王苗苗,等.
濒危兰科植物大花杓兰种子非共生萌发的研究[J].
种子,2012,31(6):3134,39.
DengL,ZhangY,WangMM,etal.
StudyonnonasymbioticgerminationofCypripediummacranthosseed[J].
Seed,2012,31(6):3134,39.
[27]朴仁哲,王艳丽,赵洪颜.
濒危植物大花杓兰种子离体培养条件初探[J].
安徽农业科学,2011,39(30):1842818429,18445.
PiaoRZ,WangYL,ZhaoHY.
PreliminaryexplorationoninvitroseedcultureconditionsofCypripediummacranthosSw.
[J].
JAnhuiAgriSci,2011,39(30):1842818429,18445.
[28]HuhYS,LeeJK,NamSY,etal.
EffectsofalteringmediumstrengthandsucroseconcentrationoninvitrogerminationandseedlinggrowthofCypripediummacranthosSw.
withorganicadditives[J].
JPlantBiotechnol,2016,43:138145.
[29]HuhYS,LeeJK,NamSY,etal.
ImprovementofasymbioticseedgerminationandseedlingdevelopmentofCypripediummacranthosSw.
withorganicadditives[J].
JPlantBiotechnol,2016,43(1):138145.
[30]ChuCC,MudgeKW.
Effectsofprechillingandliquidsuspensioncultureonseedgerminationoftheyellowlady'sslipperorchid,Cypripediumcalceoclusvar.
pubescens[J].
Lindleyana,1994,9:153159.
[31]SteeleWK.
GrowingCypripediumreginaefromseed[J].
AmOrchidSocBull,1995,64:383391.
[32]YanN,HuH,HuangJL,etal.
MicropropagationofCypripediumflavumthroughmultipleshootsofseedlingsderivedfrommatureseeds[J].
PlantCellTissOrganCult,2006,84(1):114118.
[33]MiyoshiK,MiiM.
Stimulatoryeffectsofsodiumandcalciumhypochlorite,prechillingandcytokininsonthegerminationofCypripediummacranthosseedinvitro[J].
PhysiolPlant,1998,102(4):481486.
[34]黄家林,胡虹.
黄花杓兰种子无菌萌发的培养条件研究[J].
云南植物研究,2001,23(1):105108.
HuangJL,HuH.
SeedgerminationrequirementsofCypripediumflavuminaxenicculture[J].
ActaBotankaYunnanica,2001,23(1):105108.
[35]SteeleWK.
LargescaleseedlingproductionofNorthAmericanCypripediumspecies.
InC.
Allened,NorthAmericanNativeTerrestrialOrchidPropagationandProduction[C].
N.
AmericanNativeTerrestrialOrchidConference,Germantown,Maryland.
1996:234242.
[36]BaeKH,KwonHK,ChoiYE.
InvitrogerminationandplantletconversionfromthecultureoffullymatureseedsofCypripediumguttatumSwartz.
[J].
PropagationofOrnamentalPlants,2009,9(3):160165.
[37]BaeKH,KimCH,SunBY,etal.
Structuralchangesofseedcoatsandstimulationofinvitrogermination··47徐可等:兰科杓兰属(Cypripedium)植物种子非共生萌发研究进展offullymatureseedsofCypripediummacranthosSwartz(Orchidaceae)byNaOClpretreatment[J].
PropagOrnamPlants,2010,10(2):107113.
[38]OlivaAP,ArdittiJ.
SeedgerminationofNorthAmericanorchidsII.
nativeCaliforniaandrelatedspeciesofAplectrum,CypripediumandSpiranthes[J].
BotGazette,1984,145(4):495501.
[39]CurtisJT.
GerminationandseedlingdevelopmentinfivespeciesofCypripedium[J].
AmJBot,1943,30(3):199.
[40]RasmussenHN.
Terrestrialorchidsfromseedtomycotrophicplant[M].
NewYork:CambridgeUniversityPress,1995.
[41]JohanssonLB,CallebergE,GedinA.
Correlationsbetweenactivatedcharcoal,FeEDTAandotherorganicmediaingredientsinculturedanthersofAnemonecanadensis[J].
PhysiolPlant,1990,80(2):243249.
[42]YamTW,ErnstR,ArdittiJ,etal.
Charcoalinorchidseedandtissueculturemedia:areview[J].
Lindleyana,1990,5:256265.
[43]LauzerD,StArnaudM,BarabeD.
TetrazoliumstainingandinvitrogerminationofmatureseedsofCypripediumacaule(Orchidaceae)[J].
Lindleyana,1994,9:197204.
[44]TomitaM,TomitaM.
EffectsofculturemediaandcoldtreatmentongerminationinasymbioticcultureofCypripediummacranthosandCypripediumjaponicum[J].
Lindleyana,1997,12:208213.
[45]ShimuraH,KodaY.
MicropropagationofCypripediummacranthosvar.
rebunensethroughprotocormlikebodiesderivedfrommatureseeds[J].
