dBts60.com
ts60.com 时间:2021-04-10 阅读:()
ORIGINALPAPERHemisphericAsymmetryofAuditoryMismatchNegativityElicitedbySpectralandTemporalDeviants:AMagnetoencephalographicStudyHidehikoOkamotoRyusukeKakigiReceived:22October2013/Accepted:16December2013/Publishedonline:24December2013TheAuthor(s)2013.
ThisarticleispublishedwithopenaccessatSpringerlink.
comAbstractOneofthemajorchallengesinhumanbrainscienceisthefunctionalhemisphericasymmetryofaudi-toryprocessing.
Behavioralandneurophysiologicalstudieshavedemonstratedthatspeechprocessingisdominantlyhandledinthelefthemisphere,whereasmusicprocessingdominantlyoccursintheright.
Usingmagnetoencepha-lography,wemeasuredtheauditorymismatchnegativityelicitedbyband-passlteredclick-trains,whichdeviatedfromfrequentlypresentedstandardsoundsignalsinaspectralortemporaldomain.
Theresultsshowedthatspectralandtemporaldeviantsweredominantlyprocessedintherightandlefthemispheres,respectively.
Hemisphericasymmetrywasnotlimitedtohigh-levelcognitivepro-cesses,butalsooriginatedfromthepre-attentiveneuralprocessingstagerepresentedbymismatchnegativity.
KeywordsAuditoryevokedresponseHemisphericlateralityMagnetoencephalography(MEG)Mismatchnegativity(MMNm)AbbreviationsANOVAAnalysisofvarianceMEGMagnetoencephalographyMMNMismatchnegativitySDSpectraldeviantTDTemporaldeviantTSTeststimulusIntroductionFunctionalhemisphericasymmetryinthehumanbrainhasbeeninvestigatedsincethelatenineteenthcentury(Wer-nicke1874;Broca1861).
Inadditiontotheclassicalbehavioralobservationsofneurologicaldisorderpatients,recentneuroimagingtechniqueshavemadeitpossibletoinvestigateconscioushealthyhumanbrains,andhaverevealedlefthemisphericdominanceforspeechprocessingandrighthemisphericdominanceformusicprocessing(Zatorreetal.
1994,2002;Grifthsetal.
1999;Belinetal.
2000;Eulitzetal.
1995;Szymanskietal.
2001;Alhoetal.
1998).
However,functionalhemisphericasymmetryinthehumanbrainmaynotbelimitedtohigh-levelcognitiveneuralprocesses,butmaystartfromthelowerneuralprocessinglevelofbasicacousticfeatures(e.
g.
frequency,interval,duration,andintensity).
Naturalsoundshavespecicspectraldistributionsthatchangeovertimeaccordingtospecictemporalsequences.
Bothspectralandtemporalsoundfeatureshavebeenshowntoplayanimportantroleintheperceptionofnaturalsounds(Moore2003);however,theimportanceofthesefeaturesappearstodifferbetweensoundtypes,withspectralprocessingbeingofparticularimportanceformusicperception(VosandTroost1989;WarrierandZa-torre2002)andtemporalcuesbeingessentialforspeechperception(Shannonetal.
1995;Drullmanetal.
1994a,b).
Recentfunctionalmagneticresonanceimaging(Jamisonetal.
2006),positronemissiontomography(ZatorreandBelin2001),andmagnetoencephalography(MEG)ThisisoneofseveralpaperspublishedtogetherinBrainTopographyonthe''SpecialIssue:AuditoryCortex2012''.
ElectronicsupplementarymaterialTheonlineversionofthisarticle(doi:10.
1007/s10548-013-0347-1)containssupplementarymaterial,whichisavailabletoauthorizedusers.
H.
Okamoto(&)R.
KakigiDepartmentofIntegrativePhysiology,NationalInstituteforPhysiologicalSciences,38Nishigo-Naka,Myodaiji,Okazaki444-8585,Japane-mail:hokamoto@nips.
ac.
jp123BrainTopogr(2015)28:471–478DOI10.
1007/s10548-013-0347-1(Okamotoetal.
2009)studieshavedemonstratedusingarticialbasicauditorystimuli(e.
g.
puretonesandpulse-trains)thattemporalchangesaredominantlyprocessedinthelefthemisphere,whereasspectralchangesaredomi-nantlyprocessedintheright.
Thewell-knownfunctionalhumanhemisphericasymmetryobservedforspeechandmusicprocessingmaynotbelimitedtoconscioushigh-levelcognitiveprocesses,butmaybeatleastpartiallyrelatedtothepre-attentiveprocessingoflow-levelacousticfeatures.
Mismatchnegativity(MMN)anditsmagneticcounter-partMMNmareauditoryevokedcomponentsthatreectthecorticalpre-attentivediscriminationofauditorystimuliaswellasauditorymemorytraces(Na¨a¨ta¨nenetal.
1978,2007;Kujalaetal.
2007).
MMN(m)iselicitedbyviola-tionsofregularitiesinsoundstreamsandcanberecordedwithoutanymotororotherresponseandcanevenbeobtainedfrominattentivepatientsandinfants.
PreviousstudieshaveshownthatMMN(m)elicitedbyspeechsoundswassignicantlylateralizedtothelefthemisphere(Alhoetal.
1998),whereasMMN(m)elicitedbymusicalnoteswasdominantlyprocessedintherighthemisphere(Lappeetal.
2013;Tervaniemietal.
1999).
However,whetherthehemisphericasymmetriesofthepre-attentiveMMN(m)arelimitedtomeaningfulauditorystimuli(e.
g.
speechandmusic)ororiginatefromthebasicspectralandtemporalsoundfeaturesofthesesoundstimuliremainsunknown.
Therefore,theaimofthepresentstudywastoinvesti-gatethehemisphericlateralityofMMNmevokedbyspectralversustemporalsounddeviantsthatdonotconveyspecicphonological,grammatical,ormusicalinforma-tion.
InordertoexcludethepossibilitythatthelateralityoftheMMNmoriginatedfromthesoundstimulusitself,wecounter-balancedtotalauditoryinputsidenticalbetweenspectralandtemporaldeviantconditions.
Theresultsofthepresentstudyprovideanewinsightintohowtheleftandrighthemispherespre-attentivelydealwiththespectralandtemporalfeaturesofnaturalsoundsignals.
MaterialsandMethodsSubjectsThirteenhealthysubjectsparticipatedinthisstudy(vefemales;mean±standarddeviation:32.
1±6.
2years).
Allparticipantshadnormalhearing,hadnohistoryofpsychologicalorneurologicaldisorders,andwereunam-biguouslyright-handed[assessedviatheJapaneseversionof''EdinburghHandednessInventory''(Oldeld1971)].
AllparticipantswerefullyinformedaboutthestudyandgavewritteninformedconsentfortheirparticipationinaccordancewiththeproceduresapprovedbytheEthicsCommissionoftheNationalInstituteforPhysiologicalSciences,Okazaki,Japan.
ThestudyconformedtotheCodeofEthicsoftheWorldMedicalAssociation(Decla-rationofHelsinki).
StimuliandExperimentalDesignTheexperimentaldesignisschematicallyrepresentedinFig.
1.
Theteststimulus(TS)waseithera30Hz(TS30)or60Hz(TS60)click-train,whichwasone-octaveband-passlteredeitherbetween500and1,000Hz(TS30_Low(SupplementaryAudio1S)andTS60_Low(SupplementaryAudio2S))orbetween1,000and2,000Hz[TS30_High(SupplementaryAudio3S)andTS60_High(SupplementaryAudio4S)].
TheTShadadurationof330msandthesoundonsetasynchronybetweentheTSwas1,300ms.
OneoftheTSwerepresentedasstandardstimuliwith70%probabilitypseudo-randomlyintermixedwithspectraldeviants(SD:15%probability)andtemporaldeviants(TD:15%proba-bility)inanoddballsequenceasdemonstratedinFig.
1.
IncaseofSD,band-passltersettingschangedfromthestandardstimulus,whilethetypeoftheclick-trainremainedidentical(standardsandSD:TS30_LowandTS30_High,TS30_HighandTS30_Low,TS60_LowandTS60_High,TS60_HighandTS60_Low).
Ontheotherhand,incaseofTD,theltersettingsremainedidentical,whilethetypeofclick-trainchangedfromthestandardsoundstimulus(standardsandTD:TS30_LowandTS60_Low,TS30_HighandTS60_High,TS60_LowandTS30_Low,TS60_HighandTS30_High).
Morethantwostandardstimuliwerepresentedbeforeadeviantstimulus(SDorTD).
EachMEGsessionconsistedoffourblocks.
Eachblockcontainedfoursub-blocksthatpseudo-randomlyadoptedTS30_Low,TS30_High,TS60_Low,andTS60_HighasthestandardTS,respectively.
Consequently,allTStypeswerepresentedwithaprobabilityof25%inoneblock.
Eachsub-blockhad21SD,21TD,and98standardstimuli,resultinginatotalnumberof336trialsforeachdeviantstimulusand1,568trialsforthestandardcondition.
Allsoundsweredioticallypresentedthroughplastictubes1.
5minlengthandear-piecesttedtothesubject'sears.
BeforestartinganMEGmeasurement,eachsubject'shearingthresholdforTS30_Lowwasindividuallydeterminedforeachear.
Dur-ingtheMEGrecordingsession,TS30_Lowwaspresentedatanintensityof60dBabovetheindividualsensationlevel,andotherTSwereadjustedtohavepoweridenticaltoTS30_Low.
Inordertokeepthetestsubjectsalertanddis-tractedfromtheauditorysignals,aself-chosensilentmoviewithcaptionswaspresentedduringtheMEGrecordings.
Questionsregardingthecontentofthemoviewereaskedattheendofthemeasurementtoensurethatthesubjectshadwatchedthemovie.
472BrainTopogr(2015)28:471–478123DataAcquisitionandAnalysisAuditoryevokedeldswererecordedwithahelmet-shaped,306-channelsMEGsystem(Vector-view,ELEKTA,Neu-romag,Helsinki,Finland)with102identicaltriplesensorelementslocatedinasilent,magneticallyshieldedroom.
WeanalyzedtheMEGsignalsrecordedby204planar-typegradiometers,detectingthelargestsignalsoverthecorre-spondingcerebralsources.
Signalswerepassedthrougha0.
