2.5bnb998.com
bnb998.com 时间:2021-04-08 阅读:()
ControlofsomaticembryogenesisandembryodevelopmentbyAP2transcriptionfactorsSouadElOuakfaouiJaimieSchnellAshrafAbdeenAdamColvilleHele`neLabbeShuyouHanBernardBaumSergeLabergeBrianMikiReceived:17November2009/Accepted:22July2010/Publishedonline:27August2010TheAuthor(s)2010.
ThisarticleispublishedwithopenaccessatSpringerlink.
comAbstractMembersoftheAP2familyoftranscriptionfactors,suchasBABYBOOM(BBM),playimportantrolesincellproliferationandembryogenesisinArabidopsisthaliana(AtBBM)andBrassicanapus(BnBBM)buthowthisoccursisnotunderstood.
WehaveisolatedthreeAP2genes(GmBBM1,GmAIL5,GmPLT2)fromsomaticembryoculturesofsoybean,Glycinemax(L.
)Merr,anddiscoveredGmBBM1tobehomologoustoAtBBMandBnBBM.
GmAIL5andGmPLT2werehomologoustoArabidopsisAINTEGUMENTA-like5(AIL5)andPLETH-ORA2(PLT2),respectively.
ConstitutiveexpressionofGmBBM1inArabidopsisinducedsomaticembryosonvegetativeorgansandotherpleiotropiceffectsonpost-germinativevegetativeorgandevelopment.
SequencecomparisonsofBBMorthologuesrevealedthepresenceoftensequencemotifsoutsideoftheAP2DNA-bindingdomains.
Oneofthemotifs,bbm-1,wasspecictotheBBM-likegenes.
Deletionanddomainswapanalysesrevealedthatbbm-1wasimportantforsomaticembryo-genesisandactedcooperativelywithatleastoneothermotif,euANT2,intheregulationofsomaticembryogene-sisandembryodevelopmentintransgenicArabidopsis.
TheresultsprovidenewinsightsintothemechanismsbywhichBBMgovernsembryogenesis.
SouadElOuakfaoui,JaimieSchnell,AshrafAbdeenandAdamColvillehavecontributedequally.
ElectronicsupplementarymaterialTheonlineversionofthisarticle(doi:10.
1007/s11103-010-9674-8)containssupplementarymaterial,whichisavailabletoauthorizedusers.
S.
ElOuakfaouiJ.
SchnellA.
AbdeenA.
ColvilleH.
LabbeS.
HanB.
BaumB.
Miki(&)AgricultureandAgri-FoodCanada,960CarlingAve,Ottawa,ONK1A0C6,Canadae-mail:mikib@agr.
gc.
caS.
ElOuakfaouie-mail:Souad.
ElOuakfaoui@ec.
gc.
caJ.
Schnelle-mail:Jaimie.
Schnell@inspection.
gc.
caA.
Abdeene-mail:Ashraf.
Abdeen@mail.
mcgill.
caA.
Colvillee-mail:Adam.
Colville@IOGEN.
caH.
Labbee-mail:labbehe@agr.
gc.
caS.
Hane-mail:hans@agr.
gc.
caB.
Baume-mail:bernard.
baum@agr.
gc.
caA.
ColvilleBiologyDepartment,CarletonUniversity,Ottawa,ONK1S5B6,CanadaS.
ElOuakfaouiS.
LabergeAgricultureandAgri-FoodCanada,2560HochelagaBlvd,Quebec,QCG1V2J3,Canadae-mail:laberges@agr.
gc.
caPresentAddress:S.
ElOuakfaouiBiotechnologySection,EmergingPrioritiesDivision,EnvironmentCanada,FontaineBuilding,7thoor,#775,200Sacre-Coeur,Gatineau,QuebecK1A0H3,CanadaPresentAddress:J.
SchnellPlantandBiotechnologyRiskAssessmentUnit,CanadianFoodInspectionAgency,1400MerivaleRoad,Ottawa,OntarioK1A0Y9,Canada123PlantMolBiol(2010)74:313–326DOI10.
1007/s11103-010-9674-8KeywordsAP2/ANTArabidopsisBABYBOOMEmbryogenesisSoybeanTranscriptionfactorIntroductionBABYBOOM(BBM)isamemberoftheAP2familyoftranscriptionfactorswhichhavediversefunctionsinplantdevelopment(Nole-Wilsonetal.
2005;FloydandBowman2007;Fengetal.
2005).
TheAP2familybelongstotheAP2/ERFsuperfamily.
Thisisoneofthelargestgroupsofplanttranscriptionfactorsandhasundergoneextensiveduplicationanddomainshufingduringitsevolution(Ri-echmannetal.
2000;Kimetal.
2006;Nakanoetal.
2006).
MembershavedoubleAP2/ERFdomainsintheAP2family,singleAP2/ERFDNA-bindingdomainsintheERFfamilyandsingleAP2/ERFdomainstogetherwithaB3DNA-bindingdomainintheRAVfamily(RiechmannandMeyerowitz1998).
Thesuperfamilyconsistsof147membersinArabidopsis,157membersinrice(Nakanoetal.
2006)and148membersinsoybean(Zhangetal.
2008).
Membersfunctionindiverseprocessesfundamentaltoplantgrowth,reproductionandenvironmentalinterac-tions(RiechmannandMeyerowitz1998;Fengetal.
2005;Nole-Wilsonetal.
2005).
InArabidopsis,riceandsoybean(Sakumaetal.
2002;Gongetal.
2004;Nakanoetal.
2006;Zhangetal.
2008)thememberscanbegroupedbysequencesimilarityintothesamefamilyandsubfamilygroupings(FloydandBowman2007).
BBMclusterswithinoneofthesublineages,euANT,whichappearstohavespecializedinmeristemdifferentiationandmaintenance(FloydandBowman2007).
FunctionalstudiesoftheAP2familymembers,suchasAPETALA2(AP2),AINTEGUMENTA(ANT),BABYBOOM(BBM),PLETHORA1(PLT1),PLETHORA2(PLT2)andtheAINTEGUMENTA-like(AIL)genes,haverevealeddiversetranscriptionalnetworksanddevelop-mentalprocessesthatthefamilycontrolsaswellasredundanciesthatexistsamongandwithinthedifferentgroups.
APETALA2(AP2),therstmemberofthefamilytobereported,functionsindependentlyinthespecicationoforalorganidentity(Jofukuetal.
1994;Okamuroetal.
1997,Maes1999)andinthemaintenanceofthestemcellnicheoftheshootmeristem(Wu¨rschumetal.
2006).
AINTEGUMENTA(ANT)isrequiredforovuledevelop-mentandoralorgangrowth(Elliottetal.
1996,Klucheretal.
1996).
ANTcanactredundantlywithAP2inoraldevelopment(Krizeketal.
2000).
BABYBOOM(BBM)hasbeenimplicatedinthedifferentiationofembryonalstemcellsfromsomaticcells(Boutilieretal.
2002)andclusterswithinthesamecladeasPLETHORA(PLT1andPLT2)whichcontrolsrootstemcellidentityandmaintenance(Aidaetal.
2004).
PLT1,PLT2,BBMandPLT3/AIL6functionredundantlyinrootmeristemandembryodiffer-entiation(Galinhaetal.
2007).
TheyarealsocloselyrelatedtoanumberofotherAINTEGUMENTA-like(AIL)genes(Nole-Wilsonetal.