PlantCellTissueOrganCult,2004,78(3):273276.
[46]StArnaudM,LauzerD,BarabeD.
InvitrogerminationandearlygrowthofseedlingsofCypripediumacaule(Orchidaceae)[J].
Lindleyana,1992,7:2227.
(编辑:杨晓翠)··48
北京市植物园北京市花卉园艺工程技术研究中心城乡生态环境北京实验室,北京100093;2.
天津师范大学地理与环境科学学院,天津300387)摘要:兰科杓兰属(Cypripedium)植物主要分布于东亚、北美等温带地区和亚热带山地.
杓兰不仅具有极高的观赏价值,而且其经济价值和科研价值也越来越受到人们的重视.
近年来对杓兰属植物人工繁殖的相关研究不断深入,主要集中于种子的非共生萌发等方面.
本文对濒危兰科杓兰属植物进行了简要介绍,并就其种子非共生萌发研究从种子成熟度、预处理和有机添加物的作用、培养基的配置等方面进行综述,为目标杓兰种类的非共生萌发试验方案的制定奠定基础,将有助于温带/高山兰科植物保育研究的发展.
关键词:杓兰属;非共生萌发;离体培养;研究进展中图分类号:Q943文献标识码:A文章编号:20963491(2020)01004306AdvancesinthestudyofasymbioticgerminationofCypripediumseedsXUKe1,2,WANGTao1,ZHANGYu1*(1.
BeijingFloricultureEngineeringTechnologyResearchCentre,BeijingLaboratoryofUrbanandRuralEcologicalEnvironment,BeijingBotanicalGarden,Beijing100093,China;2.
SchoolofGeographicandEnvironmentalSciences,TianjinNormalUniversity,Tianjin300387,China)Abstract:CypripediumplantsaremainlydistributedintemperateAsiaandNorthAmerica.
Theynotonlyhaveveryhighornamentalvalue,butalsotheireconomicvalueandscientificresearchvaluearemoreandmoreconcernedbypeople.
Inrecentyears,researchesonpropagationofCypripediumhavebeendeepened,mainlyfocusonasymbioticgerminationoftheseeds.
ThispaperbrieflyintroducestheendangeredorchidsofCypripedium,andsummarizestheresearchprogressofseedasymbioticgerminationfromaspectsofseedmaturity,pretreatment,theroleoforganicadditives,andtheconfigurationofmedia.
Thestudylaysthefoundationfortheasymbioticgerminationexperimentprogramoftheorchidspecies,andpromotestheresearchanddevelopmentoforchidplantconservation.
Keywords:Cypripedium;asymbioticgermination;invitroculture;researchprogress1杓兰属植物简介杓兰属(Cypripedium)植物属兰科(Orchidaceae)植物大家族,为多年生宿根花卉,因其花唇瓣特化成兜状、杓状或拖鞋状而得名,西方人称为拖鞋兰,即"lady'sslipper",台湾称作"仙履兰".
杓兰独特的外形,高观赏价值以及在植物保育中的旗舰地位,被称为"植物中的大熊猫"[1].
杓兰属植物全世界约有50种,北温带地区主要分布于温带的亚洲和北美洲,南至喜马拉雅地区和中美洲[2,3].
我国是杓兰属植物的世界分布中心,共有36种,其中25种为我国特有种,广泛分布于东北至西南的山地和台湾DOI:10.
14188/j.
ajsh.
2020.
01.
006收稿日期:20191009修回日期:20191120接受日期:20191203作者简介:徐可(1995-),男,硕士,主要从事园林植物研究.
E-mail:xuke19951010@126.
com通讯联系人:张毓(1971-),女,博士,教授级高级工程师,主要从事珍稀濒危植物保护研究.
E-mail:bjyzhang@hotmail.
com基金项目:北京市共建项目专项;北京市公园管理中心项目"菌根真菌对北京杓兰属植物的促生机制研究"引用格式:徐可,王涛,张毓.
兰科杓兰属(Cypripedium)植物种子非共生萌发研究进展[J].
生物资源,2020,42(1):4348.
XuK,WangT,ZhangY.
AdvancesinthestudyofasymbioticgerminationofCypripediumseeds[J].
BioticResources,2020,42(1):4348.
徐可等:兰科杓兰属(Cypripedium)植物种子非共生萌发研究进展高山,尤以四川和云南为多[4].
杓兰种子极为细小,无胚乳,自然条件下需与适宜的真菌共生,形成共生菌根才能萌发[5].
自然环境的破坏和人为过度采挖,加上种子休眠机制等自身因素,野生杓兰资源日趋减少,野外种群自然更新缓慢,已有11个种被列入世界自然保护联盟(internationalunionforconservationofnature,IUCN)濒危名录[6].
亟需采取有效的迁地和就地保育措施,非共生萌发是实现濒危杓兰属植物人工繁育,资源再生的重要途径.
本文将从以下方面对杓兰种子非共生萌发研究进展进行概述.