03–200Hzband-passlteranddigitizedat600Hz.
ThemagneticeldsevokedbyTSwereselectivelyaveragedforeachcondition(standard,SD,andTD)includingpre-andpost-stimulusintervals(-100to600ms).
Inthepresentstudy,TSonset(latency=0ms)wasdenedwhentherstclickoftheTSreachedtheeardrumsimulatedbyanarti-cialear(Type4157,Bru¨el&KjrSoundandVibrationMeasurement,Nrum,Denmark).
Subjectswereinstructednottomovetheirheadsduringtherecordingsandtheircompliancewasmonitoredthroughavideocamerabytheexperimenter.
Inordertoimprovethesignal-to-noiseratiooftheauditoryevokedmagneticresponses,epochscontainingamplitudechangesgreaterthan2.
7pTwithinthepre-andpost-stimulusintervals(-100to600ms)wereautomati-callydiscardedasartifact-contaminatedepochs.
Afterarti-factrejection,epochswereaveragedforeachcondition(standard,SD,andTD),regardlessofthesoundtypes(TS30_Low,TS30_High,TS60_Low,andTS60_High).
ToanalyzetheMMNmcomponent,whichiselicitedbydeviantauditorysignals(Na¨a¨ta¨nenetal.
2007;Alho1995),theaveragedauditoryevokedeldsineachcondition(SD,TD,andstandardstimuli)were1–30Hzband-passlteredinordertoextractthetransientevokedresponses,andthebaselinewascorrectedrelativetothe100mspre-stimulusinterval.
Thereafter,inordertoobtaintheMMNmwave-formselicitedbySD(MMNm_SD)andTD(MMNm_TD),theauditoryevokedeldselicitedbythestandardTSweresubtractedfromthoseelicitedbySDandTD.
TheonsetofSDmatchedwiththerstclickoftheTS(latency=0ms),whereasTDdidnotoccurattherstclickoftheTS.
When30Hzclicktrains(TS30_LoworTS30_High)wereusedasthestandardTS,thetemporaldeviantoccurredatthepre-sentationofthesecondclickofTS60_LoworTS60_High(latency=16.
7ms).
When60Hzclicktrains(TS60_LoworTS60_High)wereusedasthestandardTS,theonsetofTDcouldbethetimingofthemissingsecondclickofthestandardstimuli(latency=16.
7ms).
Therefore,afterobtainingthesubtractedmagneticwaveforms(MMNm_SDandMMNm_TD)thelatencyofMMNm_TDwasoffsetbyareductionin16.
7msandwasthenusedforthesubsequentstatisticalanalysis.
Inordertoinvestigatedifferencesinthemagneticsen-sors,thetimecoursesoftheroot-mean-square(RMS)amplitudesofthesubtractedmagneticelds(MMNm_SDorMMNm_TD)werecalculatedbyusingalloftheleft-side(96sensors)orright-side(96sensors)planar-typegradiometersineachsubject.
ThemostprominentRMSpeakineachhemisphererangingfrom100to250msafterthesoundonsetwasdenedastheMMNmresponseineachsubject.
ThemeanRMSvaluewithinthe10mstimewindowaroundtheRMSpeakineachcondition,eachside,andeachsubjectwasusedinstatisticalanalysis.
ThemeanRMSamplitudesandlatenciesoftheMMNmresponseswereevaluatedseparatelybymeansofrepeated-measuresanalysesofvariance(ANOVA)usingthetwofactorsDEVIANT_CONDITION[spectraldeviant(SD)vs.
tem-poraldeviant(TD)]andHEMISPHERE(leftvs.
right).
Theestimatedsingledipolesourcestrengthwasshowntobemodulatedeasilybythedepthoftheestimatedlocation(HillebrandandBarnes2002).
Wecouldobtainreliablesourcestrengthsusingidenticalsourcelocationsandorientationsbetweenconditions.
Inordertoimprovethesignal-to-noiseratio,weaveragedMMNm_SDandMMNm_TDineachsubjectandusedtheaveragedmag-neticwaveformstoestimatethesingleequivalentcurrentdipolesreectingtheMMNmresponse.
ThepeakMMNmresponsewasinitiallyidentiedasthemaximalRMSvalueoftheglobaleldpowerbetween100and250msafterTSonset.
A10msintervalaroundtheMMNmpeaklatencywasselected,andthesourcelocationsandorientationswereestimatedusingsingleequivalentcurrentdipolemodeling(onedipoleperhemisphere)foreachsubjectindividually(BESAResearch5.
3.
7,BESAGmbH,Ger-many).
Wecalculatedthetwoequivalentcurrentdipoles(onedipoleperhemisphere)simultaneouslybyusingallwhole-headplanar-typegradiometers(204channels)fortheMMNmsourceestimation.
Dipoleestimationwasnotsuccessfulinonesubject,whichreducedthenumberofsubjectstoN=12.
Thegoodness-of-tfortheMMNmdipolesoftheremaining12subjectswasmorethan80%(mean±standarddeviation:89.
0±3.
0).
Theestimated1.
3secTemporalDeviantSpectralDeviantStandardStandardStandardStandardStandardFig.
1Schematicdepictionofthesoundstimulation.
Standardteststimuli(70%)werepresentedtogetherwithspectraldeviants(SD:15%)andtemporaldeviants(TD:15%)withinanoddballparadigmBrainTopogr(2015)28:471–478473123sources,whichwerexedinlocationandorientationforeachhemisphereofeachsubject,servedasaspatiallter(Tescheetal.
1995)tocalculatethesourcestrengthforeachcondition(SDandTD)andineachhemisphere(leftandright)ofeachsubject.
Themeansourcestrengthwithinthe10mstimewindowaroundthepeakMMNmlatencywasusedforfurtherstatisticalanalysisoftheMMNm.
Inordertoevaluatetheeffectsofthedevianttypeandhemisphere,thesourcestrengthsandlatenciesoftheesti-matedequivalentcurrentdipolescorrespondingtotheMMNmresponseselicitedbythedeviantstimuli(SDandTD)ineachhemispherewereevaluatedseparatelyviaarepeated-measuresANOVAusingthetwofactorsDEVI-ANT_CONDITION(SDvs.
TD)andHEMISPHERE(Leftvs.
Right).
ResultsTwelvesubjects(exceptforoneexcludedsubject)underwentanadequatenumberoftrialstoobtainauditoryevokedeldsforeachconditionaftertheartifactrejection[mean±stan-darddeviation:SD=332.
7±3.
6(99.
0±1.
1%),TD=333.
8±1.
7(99.
3±0.
5%),standardstimuli=1556.
2±9.
3(99.
2±0.
6%)].
Anexampleofindividualmagneticeldwaveformsineachcondition(SD,TD,andstandard)andsubtractedwaveforms[MMNm_SD(SD–standard),MMNm_TD(TD–standard)]isshowninFig.
2,whichdemonstratestheclearN1m-responseselicitedbyTSonsetintheupperpanelsaswellasMMNm-responsesinthesub-tractedwaveformsinthelowerpanels.
ThecalculatedmeansoftheRMSvaluesoftheauditoryevokedeldsforeachcondition(MMNm_SDandMMNm_TD)ineachhemisphereaveragedacross12subjectsaredisplayedinFig.
3,inwhichtheRMSwave-formselicitedbyTDwereshifted16.
7mstotheleft-sideinordertoadjustthetimingofthedeviantsoundonset.
ClearMMNmresponseswereobservedinbothconditionsandhemispheres.
TheRMSpeaksintheMMNm_TDconditionwerelaterthanthoseintheMMNm_SDcondi-tioninbothhemispheres.
ThemeanRMSamplitudesandlatenciesoftheMMNmresponsesaveragedacross12subjectsforeachconditionineachhemispherearepresentedinFig.
4witherrorbarsdenotingthe95%condenceintervalscalculatedbythemeansofbootstrapresamplingtests(iteration=100,000).
Therepeated-measuresANOVAappliedtothemaximalRMSamplitudesoftheMMNmresponsesineachhemi-sphereresultedinasignicantmaineffectforDEVI-ANT_CONDITION(F(1,11)=11.
78,p\0.
01),butnotforHEMISPHERE(F(1,11)=2.
53,p=0.
14).
Additionally,amarginaltrendtowardsignicancewasobservedintheinteractionbetweenDEVIANT_CONDITIONandHEMI-SPHERE[F(1,11)=4.
54,p=0.
056].
Therepeated-mea-suresANOVAappliedtothelatenciesofthemaximalRMSamplitudesoftheMMNmresponsesresultedinaFig.
2Examplesofindividualmagneticwaveforms.
Theupperpanelsrepresenttheauditoryevokedeldsofonerepresentativesubjectelicitedbyaspectraldeviant(SD),bstandard,andctemporaldeviant(TD)soundstimuli.
Thelowerpanelsshowthemagneticwaveformsobtainedbythesubtractionbetweenaandb[dspectralmismatchnegativity(MMNm_SD)]andbetweencandb[etemporalmismatchnegativity(MMNm_TD)]474BrainTopogr(2015)28:471–478123signicantmaineffectforDEVIANT_CONDITION[F(1,11)=36.
30,p\0.
001],butneitherasignicantmaineffectnorasignicantinteractionwereobserved[HEMI-SPHERE[F(1,11)=0.
33,p=0.
58];DEVIANT_CONDI-TION9HEMISPHERE[F(1,11)=0.
78,p=0.
40].
ThecalculatedmeansoftheMMNmsourcestrengthwaveformsforeachhemisphereaveragedacross12subjectsaredisplayedinFig.
3,inwhichMMNmsourcestrengthwaveformselicitedbyTDwereshifted16.
7mstotheleft-side.
ClearMMNm-responsesrangingbetween100and200mswereobservedinbothhemispheresafterTSonset.
ThemeanMMNmsourcestrengthsandlatenciesaveragedacross12subjectsforeachconditionineachhemispherearepresentedinFig.
4witherrorbarsdenotingthe95%con-denceintervalscalculatedbymeansofbootstrapresam-plingtests(iteration=100,000).
Therepeated-measuresANOVAappliedtotheMMNmsourcestrengthsrevealedasignicantmaineffectforDEVIANT_CONDITION[F(1,11)=6.
44,p\0.
03].
Additionally,asignicantinter-actionwasobservedbetweenDEVIANT_CONDITIONandFig.