2005;Tsuwamotoetal.
2010)whicharegenerallyinvolvedinthespecicationofmer-istemsordivision-competentstates(Nole-Wilsonetal.
2005).
MembersoftheAP2familysharetwohighly-conservedAP2DNA-bindingdomainrepeatsseparatedbyalinkerregion;however,theN-terminalandC-terminalsequencesareverydistinct.
Thedomainswithintheseregionshavenotbeenstudiedbuttheyarelikelytobeimportantforthespecictranscriptionalactivities,proteininteractionsandnuclearlocalizationsthatconfertheuniquefunctionsassociatedwitheachmember(Nakanoetal.
2006).
SequencecomparisonsoftheAP2/ERFsuperfamilymem-bersfromsoybean,ArabidopsisandricehaverevealedthepresenceofmanyconservedmotifsoutsideoftheAP2/ERFDNAbindingdomainraisingthepossibilitythatsharedconservedmotifsmayformthebasisforfunctionalsimi-laritiesamongdifferentgroups(Zhangetal.
2008).
BABYBOOM(BnBBM)wasclonedfromBrassicanapusmicrosporeembryoculturesandwasshowntoinducesomaticembryoswhenectopicallyoverexpressedinArabidopsisorB.
napus(Boutilieretal.
2002).
Theacquisitionoftotipotencythroughthisprocesswasaccom-paniedbyanumberofpleiotropiceffectsonplantdevelop-ment(Boutilieretal.
2002).
Intransgenictobacco,BnBBMexpressioninducedpleiotropiceffectsonvegetativegrowthanddevelopmentbutdidnotinduceembryogenesis(Srinivasanetal.
2007)indicatingthatembryogenicpath-waysdifferamongspeciesorthatthedomainswithinBBMthatgovernembryogenesishavedivergedinsequenceamongplantsandwerenotrecognized.
ItiscurrentlybelievedthatBnBBMenhancescellproliferationsthatcanresultindifferentdevelopmentaloutcomesincludingorganogenesisorembryogenesis(Srinivasanetal.
2007).
Thevarietyofdifferentpleiotropiceffectsonplantdevel-opmentthatwereobservedwithectopically-expressedBnBBMmayindicatebroadredundanciesamongAP2familyPresentAddress:A.
AbdeenDepartmentofBiology,McGillUniversity,1205DocteurPeneld,RoomN5/2Montreal,QuebecH3A1B1,CanadaPresentAddress:A.
ColvilleIogenCorporation,400HuntClubRoad,Ottawa,OntarioK1V1C1,CanadaPresentAddress:S.
HanAgricultureandAgri-FoodCanada,1391SandfordStreet,London,OntarioN5V4T3,Canada314PlantMolBiol(2010)74:313–326123membersbeyondthosealreadydemonstratedwiththePLT-likeandBBMgenes(Galinhaetal.
2007)ortheAP2andANTgenes(Krizeketal.
2000).
InthisstudyweexaminedBBMorthologuesfromanon-cruciferousspecies,soybean,todetermineifthesamedevelopmentalpathwayswereinducedintransgenicplants;toidentifyconservedmotifsinBBM-likegenes;andtolocatethedeterminantsofembryogenesis.
GmBBM1wasidentiedasthefunctionalorthologueofAtBBMandBnBBMthroughbothstructuralandfunctionalstudies.
ThemodulardomainstructureofBBM-likegeneswasalsoanalyzedandrevealedmotifsthatwereimportantfortheirspecicityintheinductionofsomaticembryogenesis.
MaterialsandmethodsPlantmaterialEmbryoswereisolatedfromsoybean,Glycinemax(L.
)MerrilgenotypeX5,formerlycalledX2650-7-2-3(Sim-mondsandDonaldson2000).
Donorplantsweregrownundera16hphotoperiodinthegreenhouseaspreviouslyreported(SimmondsandDonaldson2000).
Embryogeniccultureswereestablishedaspreviouslydescribed(Finer1988;FinerandNagasawa1988).
Immaturecotyledons,4and5.
9mminsize,wereplacedabaxialside-downonMSD40medium(FinerandNagasawa1988)containingMSsalts,B5vitamins,6%(w:v)sucrose,40mg/l2,4-Dand0.
2%Gelrite,pH5.
8andculturedat27°Cunderaphoto-periodof16hwithalightintensityof60–70lmolm-2s-1providedbycool-whiteuorescentlamps.
Secondaryglob-ularembryosdevelopingontheculturedcotyledonsweretransferredto125mlErlenmeyeraskswith30mlof10A40Nmedium(FinerandNagasawa1988)containingmodiedMSsalts(withMSnitrogenreplacedby10mMNH4NO3and30mMKNO3),B5vitamins,6%sucrose,5mg/l2,4-D,and5mMasparagine,pH5.
8andculturedasaboveat30lmolm-2s-1onarotaryshakerat125rpm.
Embryogenictissue(30–75mg)wassubculturedtofreshmediumevery2–4weekstomaintainproliferation.
Prolif-erationwasstoppedandembryomaturationwasinitiatedbyculturingembryogenictissuefor4weeksonsolidOMSM6mediumwithoutcharcoalunderalightintensityof70lmolm-2s-1.
Embryosweredesiccatedin85%relativehumidityandgerminatedonB5mediumwith3%sucroseand0.
6%Phytagaror0.
2%Gelrite,pH5.
8.
Plantswithtwotrifoliateleavesweretransplantedtosoilandgrownundera16-hphotoperiodpriortotransfertoa12-hphotoperiodforseedformation.
Roottissueswereharvestedfrom1-weekoldseedlings.
Leafandstemtissueswerefrom3-weeksoldseedlings.
Flowerswereharvested2daysafterfullanthesis.
Developingseedsandpodswereharvestedfromdifferentstagesbetween6and24dayspostanthesis(dpa)at3daysintervals.
TostudytheexpressionofGmBBM1genesindevelopingseedstissuesfromseedcoat,embryoandpodswerecollectedseparatelyandRNAwasextractedusingtheRNeasyKit(Qiagen,Canada).
Tissuesfromsoybeanroots,leaves,stemsandowerswerealsocollected,homoge-nizedinliquidnitrogenandtotalRNAwasextractedusingTrizol(Invitrogen,Canada).
RT-PCRProbesgeneratedbyRT-PCRwereusedforlibraryscreeningandexpressionproling.
Primersspecictoeachofthethreesoybeangenes,GmBBM1,GmPLT2andGmAIL5,areshowninSupplementaryTable1.
TheSoy-Tub2genewasusedaspositivecontrol.
PCRconditionsforGmPLT2andGmAIL5wereasfollows:3minat95°C;30cycles[1min95°C,1minspecicannealingtemperature(SupplementaryTable1),1min72°C]and5minexten-sionat72°C.
PCRconditionsforGmBBM1andSoyTub2wereasfollows:31minat50°C,15minat95°C;30cycles(30s94°C,30sspecicannealingtemperature(Supple-mentaryTable1),30s72°C)and2minextensionat72°C.
LibraryconstructionTotalRNA(2mg/gfreshtissue)wasextractedfromculturedembryogenicsoybeansuspensionculturesandmRNA(0.
2%oftotalRNA)waspuriedusingtheQuikmRNAIsolationKit(Stratagene/VWR,Canada).