2杓兰属植物非共生萌发研究为尽可能多地保存遗传多样性的丰富度,以保育为目的的繁殖方式,常常采用有性繁殖方式——种子播种.
兰科植物种子萌发有非共生萌发(无真菌参与)和共生萌发(与真菌共生)两条途径.
其中非共生萌发是指由人工培养基提供营养,不需要任何真菌侵染就可以使杓兰种子萌发的繁殖方式,它能够在短期内获得一定数量的,具有遗传差异的植株群体[7],是目前兰科植物人工繁殖的主流繁育方式.
2.
1种子非共生萌发的影响因素杓兰种子萌发成功与否和萌发率的影响因素很多,一方面受种子质量和发育成熟度影响[8~12];另一方面受培养条件的影响,如培养基的组成、有机添加物的种类和浓度、激素、pH值、活性炭和温度等.
2.
1.
1种子成熟度杓兰是典型的高山植物,种子在严冬来临前成熟,如果成熟后即萌发,幼嫩的个体会在低温中夭折,因此在漫长的进化过程中形成了以休眠来应对的自我保护机制.
杓兰种子特别微小,结构简单,胚乳缺失,仅由一个简单的球形胚和外面包被着的种皮构成[13].
杓兰的种子由于缺乏胚乳供给萌发生长早期所需要的充足养分,萌发时需要由外界提供营养帮助其种子萌发及早期的生长发育[14~16].
研究发现,来自温带和高山地区的杓兰比热带附生兰的萌发困难得多,一般需要特殊的培养基配方和培养条件[15].
为提高无菌萌发效率,确定最佳采种时期,对多种杓兰种子在不同发育阶段的内源激素水平进行了测定,认为高水平的脱落酸(abscisicacid,ABA)是抑制种子萌发的重要因素之一[17,18].
通过观察种子结构发现,杓兰成熟种子具有内外双层(珠被)种皮,且种皮木质化程度较高[19],这可能是杓兰进化中的一种保护性适应,但也成为导致杓兰成熟种子萌发困难的原因之一[4,18,20].
杓兰种子无菌条件下的离体培养进展缓慢,目前只有部分种类能顺利萌发.
大量研究表明,特定发育阶段的杓兰未成熟种子比成熟种子更易萌发[4,17,21].
Hsu等[22]研究表明,对叶杓兰(C.
debile)授粉150天后采集的成熟种子具有最佳萌发率(达75%),相同培养条件下,授粉后30~90天的未成熟种子未见萌发,120天未成熟种子萌发率为72.
7%.
然而Pauw和Remphrey[23]研究发现釉白杓兰(C.
candidum)授粉后8周采收的种子发芽率最高.
Lee等[24]采集授粉后60、90和135天的台湾杓兰(C.
formosanum)绿果荚进行无菌播种试验,结果表明授粉60天以前的种子不能萌发,授粉90天的种子萌发率最高(达70%).
张毓等[25]和邓莲等[26]的研究均表明,大花杓兰(C.
macranthos)授粉后56天的种子萌发率最高,萌发率可达68.
08%,相同培养条件下,授粉后发育时期低于28天和高于84天时种子萌发困难,萌发率为零.
Jiang等[4]研究表明,长瓣杓兰(C.
lentiginosum)在授粉后90~105天采集的未成熟种子具有最佳萌发率(可达57.
8%).
对大花杓兰不同发育时期的种子进行切片观察,认为离体萌发最佳采种期是胚体已发育成球形胚,但内珠被还未发育成内种皮的时期[25].
2.
1.
2培养基成分不同种的杓兰属植物的最适基本培养基各不相同,不同种类及同一种类不同时期在相同培养基上的萌发率也不同.
Peterson和Remphrey[23]等研究发现,不同培养基对釉白杓兰种子萌发的影响存在差异,主要受年份、采收日期和品种的影响.
朴仁哲等[27]将大花杓兰播种在五种培养基中(MS、Hyponex、VWD、Harvais、改良Harvais),结果显示Harvais培养基是大花杓兰萌发的优选培养基.
将未成熟(授粉后105d)的长瓣杓兰种子播种在(1/2,1/4,1/8和1/10)的MS培养基中,结果表明在1/4和1/2MS培养基上种子萌发率最高(分别为54.
1%和52.
8%)[4].
Huh等[28]将大花杓兰播种在2倍MS,MS,1/2MS,1/4MS和1/8MS的培养基中,结果显示,在1/4MS培养基下具有最高萌发率(68.
1%).
浓度过高的MS培养基对大花杓兰种子的萌发具有抑制作用,浓度过低营养不足也会影响种子的萌发和原球茎生成率.
糖类物质作为培养基的能源物质和渗透调节剂,对种子萌发和原球茎的生长影响较大,不同种类和浓度的糖源对杓兰萌发的作用不同.