3Grand-averaged(N=12)root-mean-square(RMS)valuesofthemagneticelds(leftpanel)andgrand-averagedsourcestrengths(rightpanel)ofthemismatchnegativity(MMNm)waveforms.
Solidanddashedlinesrepresentthespectraldeviant(MMNm_SD)andthetemporaldeviant(MMNm_TD)conditions,respectively.
Graylinesrepresenttheleftsensor(leftpanel)andlefthemisphere(rightpanel)andblacklinesrepresenttherightsensor(leftpanel)andrighthemisphere(rightpanel)Fig.
4Theleftandrightgraphsdisplaythemeanroot-mean-square(RMS)valuesandlatenciesofthemagneticeldscorrespondingtothemismatchnegativity(MMNm)andmeanMMNmsourcestrengthsandlatencieswitherrorbarsdenoting95%condenceintervals,respectively.
Filledbarsdenotetheleftsensor(LS:leftpanels)andlefthemisphere(LH:rightpanels)responsesandopenbarsdenotetherightsensor(RS:leftpanels)orrighthemisphere(RH:rightpanels)responsesBrainTopogr(2015)28:471–478475123HEMISPHERE[F(1,11)=6.
67,p\0.
03],whichindicatedthattheMMNmresponseelicitedbySDwasrelativelylargerintherighthemisphere,whereastheMMNmresponseelicitedbyTDwasrelativelylargerinthelefthemisphere.
Therepeated-measuresANOVAappliedtotheMMNmlatenciesrevealedasignicantmaineffectforDEVI-ANT_CONDITION[F(1,11)=48.
45,p\0.
001],butnosignicantinteractionbetweenfactors:MMNm_TDwassignicantlylongerthanthatofMMNm_SD.
WealsoanalyzedMMNmsourcestrengthsandlatencieswhentheMMNm_TDwasnotshiftedby16.
7msduringthecalculation.
Arepeated-measuresANOVAperformedontheMMNmsourcestrengthsrevealedasignicantmaineffectforDEVIANT_CONDITION[F(1,11)=6.
29,p\0.
03]andasignicantinteractionbetweenDEVIANT_CONDITIONandHEMISPHERE[F(1,11)=6.
87,p\0.
03].
Arepeated-measuresANOVAperformedontheMMNmlatenciesrevealedasignicantmaineffectforDEVIANT_CONDI-TION[F(1,11)=125.
3,p\0.
001],butnosignicantinter-actionbetweenfactors.
DiscussionTheresultsobtainedinthepresentstudyclearlydemon-stratedadifferenceinthehemisphericlateralityofMMNmamplitudesbetweenSDandTDconditions.
TheamplitudesofMMNmevokedbySD(MMNm_SD)wererelativelylargerintheright,whereasthoseevokedbyTD(MMNm_TD)wererelativelylargerintheleft(Figs.
3,4).
NohemisphericdifferencewasobservedintheMMNmlatency;however,thelatenciesofMMNm_TDweresig-nicantlylongerthanthoseofMMNm_SDinboththeleftandrighthemispheresevenwhentheonsettimedifferencebetweenSDandTD(16.
7ms)wasconsidered.
Incontrasttopreviousstudies(Alhoetal.
1998;Shtyrovetal.
2000),whichalsoinvestigatedthehemisphericasymmetryofMMN(m),thetotalsoundinputswereidenticalbetweenSDandTDconditionsinthepresentstudy.
Therefore,thesoundpropertyitselfcannotexplaintheobtainedresults;thedeviationpattern(SDorTD)fromthestandardsoundstreamwassolelyresponsiblefortheresultsobtained.
Weusedband-passlteredclick-trainsthatdidnotconveyspecicmeaningstoensurethathemisphericlateralizationforpre-attentivehumanauditoryprocessing,representedbyMMN(m),wasnotlimitedtothecomplexwaveformsfromnaturalsoundsources(e.
g.
humanvoiceormusicalinstruments),butinpartoriginatedfromearly,low-levelauditoryneuralprocessingdealingwithbasicsoundchar-acteristics,namely,spectralandtemporalfeatures(ZatorreandBelin2001;Tallaletal.
1993;Poeppel2003;Boemioetal.
2005).
Itseemsplausiblethatspectralandtemporalsoundinformationisdifferentiallyencodedintoneuralactivity(BendorandWang2007;Sakaietal.
2009).
Spectralinformationisencodedintothemaximalmovementposi-tionofthebasilarmembraneinthecochlea.
Therefore,incaseoftheSDcondition,thegroupsofinnerhaircellscorrespondingtoSDsoundsweredifferentfromthosecorrespondingtostandardsounds.
Incontrast,TDsoundshadsimilarfrequencycharacteristicstostandardsoundsignals.
Similargroupsofinnerhaircellsonthetonotopicmapinthecochleaareactivated.
InordertodetecttheTDsoundsignal,thecentralauditorysystemshouldanalyzethetemporalpatternsofneuralactivity.
ThepresentresultsdemonstratedthattheMMNmlatencieselicitedbyTDweresignicantlylongerthanthoseelicitedbySD(Figs.
3,4).
First,wehavetoconsiderthetimingoftheSDandTDonsets.
Theoretically,SDisdetectablefromtherstclickoftheTSinthecochlea,whereasTDdetectionrequiresthesecondclickofthe60Hzband-passlteredclicktrainsdeviatedfromthestandard30Hzband-passlteredclicksorthemissingsecondclickofthestandard60Hzband-passlteredclicktrainsduringpresentationofthedeviant30Hzband-passlteredclicktrainstomanifestinthecentralauditorysystem.
Therefore,werstsubtracted16.
7msfromtheMMNm_TDlatencyinordertocompareitwiththeMMNm_SDlatency.
Evenafterthisadjustment,MMNm_TDwassignicantlylongerthanMMNm_SD(Fig.
4),whichsuggestedthatdifferentneuralmechanismscontributetothedetectionofspectralandtemporalsounddeviants.
Neuralencodingofthetemporalpatternsofauditorysignalstooklongerandappearedtotakeplaceatahigherleveloftheauditorysystemthanspectralcoding.
PreviousMEGstudies(Okamotoetal.
2009,2012)alsosupportthishypothesisbydemonstratingthatthetemporalchangeselicitedsignicantlydelayedauditoryN1mresponses,withamajordeectionintheauditoryevokedresponsehavingalatencyofapproximately100ms(Na¨a¨ta¨nenandPicton1987),thanthoseelicitedbyspectralchanges.
AuditoryMMNmisapre-attentiveautomaticbrainresponseelicitedbyanychangeinauditorystimulation(Na¨a¨ta¨nenetal.
2007).
Inthepresentstudy,weusedband-passlteredclicktrainsthatdidnotconveyspecicmeaningandsubjectsweredistractedfromtheauditorymodality;therefore,itislesslikelythatsubjectsinvolun-tarilyprocessedandperceivedthetestsoundsasmusicalorspeechsignals.
Theobtainedresultsindicatedthatthehemisphericasymmetryofauditoryprocessinginhumansstartsfromthebasic,pre-attentiveauditoryprocessinglevel.
Moreover,soundinputswerecompletelycounter-balancedbetweentheSDandTDconditions.
Therefore,thehemisphericasymmetryoftheMMNmresponseselicitedbytheSDandTDcouldnotbeexplainedsolelyby476BrainTopogr(2015)28:471–478123stimulusfeatures.
Thelateralizedmemorytracesofbasicauditoryprocessesintermsofspectralandtemporalsoundfeaturesappeartoberesponsiblefortheresultsobtained.
Recenthumanneuroimagingstudiesrevealedthatthefunctionalhemisphericasymmetryofauditoryprocessingwasnotlimitedtocomplexsoundsignalsconveyingspe-cicmeaningandrules(e.
g.
musicandspeech),butorig-inatedfromthebasicauditoryprocessinglevel,namely,thetemporalintegrationwindow(Poeppel2003;Belinetal.
1998;ZatorreandBelin2001;Zatorreetal.
2002).
Itisimportanttoquicklyandpreciselyencodeenvironmentalsoundsindailylife.
However,becauseofthetrade-offbetweentemporalandspectralanalysisprecision[Acousticuncertaintyprinciple;(Joos1948;Zatorreetal.
2002)],itisimpossibletoachievehighspectralandhightemporalsoundanalysesatthesametimeusingonetemporalinte-grationwindow.
Ashorttemporalintegrationwindowleadstohightemporalresolution,butrelativelylowspec-tralresolutionofthesoundanalyses.
Ontheotherhand,alongtemporalwindowleadstohighspectralresolution,butrelativelylowtemporalresolutionofthesoundanalyses.
Therefore,itseemsplausiblethatthehumanauditorycorticesintheleftandrighthemispheresadoptdifferentintegrationtimewindowsinsteadofapplyingonespecictemporalintegrationtimewindowinbothhemispheres.
Belinetal.
(1998)andPoeppel(2003)hypothesizedthatthelefthemisphereappliedashortertemporalintegrationwindow,resultinginabettertemporalresolutioncapabil-ity,andtherighthemisphereappliedalongertemporalintegrationwindow,resultinginabetterspectralresolutioncapability.
Inthepresentstudy,thelongertemporalinte-grationwindowwithhigherspectralresolutionintherighthemisphereappearstohavedominantlycontributedtodetectingspectrallydeviatedsoundsignalsandresultedinrelativelylargerMMNm_SDamplitudesintherighthemisphere.
Incontrast,theshortertemporalintegrationwindowwithhightemporalresolutioninthelefthemi-sphereappearstohavedominantlyprocessedtemporallydeviatedsoundsignalsandresultedinrelativelylargerMMNm_TDamplitudesinthelefthemisphere.
TheMMNmamplitudesandlatenciesobtainedinthesen-sorspaceandsourcespaceexhibitedsimilarpatterns(Figs.
3,4):theMMNm_SDandMMNm_TDamplitudeswerelargerintherightandlefthemispheres,respectively.
However,theANOVAexaminingMMNmamplitudesresultedinasigni-cantinteractionbetweenDEVIANT_CONDITIONandHEMISPHEREinthesourcespacedata[F(1,11)=6.
67,p\0.
03],butonlyamarginaltrendtowardsignicancewasobservedinthesensorspacedata[F(1,11)=4.
54,p=0.
056].