AcDNAlibrarywascon-structedusingLambda-ZAPIIcDNAsynthesiskit(Strata-gene/VWR,Canada).
Phages(1.
69106pfu)wereplatedaccordingtothemanufacturer'sinstructions.
TheampliedcDNAlibrarytitrewas2.
59109pfu.
ThecDNAlibraryphageplaqueswerehybridizedusingaprobecontainingtheAP2domainandlinkerusingprimersdesignedfromthealignmentofESTAW200688codingforasoybeanAP2proteinandBnBBM1fromBrassicanapusasdescribedinResults.
cDNAwereexcisedfromthelambda-ZAPIIvectorsfollowingtheconversionofthelambda-ZAPcloneplasmidspBluescriptSKDNA.
SequencesweredeterminedwiththeABIPRISMTMdyeterminatorcyclesequencingkit(PEAppliedBiosystems).
DatawasanalyzedusingDNAsis(HitachiCorporation2003)GenejumperprimerinsertionkitforsequencingversionB(Invitrogen,Canada)wasusedinordertobeabletosequencethefull-lengthclones.
ArabidopsistransformationandassessmentPlanttransformationswerecarriedoutaccordingtoCloughandBent(1998).
GmBBM1wasclonedintothepBINPLUSvectorandexpressionwasdrivenbythedouble35SPlantMolBiol(2010)74:313–326315123promoterandalfalfamosaicvirus(AMV)translationalenhancerasdescribedinBoutilieretal.
(2002).
Selectionwasperformedon50mg/lkanamycin.
Thousandsofseedswerescreeneduntilacollectionofapproximately35lineswithsomaticembryoswererecovered.
Ofthese,sixrep-resentativelineswerestudiedingreaterdetailoversuc-cessivegenerations.
BnBBM1mutantsweregeneratedbyinversePCRreactionsontheBnBBM1PUC19plasmid,usingExpandLongTemplatePCRSystem(Roche)andsyntheticprimerswith10-bpsequenceincludingaBamHIorXhoIsitesintheir50-endsthatreplacestheoriginalsequences.
ThefollowingprimerswereusedforeuANT2(N1BamHIF50-CGGGATCCCTGAGAAATCAACCCGTGGATAATG-30andN1BamHIR50-CTGGATCCGCCACCACCACCGTCTCCTCCTC-30)andforbbm-1(N2XhoIF50-CGGCTCGAGCCTTATGAACAAAATCACCATCG-30andN2XhoIR50-CCGCTCGAGGGTGGAAGTATTTGAAAGAAAT-30).
TheseconstructswereobtainedbyKpnI/SalIdigestionandclonedintothebinaryvectorpCAMBIA1300,alongwiththewild-typeBnBBM1geneasthecontrol.
Theywereintro-ducedintoAgrobacteriumtumefaciensGV3101forplanttransformationandselectionwasperformedwith30mg/Lhygromycin.
Approximately,3,000–4,000seedsintotalwerescreenedintriplicateexperimentstoassessthephe-notypesofthetransgeniclines.
Forthedomainswapanalysis,thePLT2,AIL5,andAIL7genesfromArabidopsisweresynthesizedwiththeBnBBM1bbm-1motif(SLGLSMIKTWLRNQP)asshowninFig.
2AbyGenScriptCorporation(Piscataway,NJ).
TheywereclonedintopCAMBIA1300undercontrolofthedouble35Spromoter.
Asabovetheconstructswereintro-ducedintoAgrobacteriumtumefaciensGV3103forplanttransformationandscreened.
Seedsfromeachtransgenicseedlineweresterilizedinasolutionof70%ethanolfor30sfollowedbyasolutionof25%javexplus0.
05%Tritonfor20–30min.
Theseedswereshakenfortheduration.
Theywerethenrinsed4timeswithsteriledistilledwaterandplacedina0.
1%solutionofaga-roseat4°Covernight.
Thenextday,theseedswereplatedonMSB5media(SigmaChemicalCo)with3%sucroseand0.
6%agarosealongwithselectableagents:50mg/lkana-mycinforpBINPLUSor30mg/lhygromycinforpCAM-BIA1300.
Seedlingswerethensurveyedforphenotypeovera2–3weekperiod.
At3–4weeksgrowththeyweretrans-ferredtosoilandbroughttooweringstage.
PhylogeneticanalysisSequencesofdoubleAP2domainproteinssimilartoAtBBMwereminedfromGenBank(NationalInstitutesofHealthgeneticsequencedatabaseatNationalCenterforBiotechnologyInformation)usingtheEntrezbrowser(availableat:http://www.
ncbi.
nlm.
nih.
gov/)andfromtheTIGRGeneIndices.
Proteinsthatwerenotfull-lengthwerenotincludedasweintendedtoeventuallyassesstheN-terminalandtheC-terminalsequencesalso.
ToidentifythosesequencesbelongingtotheeuANTsubgroup,sequenceswerescreenedforthe10aminoacidinsertionintherstAP2domaincharacteristicofthisgroupaswellasapredomainregionlongerthan127aminoacids,asdescribedbyKimetal.
(2006).
WRI1fromthebasa-lANTsubgroup,aseparateANTlineage,wasincludedasanoutgroup.
TheminedaminoacidsequenceswerealignedusingCLUSTALW(Thompsonetal.
1994)andfurtherimprovedbyvisualexaminationandeditingusingGeneDocVersion2.
6.
002(NicholasandNicholas1997).
TheAP2domainwasusedforalignmentasthefulllengthsequencesweretoodivergenttosuccessfullyalignallthesequencesretrievedfromGeneBankandTIGRGeneIndicesdatabase.
WelimitedtheboundariesofthedoubleAP2domaintothesequencesstarting5aminoacidsupstreamandapproximately10aminoacidsdownstreamoftheconserveddoubleAP2domains.
Intotal,53sequences(SupplementaryTable2)wereanalyzed.
Thealignedsequenceswererstsubjectedtoatoolfortheselectionofthebesttmodelfromamong112modelsofproteinevolution,usingProtTest(Abascaletal.
2005)version2.
2.
Afterthelikelihoodstatisticswerecompletedthreestatisticalframeworkswereselectedconsecutivelyoneatatimetodeterminewhichcandidatemodeltsthedatabest.
Althoughatreeusingthemodel-averagedesti-mateoftheparameterswasobtainedintheanalysisandexamined,wecarriedaseparateMaximumLikelihood(ML)phylogeneticanalysisusingthePROMLprogram(PROteinMaximumLikelihoodprogram)inthePHYLIPpackage(Felsenstein2008)withtheselectedevolutionarymodelalongwithitsspecicparameters,previouslyobtainedfromProtTest.
Intheanalysis100bootstraptreeswereproducedfromwhichamajorityruletreewascom-putedtoobtainbootstrapsupportforthebranchesontheMLtreeandastrictconsensustreewasalsocomputedforexamination.
Anothersimilarbootstrapanalysiswascar-riedwithMEGA4(NeiandKumar2000)with1,000repeats.
ConservedmotifswereidentiedthroughacombinationofClustalWalignment,MEMEversion4.
0.
0(BaileyandElkan1994),andBlockMaker(Henikoffetal.
1995).
TheEukaryoticLinearMotifresourceforFunctionalSitesinProtein(http://elm.
eu.
org/links.
html)wasusedtopredictmotiffunction.