Harvais[11]··44生物资源对皇后杓兰(C.
reginea)的萌发研究表明,葡萄糖、果糖和蔗糖三种糖源的浓度均为1%的萌发效果好于2%;在同一浓度下,蔗糖优于葡萄糖,葡萄糖优于果糖.
Huh等[29]的研究表明,将大花杓兰种子播种在不同蔗糖浓度(0,10,20,30,40g/L)的MS培养基中,在含有10g/L蔗糖的培养基上,萌发率最高,可达65.
8%.
2.
1.
3有机添加物兰花种子胚乳缺失,缺乏内源性营养物质供种子在萌发过程中使用,因此在常规植物组织培养基中添加适宜的天然有机物或者抽提物可以促进杓兰种子的萌发已得到众多研究证实[26,30~32].
目前常用于杓兰种子萌发的有机添加物包括椰子水、苹果匀浆、马铃薯匀浆液等天然物质.
虽然还没有关于椰子水促进兰花种子萌发的具体机理研究,但作为一种植物液体胚乳,常常是效果最佳、最常用的促进杓兰种子萌发的有机添加物.
Chu等[30]研究表明,将杓兰(C.
calceolusvar.
pubescens)种子播种在添加100ml/L椰子水的1/4MS培养基中,种子萌发的整齐度增强且可以促进原球茎的健康发育.
朴仁哲等[27]发现在改良的Harvais培养基中加入马铃薯匀浆液或苹果匀浆时,可以促进大花杓兰种子萌发.
Huh等[29]研究表明,将大花杓兰播种在不同椰子汁浓度(0,50,100,200ml/L)的MS培养基中,含有100ml/L椰子汁的基础培养基中萌发率(70.
8%)最高.
2.
1.
4激素不同植物激素的作用机理不同,且不同种杓兰萌发所需的外源激素种类和浓度也不同.
Miyoshi和Mii[33]研究表明,大花杓兰种子播种在加入植物激素(1/3MS+1μmol/L6BA)的培养基中,萌发率可达58%~70%,不加植物激素的对照组(1/3MS)萌发率仅17%.
黄家林和胡虹[34]的研究进一步证实,外源细胞分裂素可以作为一种萌发诱导物质直接起作用,明显促进种子萌发.
在黄花杓兰(C.
flavum)未成熟种子萌发中,外源细胞分裂素起明显的促进萌发作用,而且低浓度的KT(kinetin)和6BA(6benzylaminopurine)即可使黄花杓兰[34]、斑花杓兰(C.
guttatum)、大花杓兰[20,26,27]、长瓣杓兰[4]种子无菌萌发率成倍提高.
将未成熟(授粉后105d)的大花杓兰种子播种在补充有1μmol/L的2IP、6BA以及TDZ的培养基中,只有2IP对种子萌发具有促进作用[4].
对多种杓兰种子在不同发育阶段的内源激素水平进行测定,认为高水平的ABA是抑制种子萌发的重要因素之一[17,18],因此越来越多的研究认为,植物激素促进种子萌发的机理是激素拮抗ABA的抑制作用而促使种子萌发.
由于较高浓度的植物激素对杓兰幼苗的分化有一定的负面影响,因此,在不添加激素能达到萌发目的的条件下,尽量避免使用或者少用植物激素来促进种子的萌发[35].
在必需激素帮助种子萌发的情况下,萌发以后也要尽快转移到不含激素的培养基中.
2.
1.
5培养方式液体培养可以稀释兰花种皮中产生的抑制物质而促进萌发[8].
Chu和Mudge[30]对杓兰(C.
calceolusvar.
pubescens)的研究发现,液体培养能代替低温处理提高种子的萌发率,同时增加种子萌发的同步性.
1/4MS液体培养基比固体培养基更能促进对叶杓兰成熟种子萌发[22].
至于哪种培养方式更适合杓兰种子的萌发,还需进一步研究.
2.
1.
6pH值大多数杓兰植物非共生萌发的研究表明,培养基pH值通常设置在5.
5~5.
7[4,30,36,37].
但是Oliva和Arditti[38]的研究表明,加州杓兰(C.
californicum)和山地杓兰(C.
montanum)在pH值为7.
0~7.
5的范围内萌发率最高.
皇后杓兰种子在pH4.
5的培养基中萌发和生长状态优于pH5.
5的培养基[12].
2.
1.
7活性炭Curtis[39]尝试在兰花培养基中加入活性炭,随后越来越多杓兰相关研究证明,在培养基中加入活性炭可以提高一些杓兰种类的萌发率[32,36,37].
但是,不同的杓兰种类也有差异.
比如Chu等[30]研究表明,活性炭对杓兰(C.
calceolusvar.
pubescens)种子萌发没有显著影响.
活性炭不仅可以吸收酚类等培养过程中产生的代谢物质,而且在一定程度上营造部分黑暗环境,并维持pH值相对稳定[40].
此外,活性炭可以改善通气并添加微量元素[41],这有利于种子萌发和幼苗生长.