Themainreasonforthisinconsistencymaybethattheneuralsourcesinonehemispherecouldinuencetheevokedmag-neticeldsinthecontra-lateralmagneticsensors.
Moreover,headsizesandheadpositionsdifferedbetweensubjectsandthecentralsulcusofthesubjectscouldshiftfromthecenteroftheMEGdewar.
Therefore,thesefactorsmayhaveledtoalessrobuststatisticaloutcomeintheRMSamplitudesoftheMMNmresponsesthantheMMNmsourcestrengths.
Inconclusion,usingcarefullyconstructedauditorystimulithatwerecounter-balancedbetweenconditionsandhadcleartime-lockedonsetsofSDandTD,thepresentstudyclearlydemonstratedthatneuralprocessingdealingwithspectrallydeviatedsoundswererelativelydominantintherighthemispherewhilethosedealingwithtemporallydeviatedsoundswererelativelydominantinthelefthemisphere.
Theseresultsstronglysupportthehypothesisthatthehumanbrainadoptsasymmetricmemorytracesofbasicspectralandtemporalsoundfeaturesintheleftandrighthemispheresinordertoimprovethedetectionofdeviantsoundsignals.
AcknowledgmentsWethankY.
Takeshimafortechnicalhelpandourtestsubjectsfortheirdiligentcollaboration.
Thisstudywassupportedbythe''JapanSocietyforthePromotionofScienceforYoungScientists(23689070)'',''StrategicResearchProgramforBrainSciences(Developmentofbiomarkercandidatesforsocialbehavior)'',and''SoundTechnologyPromotionFoundation''.
ConictofinterestTheauthorshavedeclaredthatnocompetinginterestsexist.
OpenAccessThisarticleisdistributedunderthetermsoftheCreativeCommonsAttributionLicensewhichpermitsanyuse,dis-tribution,andreproductioninanymedium,providedtheoriginalauthor(s)andthesourcearecredited.
ReferencesAlhoK(1995)Cerebralgeneratorsofmismatchnegativity(MMN)anditsmagneticcounterpart(MMNm)elicitedbysoundchanges.
EarHear16:38–51AlhoK,ConnollyJF,CheourM,LehtokoskiA,HuotilainenM,VirtanenJ,AulankoR,IlmoniemiRJ(1998)Hemisphericlateralizationinpreattentiveprocessingofspeechsounds.
NeurosciLett258:9–12BelinP,ZilboviciusM,CrozierS,ThivardL,FontaineA(1998)Lateralizationofspeechandauditorytemporalprocessing.
JCognNeurosci10:536–540BelinP,ZatorreRJ,LafailleP,AhadP,PikeB(2000)Voice-selectiveareasinhumanauditorycortex.
Nature403:309–312BendorD,WangXQ(2007)Differentialneuralcodingofacousticutterwithinprimateauditorycortex.
NatNeurosci10:763–771BoemioA,FrommS,BraunA,PoeppelD(2005)Hierarchicalandasymmetrictemporalsensitivityinhumanauditorycortices.
NatNeurosci8:389–395BrocaP(1861)Remarquessurlesiegedelafacultedulangagearticulesuiviesd'uneobservationd'aphemie(pertedelaparole).
BullSocAnat6:330–357DrullmanR,FestenJM,PlompR(1994a)Effectofreducingslowtemporalmodulationsonspeechreception.
JAcoustSocAm95:2670–2680DrullmanR,FestenJM,PlompR(1994b)Effectoftemporalenvelopesmearingonspeechreception.
JAcoustSocAm95:1053–1064BrainTopogr(2015)28:471–478477123EulitzC,DieschE,PantevC,HampsonS,ElbertT(1995)Magneticandelectricbrainactivityevokedbytheprocessingoftoneandvowelstimuli.
JNeurosci15:2748–2755GrifthsTD,JohnsrudeI,DeanJL,GreenGG(1999)AcommonneuralsubstratefortheanalysisofpitchanddurationpatterninsegmentedsoundNeuroreport10:3825–3830HillebrandA,BarnesGR(2002)Aquantitativeassessmentofthesensitivityofwhole-headMEGtoactivityintheadulthumancortex.
Neuroimage16:638–650JamisonHL,WatkinsKE,BishopDV,MatthewsPM(2006)Hemisphericspecializationforprocessingauditorynonspeechstimuli.
CerebCortex16:1266–1275JoosM(1948)Acousticphonetics.
Languagemonograph,vol23.
LinguisticSocietyofAmerica,BaltimoreKujalaT,TervaniemiM,SchrogerE(2007)Themismatchnegativityincognitiveandclinicalneuroscience:theoreticalandmethod-ologicalconsiderations.
BiolPsychol74:1–19LappeC,Steinstra¨terO,PantevC(2013)AbeamformeranalysisofMEGdatarevealsfrontalgeneratorsofthemusicallyelicitedmismatchnegativity.
PLoSOne8:e61296MooreBCJ(2003)Anintroductiontothepsychologyofhearing.
AcademicPress,BostonNa¨a¨ta¨nenR,PictonT(1987)TheN1waveofthehumanelectricandmagneticresponsetosound:areviewandananalysisofthecomponentstructure.
Psychophysiology24:375–425Na¨a¨ta¨nenR,GaillardAW,Ma¨ntysaloS(1978)Earlyselective-attentioneffectonevokedpotentialreinterpreted.
ActaPsychol42:313–329Na¨a¨ta¨nenR,PaavilainenP,RinneT,AlhoK(2007)Themismatchnegativity(MMN)inbasicresearchofcentralauditoryprocess-ing:areview.
ClinNeurophysiol118:2544–2590OkamotoH,StrackeH,DraganovaR,PantevC(2009)Hemisphericasymmetryofauditoryevokedeldselicitedbyspectralversustemporalstimuluschange.
CerebCortex19:2290–2297OkamotoH,TeismannH,KakigiR,PantevC(2012)Auditoryevokedeldselicitedbyspectral,temporal,andspectral-temporalchangesinhumancerebralcortex.
FrontPsychol3:149OldeldRC(1971)Theassessmentandanalysisofhandedness:theEdinburghinventory.
Neuropsychologia9:97–113PoeppelD(2003)Theanalysisofspeechindifferenttemporalintegrationwindows:cerebrallateralizationas'asymmetricsamplingintime'.
SpeechCommun41:245–255SakaiM,ChimotoS,QinL,SatoY(2009)Differentialrepresentationofspectralandtemporalinformationbyprimaryauditorycortexneuronsinawakecats:relevancetoauditorysceneanalysis.
BrainResCognBrainRes1265:80–92ShannonRV,ZengFG,KamathV,WygonskiJ,EkelidM(1995)Speechrecognitionwithprimarilytemporalcues.
Science270:303–304ShtyrovY,KujalaT,PalvaS,IlmoniemiRJ,NaatanenR(2000)Discriminationofspeechandofcomplexnonspeechsoundsofdifferenttemporalstructureintheleftandrightcerebralhemispheres.
Neuroimage12:657–663SzymanskiMD,PerryDW,GageNM,RowleyHA,WalkerJ,BergerMS,RobertsTP(2001)Magneticsourceimagingoflateevokedeldresponsestovowels:towardanassessmentofhemisphericdominanceforlanguage.
JNeurosurg94:445–453TallalP,MillerS,FitchRH(1993)Neurobiologicalbasisofspeech:acaseforthepreeminenceoftemporalprocessing.
AnnNYAcadSci682:27–47TervaniemiM,KujalaA,AlhoK,VirtanenJ,IlmoniemiRJ,Na¨a¨ta¨nenR(1999)Functionalspecializationofthehumanauditorycortexinprocessingphoneticandmusicalsounds:amagnetoencephalographic(MEG)study.
Neuroimage9:330–336TescheCD,UusitaloMA,IlmoniemiRJ,HuotilainenM,KajolaM,SalonenO(1995)Signal-spaceprojectionsofmegdatacharac-terizebothdistributedandwell-localizedneuronalsources.
ElectroencephalogrClinNeurophysiolSuppl95:189–200VosPG,TroostJM(1989)Ascendinganddescendingmelodicintervals—statisticalndingsandtheirperceptualrelevance.
MusicPercept6:383–396WarrierCM,ZatorreRJ(2002)Inuenceoftonalcontextandtimbralvariationonperceptionofpitch.
PerceptPsychophys64:198–207WernickeC(1874)Symptomenkomplex.
EinepsychologischeStudieaufanatomischerBasis.
CohnundWeigert,BreslauZatorreRJ,BelinP(2001)Spectralandtemporalprocessinginhumanauditorycortex.
CerebCortex11:946–953ZatorreRJ,EvansAC,MeyerE(1994)Neuralmechanismsunderlyingmelodicperceptionandmemoryforpitch.
JNeurosci14:1908–1919ZatorreRJ,BelinP,PenhuneVB(2002)Structureandfunctionofauditorycortex:musicandspeech.
TrendsCognSci6:37–46478BrainTopogr(2015)28:471–478123
ThisarticleispublishedwithopenaccessatSpringerlink.
comAbstractOneofthemajorchallengesinhumanbrainscienceisthefunctionalhemisphericasymmetryofaudi-toryprocessing.
Behavioralandneurophysiologicalstudieshavedemonstratedthatspeechprocessingisdominantlyhandledinthelefthemisphere,whereasmusicprocessingdominantlyoccursintheright.
Usingmagnetoencepha-lography,wemeasuredtheauditorymismatchnegativityelicitedbyband-passlteredclick-trains,whichdeviatedfromfrequentlypresentedstandardsoundsignalsinaspectralortemporaldomain.
Theresultsshowedthatspectralandtemporaldeviantsweredominantlyprocessedintherightandlefthemispheres,respectively.
Hemisphericasymmetrywasnotlimitedtohigh-levelcognitivepro-cesses,butalsooriginatedfromthepre-attentiveneuralprocessingstagerepresentedbymismatchnegativity.
KeywordsAuditoryevokedresponseHemisphericlateralityMagnetoencephalography(MEG)Mismatchnegativity(MMNm)AbbreviationsANOVAAnalysisofvarianceMEGMagnetoencephalographyMMNMismatchnegativitySDSpectraldeviantTDTemporaldeviantTSTeststimulusIntroductionFunctionalhemisphericasymmetryinthehumanbrainhasbeeninvestigatedsincethelatenineteenthcentury(Wer-nicke1874;Broca1861).