316PlantMolBiol(2010)74:313–326123ResultsAP2familygenesclonedfromsoybeansomaticembryosSoybeancDNAswithsequencessimilartoBnBBM1,wereisolatedfromimmaturesomaticembryosofsoybean,G.
maxcvMerrilgenotypeX5(SimmondsandDonaldson2000).
TheprobeusedtoscreenthecDNAlibrarywasstrategicallydesignedusingPCRprimers(SupplementaryTable1)targetedtosequencesidentiedfromalignmentsofBnBBM1andapartialsoybeanputativeAP2proteinESTAW200688fromacDNAlibraryconstructedfromcotyledonsof3-and7-day-oldseedlingsofcultivarWilliams(http://www.
ncbi.
nlm.
nih.
gov/entrez/viewer.
fcgidb=nucest&val=6481417)(ShoemakerR,ThePublicSoybeanESTProject)(BiogeneticsServices,SD,USA)sequences.
Twoclones,GmBBM1(2.
5kb,707aa,GenBankAccessionHM775856)andGmAIL5(2.
3kb,558aa,GenBankAccessionHM775857),wereidentiedandiso-lated.
AnothercDNAclone(2Kb,560aa),GmPLT2,wasrstidentiedastwo50-trunctedcDNAclonesthatlaterappearedasafulllengthclone(EU677381).
AllpossessedhighlevelsofsequencesimilarityinthedoubleAP2domainregion(97%)whichdecreasedintheN-andC-terminalregions(SupplementaryFig.
1A).
ThesoybeanGmBBM1sequencewassimilartothoseofthecruciferousgenes,BnBBM1andAtBBM,withmorethan91%simi-larityinthedoubleAP2domains(SupplementaryFig.
1B).
ThisdecreasedintheN-andC-terminalsequencesto48and30%respectively(SupplementaryFig.
1B).
PhylogeneticanalysisofGmBBM1,GmAIL5andGmPLT1ToexaminetherelationshipsofthethreesoybeangeneswithotherAP2familymembersallavailablegenesinTheAra-bidopsisInformationResource(http://www.
arabidopsis.
org/),GenBank,DFCI(DanaFarberCancerInstitute)GeneIndex(http://compbio.
dfci.
harvard.
edu/tgi/;formerlyTIGRgeneindex),andtheTIGRRiceGenomeAnnotationProjectDatabase(http://rice.
plantbiology.
msu.
edu)atthetimeofthestudywithsequencesimilaritytoBBMgeneswereretrievedusingthedoubleAP2domainregionoftheAtBBMsequence.
Onlyfull-lengthsequencesandthosewithatleast60%similarityinthedoubleAP2domainwereretained.
SequencesbelongingtotheeuANTsubgrouparecharacterizedbythepresenceofa10aminoacidinsertintherstAP2domainregionandarelativelylongpredomainregion(Kimetal.
2006).
Thesetwocharacteristicswereusedtoselect49sequencesbelongingtotheeuANTsubgroup.
WRI1,belongingtothebasalANTsubgroup,wasincludedasanoutgroup.
Atotalof53sequences(SupplementaryTable2)wereincludedinthephylogeneticanalysis.
ThealignmentwascarriedoutusingtheregionspanningthetwoAP2domains,includingthelinkerregion,starting5residuesupstreamoftherstconservedAP2domainandnishingapproximately10aminoacidsdownstreamofthesecondAP2domain.
TheevolutionarymodelthatbesttthisdatawastheJTTGmodel(i.
e.
theJones-Taylor-Thorntonmodel)underthethreestatisticalframeworksanalyzed,i.
e.
AIC,AICcandBIC,anditsparameterG=0.
558,i.
e.
gammashapewith4ratecategories.
Figure1showstherelationshipofthethreesoybeangenestotheeuANTsubgroupbasedontheAP2-linker-AP2sequences.
GmBBM1isclosesttotheotherlegumeBBMgene,MtBBM,fromMedicagotruncatulaandalsoclusterscloselywiththeBBMgenesfromtheBrassicaceaandPoaceaefamilies.
GmAIL5isgroupedwiththeoilpalmEgAp2-1geneandisclosetoAIL5fromArabidopsis.
GmPLT2isgroupedwiththeArabidopsisPLT1andPLT2genesalongwithagenefromVitisvinifera,allofwhichformthelargerPLT-likegroupingwithArabidopsisAIL6andAIL7andsequencesfromrice,Brassicarapa,andanothersequencefromVitisvinifera.
MotifcompositionandidenticationofaBBM-specicmotifTofurtherdistinguishthegenesintheBBM-likegroupingfromothereuANTgenes,themorevariableN-terminalandC-terminalproteinsequenceswerealignedtosearchforconservedmotifs(seeSupplementaryFig.
1C).
IntheN-terminalsequences,vemotifswereidentiedinmostmembersoftheBBM-likegrouping.
ThreeofthesemotifswerepreviouslyidentiedintheeuANTlineage(euANT2,3,and4;Kimetal.
2006)andwerefoundthroughoutmostofthesequencesanalyzed(Fig.
2A).
Thefourthmotif(bbm-1)wasspecictotheBBM-likegenesandabsentfromtheclosely-relatedPLT-likegenes(Fig.
2A).
ThismotifwasweaklyconservedinsomeoftheANT-likegenes,particularlyGbANTL1,PtANTL1,andCAO45633.
ItwasalsofoundinOs01g67410,whichwasnotassociatedwithanyoftheidentiedgroupings.
Thefthmotif(euANT5)wasalsoidentiedinthemajorityofthemembersoftheBBM-likegroupingbutwasabsentintheBBM-likegenesfromCenchrusciliarisandPennisetumsquamulatum(Fig.
2A).
ThismotifisalsoconservedintheANT-likegenesaswellastheAIL1-likegenes,andrepre-sentativescontainingthismotifcanalsobefoundamongstthePLT-likegenes,althoughitiscompletelyabsentfromtheAIL5-likegenes.
Interestinglythebbm-1andeuANT5motifsboresomesimilarity,bothincludingthesequenceLSM.
IntheC-terminalsequencesveothermotifswereidentied(Fig.
2B).
Amotif(euANT6)wasidentiedinallPlantMolBiol(2010)74:313–326317123oftheBBM-likegenesimmediatelyfollowingthesecondAP2domain.
ItwasidentiedinrepresentativesfromallofthemajoreuANTlineages,althoughthedegreeofcon-servationvariedconsiderably(Fig.
2B).
Thismotifcon-tainsanumberofLysandArgresidues,suggestingthatitmayatleastpartiallyfunctionasanuclearlocalizationsignal,ashasbeensuggestedforthePLTgenes(Aidaetal.
2004).
ThefouradditionalmotifsintheC-terminalsequencesoftheBBM-likegeneswereconservedwithalowdegreeofspecicity.
Thebbm-2motifwasonlyweaklyconservedintheArabidopsisandB.
napusBBMgenesandbbm-3wascompletelyabsent.
SomeofthesemotifswereweaklyconservedinsequencesoutsideoftheBBM-likegroupingparticularlybbm-3andbbm-4insomeofthePLT-likesequences.
Aswiththebbm-1motifs,Os01g67410alsocontainedthebbm-2andbbm-4motifs,althoughthebbm-3motifwasonlyweaklyconservedandthebbm-5motifwasabsent.