同时,活性炭也会吸附某些化合物,包括某些维生素和植物激素,降低他们的浓度,这可能对种子非共生萌发和幼苗的生长产生负面的影响[42].
因此针对不同的植物种类和发育阶段,活性炭的使用及浓度需经过试验进行优选.
2.
2预处理对成熟种子非共生萌发的影响杓兰的种子在成熟后可以有长达数年的休眠期[16],在人工繁育中,这些保护机制则成为需要逾越的障碍.
打破杓兰种子休眠的方法有很多,前人也进行了大量的相关研究.
2.
2.
1适度破坏种皮成熟的杓兰植物种子可以通过腐蚀种皮来提高··45徐可等:兰科杓兰属(Cypripedium)植物种子非共生萌发研究进展萌发率,比如用NaClO或Ca(ClO)对种子进行预处理.
Lauzer等[43]的研究表明,在播种前对基叶杓兰(C.
acaule)种子进行NaClO预处理是影响种子萌发的重要因素.
NaClO预处理的时间由40min提高到2h,其萌发率也由3.
9%提高到10.
4%.
Miyoshi等[33]研究表明,用0.
5%NaClO处理大花杓兰1h,或用3.
2%的Ca(ClO)处理7h,萌发率可达到70%.
但是,当处理时间达到5h时,种子则不再萌发,原因可能是NaClO长时间接触种子,使种子内部的种胚失去了活性.
Bae等[36]用1%的NaClO处理成熟的斑花杓兰和大花杓兰种子30min,萌发率分别为77.
3%和27.
8%.
将长瓣杓兰成熟种子在播种前用0.
5%或1%NaOCl预处理15、30、45或60min,结果表明45min时萌发率最高(56.
9±6.
4%,60.
3±6.
1%)[4].
2.
2.
2低温处理研究发现,低温处理是打破成熟种子休眠的有效方式之一.
对皇后杓兰的研究表明,成熟种子必须通过2个月以上的低温处理才能打破休眠萌发[10].
同样,对杓兰(C.
calceolusvar.
pubescens)的研究表明,将种子放在5℃低温处理8周,萌发率从44.
8%提高到91.
7%[30].
低温处理可以影响大花杓兰和扇脉杓兰(C.
japonicum)种子的萌发[44].
将杓兰(C.
macranthosvar.
rebunense)成熟种子在4°C条件下处理3个月后,通过添加NAA和细胞分裂素的改良Hyponexpeptone培养基进行萌发培养成功获得杓兰原球茎[45].
但同时也有研究表明,低温处理并不能提高杓兰种子的萌发率,有时甚至起抑制作用.
不同条件的冷处理对釉白杓兰萌发的影响是可变的[15].
对基叶杓兰研究表明,灭菌播种后的种子在萌发培养基上做低温处理6个月,结果萌发率并未提高[43,46],说明杓兰属不同种类萌发习性差异大,不同的原生种萌发要求不同,这也是杓兰繁育习性的一个特点.
3展望对于杓兰属植物种子非共生萌发,研究者要解决的问题是如何使种子高效萌发,围绕这一目标,需要探究的是种子萌发过程中需要哪些营养物质、何种培养条件,分析不同种类、生境的杓兰属植物种子萌发的需求异同点,并从中总结规律,优化杓兰种子繁育技术体系.
以杓兰属植物为代表的温带(高山)兰科植物是繁殖和栽培都十分困难的类群,通过对杓兰属植物非共生萌发繁育体系的研究,有利于为高山兰科植物种子萌发提供指导依据,推动珍稀濒危高山兰科植物的保育研究工作.
参考文献[1]罗毅波,贾建生,王春玲.
中国兰科植物保育的现状和展望[J].
生物多样性,2003,11(1):7077.
LuoYB,JiaJS,WangCL.
AgeneralreviewoftheconservationstatusofChineseorchids[J].
ChinBiodivers,2003,11(1):7077.
[2]CribbP,SandisonMS.
ApreliminaryassessmentoftheconservationstatusofCypripediumspeciesinthewild[J].
BotJLinnSoc,1998,126:183190.
[3]WuZY,RavenPH,HongDY.
eds.
FloraofChina.
Vol.
25(Orchidaceae)[M].
Beijing:SciencePressandSt.
Louis:MissouriBotanicalGardenPress,2009:2233.
[4]JiangH,ChenMC,LeeYI.
InvitrogerminationandlowtemperatureseedstorageofCypripediumlentiginosumP.
J.
Cribb&S.
C.
Chen,arareandendangeredlady'sslipperorchid[J].
SciHortic,2017,225:471479.
[5]安曼云.
云南杓兰菌根真菌组成及共生关系研究[J].
广西植物,2017,37(6):763767.
AnMY.
CompositionofmycorrhizalfungiandsymbioticrelationshipofCypripediumyunnanense[J].
Guihaia,2017,37(6):763767.
[6]刘思思,陈娟,郭顺星.
兰科植物种子萌发的研究进展[J].