Inadditiontotheclassicalbehavioralobservationsofneurologicaldisorderpatients,recentneuroimagingtechniqueshavemadeitpossibletoinvestigateconscioushealthyhumanbrains,andhaverevealedlefthemisphericdominanceforspeechprocessingandrighthemisphericdominanceformusicprocessing(Zatorreetal.
1994,2002;Grifthsetal.
1999;Belinetal.
2000;Eulitzetal.
1995;Szymanskietal.
2001;Alhoetal.
1998).
However,functionalhemisphericasymmetryinthehumanbrainmaynotbelimitedtohigh-levelcognitiveneuralprocesses,butmaystartfromthelowerneuralprocessinglevelofbasicacousticfeatures(e.
g.
frequency,interval,duration,andintensity).
Naturalsoundshavespecicspectraldistributionsthatchangeovertimeaccordingtospecictemporalsequences.
Bothspectralandtemporalsoundfeatureshavebeenshowntoplayanimportantroleintheperceptionofnaturalsounds(Moore2003);however,theimportanceofthesefeaturesappearstodifferbetweensoundtypes,withspectralprocessingbeingofparticularimportanceformusicperception(VosandTroost1989;WarrierandZa-torre2002)andtemporalcuesbeingessentialforspeechperception(Shannonetal.
1995;Drullmanetal.
1994a,b).
Recentfunctionalmagneticresonanceimaging(Jamisonetal.
2006),positronemissiontomography(ZatorreandBelin2001),andmagnetoencephalography(MEG)ThisisoneofseveralpaperspublishedtogetherinBrainTopographyonthe''SpecialIssue:AuditoryCortex2012''.
ElectronicsupplementarymaterialTheonlineversionofthisarticle(doi:10.
1007/s10548-013-0347-1)containssupplementarymaterial,whichisavailabletoauthorizedusers.
H.
Okamoto(&)R.
KakigiDepartmentofIntegrativePhysiology,NationalInstituteforPhysiologicalSciences,38Nishigo-Naka,Myodaiji,Okazaki444-8585,Japane-mail:hokamoto@nips.
ac.
jp123BrainTopogr(2015)28:471–478DOI10.
1007/s10548-013-0347-1(Okamotoetal.
2009)studieshavedemonstratedusingarticialbasicauditorystimuli(e.
g.
puretonesandpulse-trains)thattemporalchangesaredominantlyprocessedinthelefthemisphere,whereasspectralchangesaredomi-nantlyprocessedintheright.
Thewell-knownfunctionalhumanhemisphericasymmetryobservedforspeechandmusicprocessingmaynotbelimitedtoconscioushigh-levelcognitiveprocesses,butmaybeatleastpartiallyrelatedtothepre-attentiveprocessingoflow-levelacousticfeatures.
Mismatchnegativity(MMN)anditsmagneticcounter-partMMNmareauditoryevokedcomponentsthatreectthecorticalpre-attentivediscriminationofauditorystimuliaswellasauditorymemorytraces(Na¨a¨ta¨nenetal.
1978,2007;Kujalaetal.
2007).
MMN(m)iselicitedbyviola-tionsofregularitiesinsoundstreamsandcanberecordedwithoutanymotororotherresponseandcanevenbeobtainedfrominattentivepatientsandinfants.
PreviousstudieshaveshownthatMMN(m)elicitedbyspeechsoundswassignicantlylateralizedtothelefthemisphere(Alhoetal.
1998),whereasMMN(m)elicitedbymusicalnoteswasdominantlyprocessedintherighthemisphere(Lappeetal.
2013;Tervaniemietal.
1999).
However,whetherthehemisphericasymmetriesofthepre-attentiveMMN(m)arelimitedtomeaningfulauditorystimuli(e.
g.
speechandmusic)ororiginatefromthebasicspectralandtemporalsoundfeaturesofthesesoundstimuliremainsunknown.
Therefore,theaimofthepresentstudywastoinvesti-gatethehemisphericlateralityofMMNmevokedbyspectralversustemporalsounddeviantsthatdonotconveyspecicphonological,grammatical,ormusicalinforma-tion.
InordertoexcludethepossibilitythatthelateralityoftheMMNmoriginatedfromthesoundstimulusitself,wecounter-balancedtotalauditoryinputsidenticalbetweenspectralandtemporaldeviantconditions.
Theresultsofthepresentstudyprovideanewinsightintohowtheleftandrighthemispherespre-attentivelydealwiththespectralandtemporalfeaturesofnaturalsoundsignals.
MaterialsandMethodsSubjectsThirteenhealthysubjectsparticipatedinthisstudy(vefemales;mean±standarddeviation:32.
1±6.
2years).
Allparticipantshadnormalhearing,hadnohistoryofpsychologicalorneurologicaldisorders,andwereunam-biguouslyright-handed[assessedviatheJapaneseversionof''EdinburghHandednessInventory''(Oldeld1971)].
AllparticipantswerefullyinformedaboutthestudyandgavewritteninformedconsentfortheirparticipationinaccordancewiththeproceduresapprovedbytheEthicsCommissionoftheNationalInstituteforPhysiologicalSciences,Okazaki,Japan.
ThestudyconformedtotheCodeofEthicsoftheWorldMedicalAssociation(Decla-rationofHelsinki).
StimuliandExperimentalDesignTheexperimentaldesignisschematicallyrepresentedinFig.
1.
Theteststimulus(TS)waseithera30Hz(TS30)or60Hz(TS60)click-train,whichwasone-octaveband-passlteredeitherbetween500and1,000Hz(TS30_Low(SupplementaryAudio1S)andTS60_Low(SupplementaryAudio2S))orbetween1,000and2,000Hz[TS30_High(SupplementaryAudio3S)andTS60_High(SupplementaryAudio4S)].
TheTShadadurationof330msandthesoundonsetasynchronybetweentheTSwas1,300ms.
OneoftheTSwerepresentedasstandardstimuliwith70%probabilitypseudo-randomlyintermixedwithspectraldeviants(SD:15%probability)andtemporaldeviants(TD:15%proba-bility)inanoddballsequenceasdemonstratedinFig.
1.
IncaseofSD,band-passltersettingschangedfromthestandardstimulus,whilethetypeoftheclick-trainremainedidentical(standardsandSD:TS30_LowandTS30_High,TS30_HighandTS30_Low,TS60_LowandTS60_High,TS60_HighandTS60_Low).
Ontheotherhand,incaseofTD,theltersettingsremainedidentical,whilethetypeofclick-trainchangedfromthestandardsoundstimulus(standardsandTD:TS30_LowandTS60_Low,TS30_HighandTS60_High,TS60_LowandTS30_Low,TS60_HighandTS30_High).
Morethantwostandardstimuliwerepresentedbeforeadeviantstimulus(SDorTD).
EachMEGsessionconsistedoffourblocks.
Eachblockcontainedfoursub-blocksthatpseudo-randomlyadoptedTS30_Low,TS30_High,TS60_Low,andTS60_HighasthestandardTS,respectively.
Consequently,allTStypeswerepresentedwithaprobabilityof25%inoneblock.
Eachsub-blockhad21SD,21TD,and98standardstimuli,resultinginatotalnumberof336trialsforeachdeviantstimulusand1,568trialsforthestandardcondition.
Allsoundsweredioticallypresentedthroughplastictubes1.
5minlengthandear-piecesttedtothesubject'sears.
BeforestartinganMEGmeasurement,eachsubject'shearingthresholdforTS30_Lowwasindividuallydeterminedforeachear.
Dur-ingtheMEGrecordingsession,TS30_Lowwaspresentedatanintensityof60dBabovetheindividualsensationlevel,andotherTSwereadjustedtohavepoweridenticaltoTS30_Low.
Inordertokeepthetestsubjectsalertanddis-tractedfromtheauditorysignals,aself-chosensilentmoviewithcaptionswaspresentedduringtheMEGrecordings.
Questionsregardingthecontentofthemoviewereaskedattheendofthemeasurementtoensurethatthesubjectshadwatchedthemovie.
472BrainTopogr(2015)28:471–478123DataAcquisitionandAnalysisAuditoryevokedeldswererecordedwithahelmet-shaped,306-channelsMEGsystem(Vector-view,ELEKTA,Neu-romag,Helsinki,Finland)with102identicaltriplesensorelementslocatedinasilent,magneticallyshieldedroom.
WeanalyzedtheMEGsignalsrecordedby204planar-typegradiometers,detectingthelargestsignalsoverthecorre-spondingcerebralsources.
Signalswerepassedthrougha0.
03–200Hzband-passlteranddigitizedat600Hz.
ThemagneticeldsevokedbyTSwereselectivelyaveragedforeachcondition(standard,SD,andTD)includingpre-andpost-stimulusintervals(-100to600ms).
Inthepresentstudy,TSonset(latency=0ms)wasdenedwhentherstclickoftheTSreachedtheeardrumsimulatedbyanarti-cialear(Type4157,Bru¨el&KjrSoundandVibrationMeasurement,Nrum,Denmark).
Subjectswereinstructednottomovetheirheadsduringtherecordingsandtheircompliancewasmonitoredthroughavideocamerabytheexperimenter.
Inordertoimprovethesignal-to-noiseratiooftheauditoryevokedmagneticresponses,epochscontainingamplitudechangesgreaterthan2.
7pTwithinthepre-andpost-stimulusintervals(-100to600ms)wereautomati-callydiscardedasartifact-contaminatedepochs.
Afterarti-factrejection,epochswereaveragedforeachcondition(standard,SD,andTD),regardlessofthesoundtypes(TS30_Low,TS30_High,TS60_Low,andTS60_High).
ToanalyzetheMMNmcomponent,whichiselicitedbydeviantauditorysignals(Na¨a¨ta¨nenetal.
2007;Alho1995),theaveragedauditoryevokedeldsineachcondition(SD,TD,andstandardstimuli)were1–30Hzband-passlteredinordertoextractthetransientevokedresponses,andthebaselinewascorrectedrelativetothe100mspre-stimulusinterval.
Thereafter,inordertoobtaintheMMNmwave-formselicitedbySD(MMNm_SD)andTD(MMNm_TD),theauditoryevokedeldselicitedbythestandardTSweresubtractedfromthoseelicitedbySDandTD.