Severalofthemotifs,includingeuANT2,bbm-1,euANT5,andeuANT6,containconsensussequencesforphosphorylation.
euANT5alsocontainsaconsensussequenceforsumoylation.
TheeuANT2motifalsocon-formstoaTRFHdomaindockingmotifandaWWligandmotif.
ExpressionproleofGmBBM1,GmAIL5andGmPLT1Insoybeanplants,theexpressionofGmBBM1paralleledthatofAtBBMinArabidopsis(Boutilieretal.
2002).
Itwasselectivelyexpressedinsoybeanembryosandyoungroots(Fig.
3).
GmAIL5alsopossessedthesameexpressionpat-ternasAIL5inArabidopsis(Nole-Wilsonetal.
2005).
Itwasexpressedinallofthesoybeanorgansundergoinggrowthanddevelopmentthatwereexamined(Fig.
3).
GmPLT2hadthesameexpressionpatternasPLT1andPLT2inArabidopsis(Aidaetal.
2004)andM.
truncatula(Iminetal.
2007;Holmesetal.
2008).
Itwasexpressedpredominantlyinsoybeanrootsandtoalesserextentinsoybeanembryos(Fig.
3).
Thedatasupportedtheassign-mentofGmBBM1asaBBMhomologue,GmAIL5asanFig.
1MaximumlikelihoodtreeoftheeuANTsubgroupofthedoubleAP2family318PlantMolBiol(2010)74:313–326123Fig.
2MotifsintheN-terminal(A)orC-terminal(B)sequencesoftheBBM-likegenesandtheirconservationintheeuANTsubgroupoftheAP2family.
Residuesthatmatchtheconsensussequenceareinred.
Sequencesaregroupedintocladesasindicatedinthephylogenetictree.
ThesequencesoftheeuANT2andbbm-1motifsintheBnBBM1sequencethatweredeletedtotesttheirfunctionareunderlined.
Forthedomainswapanalysis,thesiteofinsertionofthebbm-1motiffromBnBBM1(SLGLSMIKTWLRNQP)inPLT2,AIL5,andAIL7areindicatedwitharrowheadsPlantMolBiol(2010)74:313–326319123AIL5homologueandGmPLT2asaPLThomologuebasedonthegenephylogeny,conservationoftheN-terminalandC-terminalsequencesandtheexpressionpatterns.
PhenotypesgeneratedbyBBM-likegenesintransgenicArabidopsisAcomparisonoftheembryogenicphenotypeandpleio-tropiceffectsconferredbyGmBBM1andBnBBM1wascarriedoutintransgenicArabidopsisusingthepBINPLUSvectorsystemusedpreviouslyforectopicoverexpressionofBnBBM1(Boutilieretal.
2002)(Fig.
4).
Weconrmedpreviousdescriptions(Boutilieretal.
2002)thatshowedBnBBM1expressioninArabidopsisinducedsomaticembryosandcotyledon-likestructuresonpost-germinationorgansoftransgenicseedlingsaswellasauniquesetofpleiotropiceffectsonboththevegetativeandreproductiveorgans(Table1,BnBBM1lines1–3).
Approximately35independenttransgenicArabidopsislinesexpressingGmBBM1embryosonthecotyledonswereexaminedandsixrepresentativelineswerefollowedintotheT2genera-tions(Fig.
4A–C).
Table1showthedatafor3oftheGmBBM1lines.
Somaticembryogenesisgenerallyoccur-redatalowerfrequencythanwithBnBBM1(Table1).
Thehighestfrequencywas28%intheGmBBM1line1,fol-lowedby17%intheGmBBM1line2and5%intheGmBBM1line3(n[100foreachline).
Embryosalsoemergedfromtheshootapicalmeristem(Fig.
4D)andhypocotyls(Fig.
4E).
PleiotropiceffectsondevelopmentthatweretypicalofBnBBM1overexpressionwereagainevidentwithGmBBM1overexpression.
Theseincludedshortroots(80%penetrance,GmBBM1line3;n=100),Fig.
3AccumulationofGmBBM1,GmAIL5,GmPLT1andtubulin(accessionnumberM21297)mRNAinsoybeancvMerrilgenotypeX5detectedbyRT-PCR.
Rootsat1week(R),stemat3weeks(S),leafat3weeks(L),owersat2dpa(F),seedpodat21dpa(SP),seedcoatsat21dpa(SC),embryosat21dpa(E)Fig.
4PhenotypesinducedbyexpressionofGmBBM1intransgenicArabidopsisseedlings.
Somaticembryosdifferentiatingfromthetipsofcotyledons(A,B,C).
Ectopicrootsdifferentiatingfromtheembryogenictissues(A,C).
Somaticembryosdifferentiatingfromtheshootapicalmeristem(D)andhypocotyls(E).
Alterationsinseedlingdevelopmentcharacterizedbyshortroots,swollenhypocotylsandelongatedcotyledons(E,F).
Un-transformedcontrolseedlingsforcomparison(G)320PlantMolBiol(2010)74:313–326123shorthypocotyls(greaterthan80%penetranceinallGmBBM1linestested),elongatedcotyledons(lowpene-trancephenotype\25%inallGmBBM1lines)onseed-lings(Fig.
4E,F).
Laterindevelopmentarangeofpleiotropiceffectswerenotedincludingalteredleafmor-phologiesandthickenedoralstems.
Increasednumbersofinorescenceswerealsonoted(datanotshown).
Duringthereproductivephaseofgrowth,GmBBM1andBnBBM1bothinduceddelayed-oweringtime;increasedoralorgansize;thickenedandincreasednumbersofinores-cences;decreasedsiliquesize;anddecreasedseedpro-duction(Table1).
AlthoughmanyofthephenotypesweresimilarforBnBBM1andGmBBM1someinterestingdifferenceswereobserved.
Forexample,inGmBBM1transgeniclinesectopicrootsoccassionallydevelopedoncotyledonsalongwithembryos(Fig.
4A,C),afeaturethatwasnotreportedwithBnBBM1(Boutilieretal.
2002).
Furthermore,elon-gatedrootswereseenonplantletsoftheGmBBM1lineincontrasttotheshort-rootphenotypeobservedinBnBBM1plants(Table1).
Functionofthebbm-1motifinembryogenesisThesignicanceofthebbm-1sequencemotiftoBnBBM1functionwasexaminedbycreatinga9-aminoaciddeletionmutationintheBnBBM1gene(Fig.
2A)andassessingfunctionalityintransgenicArabidopsis(Fig.
5).
Forcom-parison,adeletionofmotifeuANT2(Fig.
2A)wasalsocreatedsinglyandtogetherwithadeletionofbbm-1.
Theinductionofsomaticembryosoncotyledonsbywild-typeBnBBM1wasrstconrmedin22transgeniclinesascontrolsusingthepCAMBIA1300vector(Fig.
5C).
Shootssubsequentlydevelopedinculture(Fig.
5D)andplantletsregeneratedasdescribedpreviouslybyBoutilieretal.
2002(datanotshown).
SomaticembryogenesiswasnoteliminatedbydeletionofeuANT2(Fig.
5G,H)oreuANT2andbbm-1together(Fig.
5I,J).
However,themutantphe-notypesdifferedinthattheembryo-likestructurestendedtoproliferateandshootsdidnotemergeovertime(Fig.
5H,J)aswithwild-typeBnBBM1(Fig.
5D).