种子,2015,34:4350.
LiuSS,ChenJ,GuoSX.
Reviewongerminationoforchidseeds[J].
Seed,2015,34:4350.
[7]朱泉,田甜,杨澍,等.
兰科植物种子的非共生萌发研究进展[J].
江苏农业科学,2009,37(4):205208.
ZhuQ,TianT,YangS,etal.
Reviewonnonasymbioticgerminationoforchidseeds[J].
JiangsuAgriSci,2009,37(4):205208.
[8]ArdittiJ.
Factorsaffectingthegerminationoforchidseeds[J].
BotRev,1967,33(1):197.
[9]ArdittiJ.
Micropropagationoforchids,VolumeI–II.
2nded[M].
Oxford:BlackwellPublishingLtd,2008.
[10]BallardWW.
SterilepropagationofCypripediumreginaefromseeds[J].
AmerOrchidSocBull,1987,56:935946.
[11]HarvaisG.
GrowthrequirementsanddevelopmentofCypripediumreginaeinaxenicculture[J].
CanJBot,1973,51(2):327332.
[12]HarvaisG.
AnimprovedculturemediumforgrowingtheorchidCypripediumreginaeaxenically[J].
CanJBot,1982,60(12):25472555.
[13]胡适宜.
被子植物胚胎学[M].
北京:人民教育出版社,1982:103113.
HuSY.
Embryologyofangiosperms[M].
Beijing:Peo··46生物资源ple'sEducationPress,1982:103113.
[14]ArdittiJ,PridgeonA.
Orchidbiology:reiewsandperspectives[M].
London:KIuwerAcademicPublisher,1997:3273.
[15]PetersonRL,UetakeY,ZelmerC.
Fungalsymbioseswithorchidprotocorms[J].
Symbiosis1998,25(1):2955.
[16]ArdittiJ,ErnstR.
Physiologyofgerminatingorchidseeds.
In:ArdittiJ,edOrchidbiology:reviewsandperspectivesIII[M].
Ithaca,NewYork:CornellUniversityPress,1984:176222.
[17]LeeYI,ChungM,YeungEC,etal.
Dynamicdistributionandtheroleofabscisicacidduringseeddevelopmentofalady'sslipperorchid,Cypripediumformosanum[J].
AnnBot,2015,116(3):403411.
[18]YanXN,TianM,LiuF,etal.
HormonalandmorphologicalchangesduringseeddevelopmentofCypripediumjaponicum[J].
Protoplasma,2017,25460:23152322.
[19]BarsbergST,LeeYI,RasmussenHN.
DevelopmentofCligninwithG/SligninandlipidsinorchidseedcoatsanunexpecteddiversityexposedbyATRFTIRspectroscopy[J].
SeedSciRes,2018,28(1):4151.
[20]ZhangY,LeeYI,DengL,etal.
AsymbioticgerminationofimmatureseedsandtheseedlingdevelopmentofCypripediummacranthosSw.
anendangeredlady'sslipperorchid[J].
SciHortic,2013,164:130136.
[21]WagnerJ,HanselA.
InvitroseedgerminationofCypripediumcalceolusL.
atvariousembryogenicstages[J].
AngewBot,1994,68:59.
[22]HsuRCC,LeeYI.
SeeddevelopmentofCypripediumdebilerchb.
f.
inrelationtoasymbioticgermination[J].
Horts,2012,47(10):14951498.
[23]DePauwMA,RemphreyWR.
InvitrogerminationofthreeCypripediumspeciesinrelationtotimeofseedcollection,media,andcoldtreatment[J].
CanJBot,1993,71(6):879885.
[24]LeeYI,LeeN,YeungEC,etal.
EmbryodevelopmentofCypripediumformosanuminrelationtoseedgerminationinvitro[J].
JAmSocHorticSci,2005,130(5):747753.
[25]张毓,张启翔,赵世伟,等.
濒危植物大花杓兰胚与珠被发育的研究[J].
园艺学报,2010,37(1):7276.
ZhangY,ZhangQX,ZhaoSW,etal.
EmbryoandintegumentdevelopmentoftheendangeredspeciesCypripediummacranthosSw.
[J].
ActaHorticulturaeSinica,2010,37(1):7276.
[26]邓莲,张毓,王苗苗,等.
濒危兰科植物大花杓兰种子非共生萌发的研究[J].
种子,2012,31(6):3134,39.
DengL,ZhangY,WangMM,etal.
StudyonnonasymbioticgerminationofCypripediummacranthosseed[J].
Seed,2012,31(6):3134,39.
[27]朴仁哲,王艳丽,赵洪颜.
濒危植物大花杓兰种子离体培养条件初探[J].
安徽农业科学,2011,39(30):1842818429,18445.
PiaoRZ,WangYL,ZhaoHY.
PreliminaryexplorationoninvitroseedcultureconditionsofCypripediummacranthosSw.
[J].
JAnhuiAgriSci,2011,39(30):1842818429,18445.