TheonsetofSDmatchedwiththerstclickoftheTS(latency=0ms),whereasTDdidnotoccurattherstclickoftheTS.
When30Hzclicktrains(TS30_LoworTS30_High)wereusedasthestandardTS,thetemporaldeviantoccurredatthepre-sentationofthesecondclickofTS60_LoworTS60_High(latency=16.
7ms).
When60Hzclicktrains(TS60_LoworTS60_High)wereusedasthestandardTS,theonsetofTDcouldbethetimingofthemissingsecondclickofthestandardstimuli(latency=16.
7ms).
Therefore,afterobtainingthesubtractedmagneticwaveforms(MMNm_SDandMMNm_TD)thelatencyofMMNm_TDwasoffsetbyareductionin16.
7msandwasthenusedforthesubsequentstatisticalanalysis.
Inordertoinvestigatedifferencesinthemagneticsen-sors,thetimecoursesoftheroot-mean-square(RMS)amplitudesofthesubtractedmagneticelds(MMNm_SDorMMNm_TD)werecalculatedbyusingalloftheleft-side(96sensors)orright-side(96sensors)planar-typegradiometersineachsubject.
ThemostprominentRMSpeakineachhemisphererangingfrom100to250msafterthesoundonsetwasdenedastheMMNmresponseineachsubject.
ThemeanRMSvaluewithinthe10mstimewindowaroundtheRMSpeakineachcondition,eachside,andeachsubjectwasusedinstatisticalanalysis.
ThemeanRMSamplitudesandlatenciesoftheMMNmresponseswereevaluatedseparatelybymeansofrepeated-measuresanalysesofvariance(ANOVA)usingthetwofactorsDEVIANT_CONDITION[spectraldeviant(SD)vs.
tem-poraldeviant(TD)]andHEMISPHERE(leftvs.
right).
Theestimatedsingledipolesourcestrengthwasshowntobemodulatedeasilybythedepthoftheestimatedlocation(HillebrandandBarnes2002).
Wecouldobtainreliablesourcestrengthsusingidenticalsourcelocationsandorientationsbetweenconditions.
Inordertoimprovethesignal-to-noiseratio,weaveragedMMNm_SDandMMNm_TDineachsubjectandusedtheaveragedmag-neticwaveformstoestimatethesingleequivalentcurrentdipolesreectingtheMMNmresponse.
ThepeakMMNmresponsewasinitiallyidentiedasthemaximalRMSvalueoftheglobaleldpowerbetween100and250msafterTSonset.
A10msintervalaroundtheMMNmpeaklatencywasselected,andthesourcelocationsandorientationswereestimatedusingsingleequivalentcurrentdipolemodeling(onedipoleperhemisphere)foreachsubjectindividually(BESAResearch5.
3.
7,BESAGmbH,Ger-many).
Wecalculatedthetwoequivalentcurrentdipoles(onedipoleperhemisphere)simultaneouslybyusingallwhole-headplanar-typegradiometers(204channels)fortheMMNmsourceestimation.
Dipoleestimationwasnotsuccessfulinonesubject,whichreducedthenumberofsubjectstoN=12.
Thegoodness-of-tfortheMMNmdipolesoftheremaining12subjectswasmorethan80%(mean±standarddeviation:89.
0±3.
0).
Theestimated1.
3secTemporalDeviantSpectralDeviantStandardStandardStandardStandardStandardFig.
1Schematicdepictionofthesoundstimulation.
Standardteststimuli(70%)werepresentedtogetherwithspectraldeviants(SD:15%)andtemporaldeviants(TD:15%)withinanoddballparadigmBrainTopogr(2015)28:471–478473123sources,whichwerexedinlocationandorientationforeachhemisphereofeachsubject,servedasaspatiallter(Tescheetal.
1995)tocalculatethesourcestrengthforeachcondition(SDandTD)andineachhemisphere(leftandright)ofeachsubject.
Themeansourcestrengthwithinthe10mstimewindowaroundthepeakMMNmlatencywasusedforfurtherstatisticalanalysisoftheMMNm.
Inordertoevaluatetheeffectsofthedevianttypeandhemisphere,thesourcestrengthsandlatenciesoftheesti-matedequivalentcurrentdipolescorrespondingtotheMMNmresponseselicitedbythedeviantstimuli(SDandTD)ineachhemispherewereevaluatedseparatelyviaarepeated-measuresANOVAusingthetwofactorsDEVI-ANT_CONDITION(SDvs.
TD)andHEMISPHERE(Leftvs.
Right).
ResultsTwelvesubjects(exceptforoneexcludedsubject)underwentanadequatenumberoftrialstoobtainauditoryevokedeldsforeachconditionaftertheartifactrejection[mean±stan-darddeviation:SD=332.
7±3.
6(99.
0±1.
1%),TD=333.
8±1.
7(99.
3±0.
5%),standardstimuli=1556.
2±9.
3(99.
2±0.
6%)].
Anexampleofindividualmagneticeldwaveformsineachcondition(SD,TD,andstandard)andsubtractedwaveforms[MMNm_SD(SD–standard),MMNm_TD(TD–standard)]isshowninFig.
2,whichdemonstratestheclearN1m-responseselicitedbyTSonsetintheupperpanelsaswellasMMNm-responsesinthesub-tractedwaveformsinthelowerpanels.
ThecalculatedmeansoftheRMSvaluesoftheauditoryevokedeldsforeachcondition(MMNm_SDandMMNm_TD)ineachhemisphereaveragedacross12subjectsaredisplayedinFig.
3,inwhichtheRMSwave-formselicitedbyTDwereshifted16.
7mstotheleft-sideinordertoadjustthetimingofthedeviantsoundonset.
ClearMMNmresponseswereobservedinbothconditionsandhemispheres.
TheRMSpeaksintheMMNm_TDconditionwerelaterthanthoseintheMMNm_SDcondi-tioninbothhemispheres.
ThemeanRMSamplitudesandlatenciesoftheMMNmresponsesaveragedacross12subjectsforeachconditionineachhemispherearepresentedinFig.
4witherrorbarsdenotingthe95%condenceintervalscalculatedbythemeansofbootstrapresamplingtests(iteration=100,000).
Therepeated-measuresANOVAappliedtothemaximalRMSamplitudesoftheMMNmresponsesineachhemi-sphereresultedinasignicantmaineffectforDEVI-ANT_CONDITION(F(1,11)=11.
78,p\0.
01),butnotforHEMISPHERE(F(1,11)=2.
53,p=0.
14).
Additionally,amarginaltrendtowardsignicancewasobservedintheinteractionbetweenDEVIANT_CONDITIONandHEMI-SPHERE[F(1,11)=4.
54,p=0.
056].
Therepeated-mea-suresANOVAappliedtothelatenciesofthemaximalRMSamplitudesoftheMMNmresponsesresultedinaFig.
2Examplesofindividualmagneticwaveforms.
Theupperpanelsrepresenttheauditoryevokedeldsofonerepresentativesubjectelicitedbyaspectraldeviant(SD),bstandard,andctemporaldeviant(TD)soundstimuli.
Thelowerpanelsshowthemagneticwaveformsobtainedbythesubtractionbetweenaandb[dspectralmismatchnegativity(MMNm_SD)]andbetweencandb[etemporalmismatchnegativity(MMNm_TD)]474BrainTopogr(2015)28:471–478123signicantmaineffectforDEVIANT_CONDITION[F(1,11)=36.
30,p\0.
001],butneitherasignicantmaineffectnorasignicantinteractionwereobserved[HEMI-SPHERE[F(1,11)=0.
33,p=0.
58];DEVIANT_CONDI-TION9HEMISPHERE[F(1,11)=0.
78,p=0.
40].
ThecalculatedmeansoftheMMNmsourcestrengthwaveformsforeachhemisphereaveragedacross12subjectsaredisplayedinFig.
3,inwhichMMNmsourcestrengthwaveformselicitedbyTDwereshifted16.
7mstotheleft-side.
ClearMMNm-responsesrangingbetween100and200mswereobservedinbothhemispheresafterTSonset.
ThemeanMMNmsourcestrengthsandlatenciesaveragedacross12subjectsforeachconditionineachhemispherearepresentedinFig.
4witherrorbarsdenotingthe95%con-denceintervalscalculatedbymeansofbootstrapresam-plingtests(iteration=100,000).
Therepeated-measuresANOVAappliedtotheMMNmsourcestrengthsrevealedasignicantmaineffectforDEVIANT_CONDITION[F(1,11)=6.
44,p\0.
03].
Additionally,asignicantinter-actionwasobservedbetweenDEVIANT_CONDITIONandFig.
3Grand-averaged(N=12)root-mean-square(RMS)valuesofthemagneticelds(leftpanel)andgrand-averagedsourcestrengths(rightpanel)ofthemismatchnegativity(MMNm)waveforms.
Solidanddashedlinesrepresentthespectraldeviant(MMNm_SD)andthetemporaldeviant(MMNm_TD)conditions,respectively.
Graylinesrepresenttheleftsensor(leftpanel)andlefthemisphere(rightpanel)andblacklinesrepresenttherightsensor(leftpanel)andrighthemisphere(rightpanel)Fig.
4Theleftandrightgraphsdisplaythemeanroot-mean-square(RMS)valuesandlatenciesofthemagneticeldscorrespondingtothemismatchnegativity(MMNm)andmeanMMNmsourcestrengthsandlatencieswitherrorbarsdenoting95%condenceintervals,respectively.
Filledbarsdenotetheleftsensor(LS:leftpanels)andlefthemisphere(LH:rightpanels)responsesandopenbarsdenotetherightsensor(RS:leftpanels)orrighthemisphere(RH:rightpanels)responsesBrainTopogr(2015)28:471–478475123HEMISPHERE[F(1,11)=6.
67,p\0.
03],whichindicatedthattheMMNmresponseelicitedbySDwasrelativelylargerintherighthemisphere,whereastheMMNmresponseelicitedbyTDwasrelativelylargerinthelefthemisphere.
Therepeated-measuresANOVAappliedtotheMMNmlatenciesrevealedasignicantmaineffectforDEVI-ANT_CONDITION[F(1,11)=48.
45,p\0.
001],butnosignicantinteractionbetweenfactors:MMNm_TDwassignicantlylongerthanthatofMMNm_SD.
WealsoanalyzedMMNmsourcestrengthsandlatencieswhentheMMNm_TDwasnotshiftedby16.