Thisphenotypewasconrmedin14and25transgeniclines,respectivelyandwasinstrikingcontrasttothephenotypegeneratedbydeletionofthebbm-1motifalonewhichwascharacterizedbythecompletelossofsomaticembryogenesisoncotyle-donsaswellastheaccompanyingpleiotropiceffectsonseedlings(Fig.
5E,F).
Pleiotropiceffectsassociatedwiththismutationemergedatlaterstagesofplantdevelopment.
SomeweresimilartotheclassIIpleiotropiceffectsdescribedforBnBBM1byBoutilieretal.
(2002).
Forexample,owerswithshortsepalsandpetalsrelativetocarpelswerefrequentlyobserved(Fig.
6A)comparedwithwildtypeowers(Fig.
6B).
Thechangesinleafmorphol-ogydescribedforBnBBM1(Boutilieretal.
2002)wereinfrequentlyobserved;however,serratedmarginswereoccasionallyobserved(Fig.
6C)inplantsexpressingthedeletionmutantbutnotwildtypeplants(Fig.
6D).
Thebbm-1motifwasinsertedintoArabidopsisPLT2,AIL5andAIL7(Fig.
2A)todetermineifothereuANTmemberspossessedthecomplementaryelementsneededforsomaticembryogenesisintransgenicArabidopsis.
AIL7bbm-1andAIL5bbm-1failedtogenerateectopicembryosinexperimentsparalleltothoseabove(datanotshown).
TheectopicPLT2phenotypewaspre-viouslycharacterizedbytheproliferationofectopicrootsandroothairsongerminatingembryosorseedlings(Aidaetal.
2004).
PLT2bbm-1generatedarangeofpheno-typesinvedifferenttransgeniclines.
Rapidandstrongtissueproliferationfollowedbycopiousembryodifferen-tiationwasobservedin4ofthe5lines(Fig.
7A,B).
ThisphenotyperesembledtheBBMphenotype(BenScheresandcolleagues,personalcommunication;Fig.
7).
SmallerTable1PhenotypesofindividualtransgeniclinesexpressingBnBBM1andGmBBM1constitutivelyintransgenicArabidopsisDescriptionofphenotypeTransgenicBnBBM1linesTransgenicGmBBM1linesLine1Line2Line3Line1Line2Line3SomaticembryoformationoncotyledonsShortroots,brousrootsElongatedrootsThick,shorthypocotylsElongatedcotyledonsDelayedoweringtimeReducedseedsetIncreasedoralorgansizeThephenotypesinboldattheseedlingstageofdevelopmentandusedfortheinitialscreeningoflines.
Thefrequencyofoccurrenceofthephenotypewasrecordedforeachlineusingatleast100plantlets([0\25%),(25–50%50–75%75–100%)PlantMolBiol(2010)74:313–326321123undenedgreentissueshadalsobeendescribedaspartofthePLT2ectopicphenotype(Aidaetal.
2004)In2ofthelines,bothBBMandPLT2phenotypeswereexpressedsimultaneously.
Intheselinesembryodifferentiationwaslessprolicandrootdifferentiationwasveryabundant(Fig.
7C,D).
Fig.
5Phenotypeofun-transformedArabidopsisseedlingsat14days(A)and21–28days(B)postgerminationcomparedwiththephenotypeof14dayand21–28daytransgenicArabidopsisectopicallyexpressingwild-typeBnBBM1(C,Drespectively),theBnBBM1deletionmutantwithoutbbm-1(E,Frespectively),theBnBBM1deletionmutantwithouteuANT2(G,Hrespectively),theBnBBM1deletionmutantwithouteuANT2andbbm-1(I,Jrespectively)322PlantMolBiol(2010)74:313–326123Fig.
6PleiotropiceffectsontransgenicArabidopsisower(A)andleaf(C)morphologiesgeneratedbytheBnBBM1deletionmutantthatlacksthebbm-1motifcomparedwithwildtypeowers(B)andleaf(D)Fig.
7PhenotypesoftransgenicArabidopsisseedlings,inwhichthebbm-1motiffromBnBBM1wasinsertedintothecorrespondingpositionofthePLT2gene,at11days(A,B),13days(C)and17days(D)postgerminationPlantMolBiol(2010)74:313–326323123DiscussionThephenotypesassociatedwiththeAP2/ANTfamilyfrequentlyinvolveundifferentiatedcellproliferationanddifferentiationofstemcellnicheswithinmeristems(Nole-Wilsonetal.
2005).
Recently,aroleinembryogenesishasalsoemerged.
MostoftheinformationhasbeengeneratedbytheinductionofsomaticembryogenesisbyectopicoverexpressionofBnBBM1inArabidopsisandBnapus(Boutilieretal.
2002;Passarinhoetal.
2008).
GmBBM1wasshownheretoinducesimilardevelopmentaleventsinArabidopsis.
Comparedwithmeristematiccelltypes,thedifferentiationofembryonalstemcellsfromsomaticcellsispoorlyunderstood.
Studieshaveshownthatsomaticembryogenesismaybeactivatedthroughanumberofdifferentpathwaysandbyanumberofdifferentgenes(Passarinhoetal.
2008;reviewedbyVerdeiletal.
2007).
Inthisstudy,wehaveidentiedspecicfeaturesoftheBBM-likehomologuesthatareresponsiblefortheinductionofembryogenesisandthatmayseparatetheBBM-likegenesfromcloselyrelatedmembersoftheAP2/ANTfamilythatfunctioninvegetativedevelopmentyetactredundantlyinembryodevelopment.
TounderstandhowBBM-likegenesfunctionintheconversionofsomaticcellstoembryonalcellsandthedevelopmentoftheresultingembryoitmaybeimportanttounderstandtherelationshipsamongtheAP2/ANTfamilymembers.
ThemembersoftheeuANTlineage,suchastheBBM-like,PLT-likeandAIL5-likegenes,areverycloselyrelatedphylogeneticallyandstructurally.
Ouranalysishasrevealeddomainconservationamongthembutalsodomainspecicity.
Forexample,thebbm-1motifwasspecictoalloftheBBM-likegenes.
Thedifferentmembersareknowntohavespecicrolesindevelopmentbutevidenceforredundancyisalsoemerging.
Forexample,themutantphenotypesofbbmandplt2haveshownthatAtBBMandPLT2functionasredundantpartnersinearlyArabidopsisembryogenesis(Galinhaetal.
2007).
AtBBM,PLT1,PLT2andPLT3/AIL6areexpressedinrootprimordiawheretheyfunctionasredundantpartnersinrootstemcelldifferen-tiationinembryosandrootdifferentiation(Aidaetal.
2004;Galinhaetal.
2007).
Thedemonstrationthattheadditionofthebbm-1motiftoPLT2enhancedthecapacitytoinducesomaticembryogenesisconrmedthecloserelationshipbetweenthesegenesandmayhaverevealedakeystructuralfeatureofBBMimportantforitsabilitytofunctioninembryogenesis.
ThePLT2ectopicphenotypewasnotlostwhentheBnBBM1ectopicphenotypewasexpressedbutthestrengthofthePLT2andBnBBM1ectopicphenotypeswereinverselyrelatedindicatingthattheembryogenicandrootmeristematicpathwayscouldbecompetinginthesetransgeniclines.
HowAIL5functions,particularlyinembryogenesis,isunclear.