[28]HuhYS,LeeJK,NamSY,etal.
EffectsofalteringmediumstrengthandsucroseconcentrationoninvitrogerminationandseedlinggrowthofCypripediummacranthosSw.
withorganicadditives[J].
JPlantBiotechnol,2016,43:138145.
[29]HuhYS,LeeJK,NamSY,etal.
ImprovementofasymbioticseedgerminationandseedlingdevelopmentofCypripediummacranthosSw.
withorganicadditives[J].
JPlantBiotechnol,2016,43(1):138145.
[30]ChuCC,MudgeKW.
Effectsofprechillingandliquidsuspensioncultureonseedgerminationoftheyellowlady'sslipperorchid,Cypripediumcalceoclusvar.
pubescens[J].
Lindleyana,1994,9:153159.
[31]SteeleWK.
GrowingCypripediumreginaefromseed[J].
AmOrchidSocBull,1995,64:383391.
[32]YanN,HuH,HuangJL,etal.
MicropropagationofCypripediumflavumthroughmultipleshootsofseedlingsderivedfrommatureseeds[J].
PlantCellTissOrganCult,2006,84(1):114118.
[33]MiyoshiK,MiiM.
Stimulatoryeffectsofsodiumandcalciumhypochlorite,prechillingandcytokininsonthegerminationofCypripediummacranthosseedinvitro[J].
PhysiolPlant,1998,102(4):481486.
[34]黄家林,胡虹.
黄花杓兰种子无菌萌发的培养条件研究[J].
云南植物研究,2001,23(1):105108.
HuangJL,HuH.
SeedgerminationrequirementsofCypripediumflavuminaxenicculture[J].
ActaBotankaYunnanica,2001,23(1):105108.
[35]SteeleWK.
LargescaleseedlingproductionofNorthAmericanCypripediumspecies.
InC.
Allened,NorthAmericanNativeTerrestrialOrchidPropagationandProduction[C].
N.
AmericanNativeTerrestrialOrchidConference,Germantown,Maryland.
1996:234242.
[36]BaeKH,KwonHK,ChoiYE.
InvitrogerminationandplantletconversionfromthecultureoffullymatureseedsofCypripediumguttatumSwartz.
[J].
PropagationofOrnamentalPlants,2009,9(3):160165.
[37]BaeKH,KimCH,SunBY,etal.
Structuralchangesofseedcoatsandstimulationofinvitrogermination··47徐可等:兰科杓兰属(Cypripedium)植物种子非共生萌发研究进展offullymatureseedsofCypripediummacranthosSwartz(Orchidaceae)byNaOClpretreatment[J].
PropagOrnamPlants,2010,10(2):107113.
[38]OlivaAP,ArdittiJ.
SeedgerminationofNorthAmericanorchidsII.
nativeCaliforniaandrelatedspeciesofAplectrum,CypripediumandSpiranthes[J].
BotGazette,1984,145(4):495501.
[39]CurtisJT.
GerminationandseedlingdevelopmentinfivespeciesofCypripedium[J].
AmJBot,1943,30(3):199.
[40]RasmussenHN.
Terrestrialorchidsfromseedtomycotrophicplant[M].
NewYork:CambridgeUniversityPress,1995.
[41]JohanssonLB,CallebergE,GedinA.
Correlationsbetweenactivatedcharcoal,FeEDTAandotherorganicmediaingredientsinculturedanthersofAnemonecanadensis[J].
PhysiolPlant,1990,80(2):243249.
[42]YamTW,ErnstR,ArdittiJ,etal.
Charcoalinorchidseedandtissueculturemedia:areview[J].
Lindleyana,1990,5:256265.
[43]LauzerD,StArnaudM,BarabeD.
TetrazoliumstainingandinvitrogerminationofmatureseedsofCypripediumacaule(Orchidaceae)[J].
Lindleyana,1994,9:197204.
[44]TomitaM,TomitaM.
EffectsofculturemediaandcoldtreatmentongerminationinasymbioticcultureofCypripediummacranthosandCypripediumjaponicum[J].
Lindleyana,1997,12:208213.
[45]ShimuraH,KodaY.
MicropropagationofCypripediummacranthosvar.
rebunensethroughprotocormlikebodiesderivedfrommatureseeds[J].
PlantCellTissueOrganCult,2004,78(3):273276.
[46]StArnaudM,LauzerD,BarabeD.
InvitrogerminationandearlygrowthofseedlingsofCypripediumacaule(Orchidaceae)[J].
Lindleyana,1992,7:2227.