7msduringthecalculation.
Arepeated-measuresANOVAperformedontheMMNmsourcestrengthsrevealedasignicantmaineffectforDEVIANT_CONDITION[F(1,11)=6.
29,p\0.
03]andasignicantinteractionbetweenDEVIANT_CONDITIONandHEMISPHERE[F(1,11)=6.
87,p\0.
03].
Arepeated-measuresANOVAperformedontheMMNmlatenciesrevealedasignicantmaineffectforDEVIANT_CONDI-TION[F(1,11)=125.
3,p\0.
001],butnosignicantinter-actionbetweenfactors.
DiscussionTheresultsobtainedinthepresentstudyclearlydemon-stratedadifferenceinthehemisphericlateralityofMMNmamplitudesbetweenSDandTDconditions.
TheamplitudesofMMNmevokedbySD(MMNm_SD)wererelativelylargerintheright,whereasthoseevokedbyTD(MMNm_TD)wererelativelylargerintheleft(Figs.
3,4).
NohemisphericdifferencewasobservedintheMMNmlatency;however,thelatenciesofMMNm_TDweresig-nicantlylongerthanthoseofMMNm_SDinboththeleftandrighthemispheresevenwhentheonsettimedifferencebetweenSDandTD(16.
7ms)wasconsidered.
Incontrasttopreviousstudies(Alhoetal.
1998;Shtyrovetal.
2000),whichalsoinvestigatedthehemisphericasymmetryofMMN(m),thetotalsoundinputswereidenticalbetweenSDandTDconditionsinthepresentstudy.
Therefore,thesoundpropertyitselfcannotexplaintheobtainedresults;thedeviationpattern(SDorTD)fromthestandardsoundstreamwassolelyresponsiblefortheresultsobtained.
Weusedband-passlteredclick-trainsthatdidnotconveyspecicmeaningstoensurethathemisphericlateralizationforpre-attentivehumanauditoryprocessing,representedbyMMN(m),wasnotlimitedtothecomplexwaveformsfromnaturalsoundsources(e.
g.
humanvoiceormusicalinstruments),butinpartoriginatedfromearly,low-levelauditoryneuralprocessingdealingwithbasicsoundchar-acteristics,namely,spectralandtemporalfeatures(ZatorreandBelin2001;Tallaletal.
1993;Poeppel2003;Boemioetal.
2005).
Itseemsplausiblethatspectralandtemporalsoundinformationisdifferentiallyencodedintoneuralactivity(BendorandWang2007;Sakaietal.
2009).
Spectralinformationisencodedintothemaximalmovementposi-tionofthebasilarmembraneinthecochlea.
Therefore,incaseoftheSDcondition,thegroupsofinnerhaircellscorrespondingtoSDsoundsweredifferentfromthosecorrespondingtostandardsounds.
Incontrast,TDsoundshadsimilarfrequencycharacteristicstostandardsoundsignals.
Similargroupsofinnerhaircellsonthetonotopicmapinthecochleaareactivated.
InordertodetecttheTDsoundsignal,thecentralauditorysystemshouldanalyzethetemporalpatternsofneuralactivity.
ThepresentresultsdemonstratedthattheMMNmlatencieselicitedbyTDweresignicantlylongerthanthoseelicitedbySD(Figs.
3,4).
First,wehavetoconsiderthetimingoftheSDandTDonsets.
Theoretically,SDisdetectablefromtherstclickoftheTSinthecochlea,whereasTDdetectionrequiresthesecondclickofthe60Hzband-passlteredclicktrainsdeviatedfromthestandard30Hzband-passlteredclicksorthemissingsecondclickofthestandard60Hzband-passlteredclicktrainsduringpresentationofthedeviant30Hzband-passlteredclicktrainstomanifestinthecentralauditorysystem.
Therefore,werstsubtracted16.
7msfromtheMMNm_TDlatencyinordertocompareitwiththeMMNm_SDlatency.
Evenafterthisadjustment,MMNm_TDwassignicantlylongerthanMMNm_SD(Fig.
4),whichsuggestedthatdifferentneuralmechanismscontributetothedetectionofspectralandtemporalsounddeviants.
Neuralencodingofthetemporalpatternsofauditorysignalstooklongerandappearedtotakeplaceatahigherleveloftheauditorysystemthanspectralcoding.
PreviousMEGstudies(Okamotoetal.
2009,2012)alsosupportthishypothesisbydemonstratingthatthetemporalchangeselicitedsignicantlydelayedauditoryN1mresponses,withamajordeectionintheauditoryevokedresponsehavingalatencyofapproximately100ms(Na¨a¨ta¨nenandPicton1987),thanthoseelicitedbyspectralchanges.
AuditoryMMNmisapre-attentiveautomaticbrainresponseelicitedbyanychangeinauditorystimulation(Na¨a¨ta¨nenetal.
2007).
Inthepresentstudy,weusedband-passlteredclicktrainsthatdidnotconveyspecicmeaningandsubjectsweredistractedfromtheauditorymodality;therefore,itislesslikelythatsubjectsinvolun-tarilyprocessedandperceivedthetestsoundsasmusicalorspeechsignals.
Theobtainedresultsindicatedthatthehemisphericasymmetryofauditoryprocessinginhumansstartsfromthebasic,pre-attentiveauditoryprocessinglevel.
Moreover,soundinputswerecompletelycounter-balancedbetweentheSDandTDconditions.
Therefore,thehemisphericasymmetryoftheMMNmresponseselicitedbytheSDandTDcouldnotbeexplainedsolelyby476BrainTopogr(2015)28:471–478123stimulusfeatures.
Thelateralizedmemorytracesofbasicauditoryprocessesintermsofspectralandtemporalsoundfeaturesappeartoberesponsiblefortheresultsobtained.
Recenthumanneuroimagingstudiesrevealedthatthefunctionalhemisphericasymmetryofauditoryprocessingwasnotlimitedtocomplexsoundsignalsconveyingspe-cicmeaningandrules(e.
g.
musicandspeech),butorig-inatedfromthebasicauditoryprocessinglevel,namely,thetemporalintegrationwindow(Poeppel2003;Belinetal.
1998;ZatorreandBelin2001;Zatorreetal.
2002).
Itisimportanttoquicklyandpreciselyencodeenvironmentalsoundsindailylife.
However,becauseofthetrade-offbetweentemporalandspectralanalysisprecision[Acousticuncertaintyprinciple;(Joos1948;Zatorreetal.
2002)],itisimpossibletoachievehighspectralandhightemporalsoundanalysesatthesametimeusingonetemporalinte-grationwindow.
Ashorttemporalintegrationwindowleadstohightemporalresolution,butrelativelylowspec-tralresolutionofthesoundanalyses.
Ontheotherhand,alongtemporalwindowleadstohighspectralresolution,butrelativelylowtemporalresolutionofthesoundanalyses.
Therefore,itseemsplausiblethatthehumanauditorycorticesintheleftandrighthemispheresadoptdifferentintegrationtimewindowsinsteadofapplyingonespecictemporalintegrationtimewindowinbothhemispheres.
Belinetal.
(1998)andPoeppel(2003)hypothesizedthatthelefthemisphereappliedashortertemporalintegrationwindow,resultinginabettertemporalresolutioncapabil-ity,andtherighthemisphereappliedalongertemporalintegrationwindow,resultinginabetterspectralresolutioncapability.
Inthepresentstudy,thelongertemporalinte-grationwindowwithhigherspectralresolutionintherighthemisphereappearstohavedominantlycontributedtodetectingspectrallydeviatedsoundsignalsandresultedinrelativelylargerMMNm_SDamplitudesintherighthemisphere.
Incontrast,theshortertemporalintegrationwindowwithhightemporalresolutioninthelefthemi-sphereappearstohavedominantlyprocessedtemporallydeviatedsoundsignalsandresultedinrelativelylargerMMNm_TDamplitudesinthelefthemisphere.
TheMMNmamplitudesandlatenciesobtainedinthesen-sorspaceandsourcespaceexhibitedsimilarpatterns(Figs.
3,4):theMMNm_SDandMMNm_TDamplitudeswerelargerintherightandlefthemispheres,respectively.
However,theANOVAexaminingMMNmamplitudesresultedinasigni-cantinteractionbetweenDEVIANT_CONDITIONandHEMISPHEREinthesourcespacedata[F(1,11)=6.
67,p\0.
03],butonlyamarginaltrendtowardsignicancewasobservedinthesensorspacedata[F(1,11)=4.
54,p=0.
056].
Themainreasonforthisinconsistencymaybethattheneuralsourcesinonehemispherecouldinuencetheevokedmag-neticeldsinthecontra-lateralmagneticsensors.
Moreover,headsizesandheadpositionsdifferedbetweensubjectsandthecentralsulcusofthesubjectscouldshiftfromthecenteroftheMEGdewar.
Therefore,thesefactorsmayhaveledtoalessrobuststatisticaloutcomeintheRMSamplitudesoftheMMNmresponsesthantheMMNmsourcestrengths.
Inconclusion,usingcarefullyconstructedauditorystimulithatwerecounter-balancedbetweenconditionsandhadcleartime-lockedonsetsofSDandTD,thepresentstudyclearlydemonstratedthatneuralprocessingdealingwithspectrallydeviatedsoundswererelativelydominantintherighthemispherewhilethosedealingwithtemporallydeviatedsoundswererelativelydominantinthelefthemisphere.
Theseresultsstronglysupportthehypothesisthatthehumanbrainadoptsasymmetricmemorytracesofbasicspectralandtemporalsoundfeaturesintheleftandrighthemispheresinordertoimprovethedetectionofdeviantsoundsignals.
AcknowledgmentsWethankY.
Takeshimafortechnicalhelpandourtestsubjectsfortheirdiligentcollaboration.
Thisstudywassupportedbythe''JapanSocietyforthePromotionofScienceforYoungScientists(23689070)'',''StrategicResearchProgramforBrainSciences(Developmentofbiomarkercandidatesforsocialbehavior)'',and''SoundTechnologyPromotionFoundation''.
ConictofinterestTheauthorshavedeclaredthatnocompetinginterestsexist.
OpenAccessThisarticleisdistributedunderthetermsoftheCreativeCommonsAttributionLicensewhichpermitsanyuse,dis-tribution,andreproductioninanymedium,providedtheoriginalauthor(s)andthesourcearecredited.