AIL5appearstobeinvolvedincellproliferationactivitiesinmanyorgansandmaygenerateenlargedorgans,forexamplelargeowers,whenectopicallyexpressed(Nole-Wilsonetal.
2005).
ThisphenotypeissimilartothatofANT,whichgenerateslargeowers(Nole-Wilsonetal.
2005;MizukamiandFischer2000)andAP2whichgenerateslargeembryos(Ohtoetal.
2005)whenectopicallyexpressed.
SinceAIL5mutantshavenoapparentphenotypeitisbelievedthatitactsredundantlywithotherrelatives(Nole-Wilsonetal.
2005).
AIL5andBBMdoublemutantsappeartolackdefectsinembryoorrootmeristemdevelopmentthereforeadditionalunidentiedpartnersmayexistiftheyactinthesamepathways(Tsuwamotoetal.
2010).
Thendingthatectopicover-expressionofAIL5yieldedphenotypessimilartoBBMover-expression,includingsomaticembryogenesis(Tsuwamotoetal.
2010),indicatesthatitcouldbeinvolvedintheproliferationofthetotipotentsomaticcells.
TheembryonicstructuresthatemergedexhibitelevatedexpressionofAGL15,LEC1,LEC2,FUS3andBBM(Tsuwamotoetal.
2010)allofwhichhavethecapacitytoinducesomaticembryogenesiswhenectopicallyexpressed.
Interesting,theoilpalmAIL5-likegene,EgAp2-1,isalsoexpressedinproliferatingtis-sues,especiallyembryonictissuesbutinducescallusgrowthandshootorganogenesisratherthanembryosinArabidopsis(Morcilloetal.
2007).
BnBBM1alsoinducesshootorganogenesisintobaccoratherthanembryogenesisrevealingthatBBM-likegeneshavethecapacitytoinducebothshootmeristemactivityaswellasembryogenesisdependingonthegeneticandcellularenvironmentoftheproliferatingcells(Srinivasanetal.
2007).
UnlikePLT2,theadditionofthebbm-1motiftoAIL5didnotgeneratethecapacitytoinduceembryogenesisintransgenicArabidopsisinthisstudy.
WhetherthisalterationdisruptedthecapacityofAIL5togeneratesomaticembryosasdescribedbyTsuwamotoetal.
(2010)isunknownandpointstocomplexrolesforAIL5inplantdevelopment.
Conservedsequencemotifshavebeenrecognizedamongsoybean,riceandArabidopsisAP2/ANTgenesandsoybean-specicmotifsareamongthem(Zhangetal.
2008).
Previousstudieshavesuggestedthatwithinsub-familiesconservedsequencemotifsareabundantandlikelyreectsharedfunctions(Nakanoetal.
2006).
Unlikesomeofthewell-characterizedmotifsoftheERFfamily,sequencemotifswithintheAP2/ANTfamilyhavenotyetbeenassignedspecicrolesinthetranscriptionalregula-tionofdevelopmentalprocesses(Nakanoetal.
2006;Zhangetal.
2008).
ThecharacterizationoftheGmBBM1genehereallowedustoaligntheBBM-likegenesfromdivergentspecieswhichareknowntogenerateectopicembryosinArabidopsisandthusprovideafunctionaldata324PlantMolBiol(2010)74:313–326123setthatwasusedtoidentify10conservedsequencemotifsoutsideoftheAP2-linker-AP2region.
AmongtheBBM-like,PLT-likeandAIL5-likegenesthemotifcompositionwasverysimilar,however,thebbm-1motifwasthemostspecictotheBBM-likegenesintheN-terminalregion.
IntheC-terminalregionsequenceswithalowerdegreeofconservationamongtheBBM-likegenesexist.
Ofthesebbm-2,bbm-3andbbm-4arepoorlyconservedinthecruciferousgenesandthusunlikelytobeessentialforembryogenesis.
Themotif,bbm-5,isalsopoorlyconservedrelativetobbm-1.
Bothdeletionanddomainswapanalysesrevealedthatthebbm-1motifwasneededforBnBBM1toinducesomaticembryogenesisinArabidopsis.
Furtherexaminationrevealedthatbbm-1activityisintimatelylinkedtotheactivitiesofothermotifswithinthesetranscriptionfactors,especiallyeuANT2,whichisfoundinalmostallothergenesintheeuANTlineageincludingPLT2andAIL5.
DeletionofeuANT2singlyoreuANT2andbbm-1simultaneouslypre-ventedthesomaticembryosfromgeneratingshootsevenafterprolongedtimesinculture.
Amongthemanypossibleexplanationsforbbm-1functionisthepossibilitythattheeuANT2motiffunctionsinvegetativepathwaysofdevel-opmentandthatbbm-1actstorestrainthevegetativeactivityofBnBBM1.
Ifcorrect,thenamechanismmustexisttorelievetherestraintsoneuANT2inBnBBM1duringthelaterstagesofembryodevelopmentwhentheshootandrootmeristemsdifferentiateanddevelop.
Furthermore,ifasingledomain,bbm-1,canseparatetheectopicphenotypesofredundantpartnerssuchasBBMandPLT2thenitwouldseempossiblethatthesharedmotifscouldprovidethebasisforfunctionalredundancy.
AhierarchicalprocessmustexistinthecellularenvironmenttorecruitandcoordinatethedifferentAP2/ANTmemberstothedifferentdevelopmentalpathwaysinanorderlymanner.
Asboththebbm-1andeuANT2motifshavetheconsensussequenceforphosp-orylationpost-translationalprocessesmaybeinvolvedintheregulationofsuchinteractions.
ForPLT-likegenes,expressiongradientsappeartoplayanimportantinthetransitionfromrootmeristeminitiationtorootdevelopment(Galinhaetal.
2007).
AnygeneralmodelfortheroleofAP2/ANTtranscriptionfactorsinsomaticembryogenesisshouldconsidertheobservationsthatBBM-likegenesmayplaydistinctrolesatdifferentstagesofdevelopment;interactwithredundantpartners;andinducedifferentpathwaysofdevelopmentdependingonthegeneticandcellularenvironment.
Thebbm-1motifofBBM-likegenesappearstoplayanearlyroleintheconversionofsomaticcellstoneoplasticembryonalcells.
Thismayoccurbysuppressingtheactivityofothermotifs,suchaseuANT2,whichfunctioninvegetativecelldifferentiation.
ThesharedmotifsamongtheAP2/ANTmembersmayprovideamechanismforBBMtorecruittheactivitiesofmemberswithmeristematicactivitiesessentialforthedevelopmentoftheembryoonceembryogenesisisestablished.
Asnewmembersarerecruitedtothedevel-opingembryodilutionofthebbm-1motifmayreleasethecellsfromtheneoplasticstateandallowthemeristemstodevelop.
SustainedectopicexpressionofBBMgenesdur-ingplantmaturationresultsinarangeofpleiotropiceffects.
ThismayoccurthroughsimilarunintendedinteractionswithAP2/ANTproteinsthroughsharedmotifsthatdisrupttheirnormalpatternsofactivity.
ThedifferentpleiotropiceffectsobservedforGmBBM1andBnBBM1mayrevealdifferencesintheefcienciesofinteractionsresultingfromsequencedivergence.
IfBBM-likegenesfunctionbyuti-lizingredundancieswithotherAP2/ANTtranscriptionfactorstorecruitdevelopmentalpathwaysneededforembryogenesisandembryodevelopmentinseedsthenstrictspatialcontrolofBBM-likegeneexpressionwouldbeessentialfornormalplantdevelopment.