(编辑:杨晓翠)··48
- 萌发郭杰非你莫属相关文档
- 王府郭杰非你莫属
- 武汉郭杰非你莫属
- 范畴郭杰非你莫属
- 會聞1111莫莫莫莫自自自自伐伐伐伐其其其其善善善善(之四)
- 厅属单位协助领导干部联系帮扶非公企业联络表
- 第一类非药品类易制毒化学品生产许可证核发(变更)
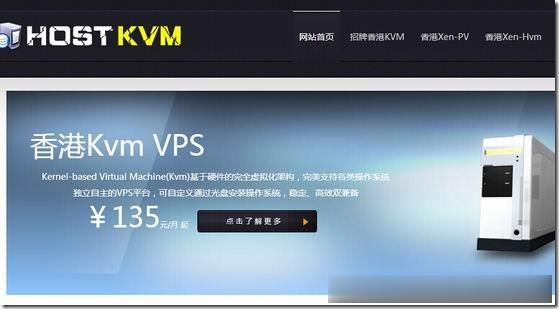
HostKvm5.95美元起,香港、韩国可选
HostKvm发布了夏季特别促销活动,针对香港国际/韩国机房VPS主机提供7折优惠码,其他机房全场8折,优惠后2GB内存套餐月付仅5.95美元起。这是一家成立于2013年的国外主机服务商,主要提供基于KVM架构的VPS主机,可选数据中心包括日本、新加坡、韩国、美国、中国香港等多个地区机房,均为国内直连或优化线路,延迟较低,适合建站或者远程办公等。下面分享几款香港VPS和韩国VPS的配置和价格信息。...
819云(240元)香港CN2 日本CN2 物理机 E5 16G 1T 20M 3IP
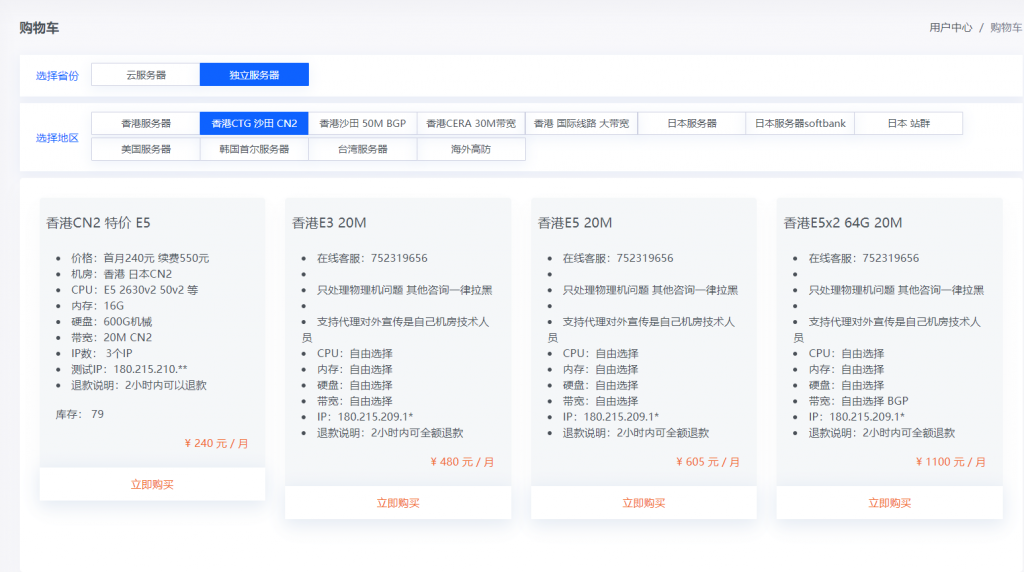
819云是我们的老熟人了,服务器一直都是稳定为主,老板人也很好,这次给大家带来了新活动,十分给力 香港CN2 日本CN2 物理机 E5 16G 1T 20M 3IP 240元0官方网站:https://www.819yun.com/ 特惠专员Q:442379204套餐介绍套餐CPU内存硬盘带宽IP价格香港CN2 (特价)E5 随机分配16G1T 机械20M3IP240元/月日本CN2 (...
TmhHost暑假活动:高端线路VPS季付8折优惠,可选洛杉矶CN2 GIA/日本软银/香港三网CN2 GIA/韩国双向CN2等
tmhhost怎么样?tmhhost正在搞暑假大促销活动,全部是高端线路VPS,现在直接季付8折优惠,活动截止时间是8月31日。可选机房及线路有美国洛杉矶cn2 gia+200G高防、洛杉矶三网CN2 GIA、洛杉矶CERA机房CN2 GIA,日本软银(100M带宽)、香港BGP直连200M带宽、香港三网CN2 GIA、韩国双向CN2。点击进入:tmhhost官方网站地址tmhhost优惠码:Tm...

郭杰非你莫属为你推荐
-
操作httpServicemy检索网易yeah全国企业信息查询全国企业信用信息公示系统查询入口 及操作说明哪里有?netshwinsockreset游戏出现battlEye Launcher 怎么办开放平台企鹅号和腾讯内容开放平台是一样的吗,有什么区别?购物车什么叫淘宝购物车腾讯官方电话腾讯公司电话多少电子商务世界世界前十大电子商务企业名字厦门三五互联科技股份有限公司厦门三五互联怎么样?