ReferencesAlhoK(1995)Cerebralgeneratorsofmismatchnegativity(MMN)anditsmagneticcounterpart(MMNm)elicitedbysoundchanges.
EarHear16:38–51AlhoK,ConnollyJF,CheourM,LehtokoskiA,HuotilainenM,VirtanenJ,AulankoR,IlmoniemiRJ(1998)Hemisphericlateralizationinpreattentiveprocessingofspeechsounds.
NeurosciLett258:9–12BelinP,ZilboviciusM,CrozierS,ThivardL,FontaineA(1998)Lateralizationofspeechandauditorytemporalprocessing.
JCognNeurosci10:536–540BelinP,ZatorreRJ,LafailleP,AhadP,PikeB(2000)Voice-selectiveareasinhumanauditorycortex.
Nature403:309–312BendorD,WangXQ(2007)Differentialneuralcodingofacousticutterwithinprimateauditorycortex.
NatNeurosci10:763–771BoemioA,FrommS,BraunA,PoeppelD(2005)Hierarchicalandasymmetrictemporalsensitivityinhumanauditorycortices.
NatNeurosci8:389–395BrocaP(1861)Remarquessurlesiegedelafacultedulangagearticulesuiviesd'uneobservationd'aphemie(pertedelaparole).
BullSocAnat6:330–357DrullmanR,FestenJM,PlompR(1994a)Effectofreducingslowtemporalmodulationsonspeechreception.
JAcoustSocAm95:2670–2680DrullmanR,FestenJM,PlompR(1994b)Effectoftemporalenvelopesmearingonspeechreception.
JAcoustSocAm95:1053–1064BrainTopogr(2015)28:471–478477123EulitzC,DieschE,PantevC,HampsonS,ElbertT(1995)Magneticandelectricbrainactivityevokedbytheprocessingoftoneandvowelstimuli.
JNeurosci15:2748–2755GrifthsTD,JohnsrudeI,DeanJL,GreenGG(1999)AcommonneuralsubstratefortheanalysisofpitchanddurationpatterninsegmentedsoundNeuroreport10:3825–3830HillebrandA,BarnesGR(2002)Aquantitativeassessmentofthesensitivityofwhole-headMEGtoactivityintheadulthumancortex.
Neuroimage16:638–650JamisonHL,WatkinsKE,BishopDV,MatthewsPM(2006)Hemisphericspecializationforprocessingauditorynonspeechstimuli.
CerebCortex16:1266–1275JoosM(1948)Acousticphonetics.
Languagemonograph,vol23.
LinguisticSocietyofAmerica,BaltimoreKujalaT,TervaniemiM,SchrogerE(2007)Themismatchnegativityincognitiveandclinicalneuroscience:theoreticalandmethod-ologicalconsiderations.
BiolPsychol74:1–19LappeC,Steinstra¨terO,PantevC(2013)AbeamformeranalysisofMEGdatarevealsfrontalgeneratorsofthemusicallyelicitedmismatchnegativity.
PLoSOne8:e61296MooreBCJ(2003)Anintroductiontothepsychologyofhearing.
AcademicPress,BostonNa¨a¨ta¨nenR,PictonT(1987)TheN1waveofthehumanelectricandmagneticresponsetosound:areviewandananalysisofthecomponentstructure.
Psychophysiology24:375–425Na¨a¨ta¨nenR,GaillardAW,Ma¨ntysaloS(1978)Earlyselective-attentioneffectonevokedpotentialreinterpreted.
ActaPsychol42:313–329Na¨a¨ta¨nenR,PaavilainenP,RinneT,AlhoK(2007)Themismatchnegativity(MMN)inbasicresearchofcentralauditoryprocess-ing:areview.
ClinNeurophysiol118:2544–2590OkamotoH,StrackeH,DraganovaR,PantevC(2009)Hemisphericasymmetryofauditoryevokedeldselicitedbyspectralversustemporalstimuluschange.
CerebCortex19:2290–2297OkamotoH,TeismannH,KakigiR,PantevC(2012)Auditoryevokedeldselicitedbyspectral,temporal,andspectral-temporalchangesinhumancerebralcortex.
FrontPsychol3:149OldeldRC(1971)Theassessmentandanalysisofhandedness:theEdinburghinventory.
Neuropsychologia9:97–113PoeppelD(2003)Theanalysisofspeechindifferenttemporalintegrationwindows:cerebrallateralizationas'asymmetricsamplingintime'.
SpeechCommun41:245–255SakaiM,ChimotoS,QinL,SatoY(2009)Differentialrepresentationofspectralandtemporalinformationbyprimaryauditorycortexneuronsinawakecats:relevancetoauditorysceneanalysis.
BrainResCognBrainRes1265:80–92ShannonRV,ZengFG,KamathV,WygonskiJ,EkelidM(1995)Speechrecognitionwithprimarilytemporalcues.
Science270:303–304ShtyrovY,KujalaT,PalvaS,IlmoniemiRJ,NaatanenR(2000)Discriminationofspeechandofcomplexnonspeechsoundsofdifferenttemporalstructureintheleftandrightcerebralhemispheres.
Neuroimage12:657–663SzymanskiMD,PerryDW,GageNM,RowleyHA,WalkerJ,BergerMS,RobertsTP(2001)Magneticsourceimagingoflateevokedeldresponsestovowels:towardanassessmentofhemisphericdominanceforlanguage.
JNeurosurg94:445–453TallalP,MillerS,FitchRH(1993)Neurobiologicalbasisofspeech:acaseforthepreeminenceoftemporalprocessing.
AnnNYAcadSci682:27–47TervaniemiM,KujalaA,AlhoK,VirtanenJ,IlmoniemiRJ,Na¨a¨ta¨nenR(1999)Functionalspecializationofthehumanauditorycortexinprocessingphoneticandmusicalsounds:amagnetoencephalographic(MEG)study.
Neuroimage9:330–336TescheCD,UusitaloMA,IlmoniemiRJ,HuotilainenM,KajolaM,SalonenO(1995)Signal-spaceprojectionsofmegdatacharac-terizebothdistributedandwell-localizedneuronalsources.
ElectroencephalogrClinNeurophysiolSuppl95:189–200VosPG,TroostJM(1989)Ascendinganddescendingmelodicintervals—statisticalndingsandtheirperceptualrelevance.
MusicPercept6:383–396WarrierCM,ZatorreRJ(2002)Inuenceoftonalcontextandtimbralvariationonperceptionofpitch.
PerceptPsychophys64:198–207WernickeC(1874)Symptomenkomplex.
EinepsychologischeStudieaufanatomischerBasis.
CohnundWeigert,BreslauZatorreRJ,BelinP(2001)Spectralandtemporalprocessinginhumanauditorycortex.
CerebCortex11:946–953ZatorreRJ,EvansAC,MeyerE(1994)Neuralmechanismsunderlyingmelodicperceptionandmemoryforpitch.
JNeurosci14:1908–1919ZatorreRJ,BelinP,PenhuneVB(2002)Structureandfunctionofauditorycortex:musicandspeech.
TrendsCognSci6:37–46478BrainTopogr(2015)28:471–478123
- dBts60.com相关文档
- thresholdsts60.com
- standbyts60.com
- Predictionts60.com
- topographicalts60.com
- SoftwarePerformanceAnalysis
- 本公司ts60.com
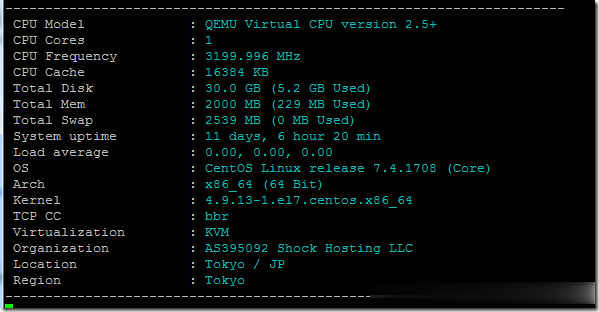
ShockHosting日本机房VPS测试点评
这个月11号ShockHosting发了个新上日本东京机房的邮件,并且表示其他机房可以申请转移到日本,刚好赵容手里有个美国的也没数据就发工单申请新开了一个,这里做个简单的测试,方便大家参考。ShockHosting成立于2013年,目前提供的VPS主机可以选择11个数据中心,包括美国洛杉矶、芝加哥、达拉斯、杰克逊维尔、新泽西、澳大利亚、新加坡、日本、荷兰和英国等。官方网站:https://shoc...
Ceranetworks顶级合作伙伴 香港E3 16G 299元 香港E5 32G 650元 美国E3 16G 650元
提速啦(www.tisula.com)是赣州王成璟网络科技有限公司旗下云服务器品牌,目前拥有在籍员工40人左右,社保在籍员工30人+,是正规的国内拥有IDC ICP ISP CDN 云牌照资质商家,2018-2021年连续4年获得CTG机房顶级金牌代理商荣誉 2021年赣州市于都县创业大赛三等奖,2020年于都电子商务示范企业,2021年于都县电子商务融合推广大使。资源优势介绍:Ceranetwo...
ATCLOUD.NET-OVH海外高防云主机,采用KVM架构,稳定安全且便宜好用,仅3刀起
官方网站:点击访问ATCLOUD.NET官网优惠码:目前提供Cloud VPS与Storage VPS两款产品的六折优惠活动(续费同价,截止至2021年5月31日)优惠码:UMMBPBR20Z活动方案:一、型号CPU内存磁盘流量优惠价格购买链接VPS-1GB0.5×2.6+GHz1GB20GB1TB$3立即购买VPS-2GB1×2.6+GHz2GB50GB2TB$6立即购买VPS-4GB2×2.6...

ts60.com为你推荐
-
乐划锁屏oppofindx2乐划锁屏点进去闪退 是什么情况?对对塔对对塔和魔方格那个是正宗的?月神谭适合12岁男孩的网名,要非主流的,帮吗找找,谢啦钟神发跪求钟神发名言出处,A站大神看过来梦遗姐男人梦遗,女人会吗?铂金血痕仇家血痕是个成语吗?国风商讯国风塑钢质量怎么样www.xiaoyuan.com校园网、校园网站和校园网络的区别?悠达网大学生校园网购有什么好的平台可以推荐给我吗?查看源代码如何查看软件源代码