AsshownhereandinallotherstudiesofBBM–likegeneexpression(Boutilieretal.
2002;Nole-Wilsonetal.
2005;Iminetal.
2007)expressioninseedsisunderstrictspatialanddevelopmentalregulationasaretheotherAP2/ANTfamilymembers(Fengetal.
2005).
AcknowledgmentsTheauthorsareverygratefultoLiWangforArabidopsistransformation,MichelleLopezforthetransgenicstud-ies,WayneStanglefortheconstruct,DainaSimmonds,SherylHubbardandSuquinZhengforthesoybeantissues,RejeanDesa-gagneandDavidGagnefortheirsupportandcooperationandBenScheresandhislabforexaminationofourdatapriortopublication.
TheresearchwasfundedbytheCanadianBiotechnologyStrategy.
OpenAccessThisarticleisdistributedunderthetermsoftheCreativeCommonsAttributionNoncommercialLicensewhichper-mitsanynoncommercialuse,distribution,andreproductioninanymedium,providedtheoriginalauthor(s)andsourcearecredited.
ReferencesAbascalF,ZardoyaR,Posada,D(2005)ProtTestselectionofbesttmodelsofproteinevolution.
Bioinformatics21:2104–2105.
http://darwin.
uvigo.
es/software/prottest.
htmlVersion2.
2acces-sed28Sept.
2008AidaM,BeisD,HeidstraR,WillemsenV,BliloumI,GalinhamC,NussaumemL,NohmY-S,AmasinomR,ScheresB(2004)ThePLETHORAgenesmediatepatterningofthearabidopsisrootstemcellniche.
Cell119:109–120BaileyTL,ElkanC(1994)Fittingamixturemodelbyexpectationmaximizationtodiscovermotifsinbiopolymers.
In:AltmanR,BrutlagD,KarpP,LathropR,SearlsD(eds)Proceedingsofthesecondinternationalconferenceonintelligentsystemsformolecularbiology.
AmericanAssociationforArticialIntelli-gencePress,MenloPark,CA,pp28–36BoutilierK,OffringaR,SharmaVK,KieftH,OuelletT,ZhangL,HattoriJ,LiuCM,vanLammerenAA,MikiBL,CustersJB,vanLookerenCampagneMM(2002)EctopicexpressionofBABYBOOMtriggersaconversionfromvegetativetoembryonicgrowth.
PlantCell14:1737–1749PlantMolBiol(2010)74:313–326325123CloughSJ,BentAF(1998)Floraldip:asimpliedmethodforAgrobacterium-mediatedtransformationofArabidopsisthali-ana.
PlantJ16:735–743ElliottRC,BetznerAS,HuttnerE,OakesMP,TuckerWQJ,GerentesD,PerezP,SmythDR(1996)AINTEGUMENTA,anAPET-ALA2-likegeneofarabidopsiswithpleiotropicrolesinovuledevelopmentandoralorgangrowth.
PlantCell8:155–168FelsensteinJ(2008)PHYLIP,PhylogenyInferencePackage,version3.
68.
Availableathttp://evolution.
gs.
washington.
edu/phylip.
html.
AccessedNovember2008FengJ-X,LiuD,PanY,GongW,MaL-G,LuoJ-C,DengXW,ZhuY-X(2005)AnannotationupdateviacDNAsequenceanalysisandcomprehensiveprolingofdevelopmental,hormonalorenvironmentalresponsivenessoftheArabidopsisAP2/EREBPtranscriptionfactorgenefamily.
PlantMolBiol59:853–868FinerJJ(1988)Apicalproliferationofembryogenictissueofsoybean[Glycinemax(L.
)Merrill].
PlantCellRep7:238–241FinerJJ,NagasawaA(1988)Developmentofanembryogenicsuspensioncultureofsoybean[Glycinemax(L.
)Merrill].
PlantCellTissueOrgCult15:125–136FloydSK,BowmanJL(2007)Theancestraldevelopmentaltoolkitoflandplants.
IntJPlantSci168:1–35GalinhaC,HofhuisH,LuijtenM,WillemsenV,BlilouI,HeidstraR,ScheresB(2007)PLETHORAproteinsasdose-dependentmasterregulatorsofArabidopsisrootdevelopment.
Nature449:1053–1056GongW,ShenY-P,MaL-G,PanY,DuY-L,WangD-H,YangJ-Y,HuL-D,LiuX-F,DongC-X,MaL,ChenY-H,YangX-Y,GaoY,ZhuD,TanX,MuJ-Y,ZhangD-B,LiuY-L,Dinesh-KumarSP,LiY,WangX-P,GuH-Y,QuL-J,BaiS-N,LuY-T,LiJ-Y,ZhaoJ-D,ZuoJ,HuangH,DengXW,ZhuY-X(2004)Genome-wideORFeomecloningandanalysisofarabidopsistranscriptionfactorgenes.
PlantPhysiol135:773–782HenikoffS,HenikoffJG,AlfordWJ,PietrokovskiS(1995)Auto-matedconstructionandgraphicalpresentationofproteinblocksfromunalignedsequences.
Gene163:GC17–GC26HolmesP,GoffardN,WeillerGF,RolfeBG,IminN(2008)TranscriptionalprolingofMedicagotruncatulameristematicrootcells.
BMCPlantBiol8:21IminN,NizamidinM,WuT,RolfeBG(2007)FactorsinvolvedinrootformationinMedicagotruncatula.
JExpBot58:439–451JofukuKD,BoerB,MontaguMV,OkamuroJK(1994)ControlofarabidopsisowerandseeddevelopmentbythehomeoticgeneAPETALA2.
PlantCell6:1211–1225KimS,SoltisPS,WallK,SoltisDE(2006)PhylogenyanddomainevolutionintheAPETALA2-likegenefamily.
MolBiolEvol23:107–120KlucherKM,ChowH,ReiserL,FischerRL(1996)TheAINTE-GUMENTAgeneofarabidopsisrequiredforovuleandfemalegametophytedevelopmentisrelatedtotheoralhomeoticgeneAPETALA2.
PlantCell8:137–153KrizekBA,ProstV,MaciasA(2000)AINTEGUMENTApromotespetalidentityandactsasanegativeregulatorofAGAMOUS.
PlantCell12:1357–1366MaesT(1999)TheinorescencearchitectureofPetuniahybridaismodiedbytheArabidopsisthalianaAp2gene.
DevGenet25:199–208MizukamiY,FischerRL(2000)Plantorgansizecontrol:AINTE-GUMENTAregulatesgrowthandcellnumbersduringorgano-genesis.
ProcNatlAcadSciUSA97:942–947MorcilloF,GallardA,PillotM,JouannicS,Aberlene-BertossiF,CollinM,VerdeilJL,TregearJW(2007)EgAP2–1,andAINTEGUMENTA-like(AIL)geneexpressedinmeristematicandproliferatingtissuesofembryosinoilpalm.
Planta226:1353–1362NakanoT,SuzukiK,FujimuraT,ShinshiH(2006)Genome-wideanalysisoftheERFgenefamilyinArabidopsisandrice.
PlantPhysiol140:411–432NeiM,KumarS(2000)MolecularEvolutionandPhylogenetics.
OxfordUniversityPress,NewYorkNicholasKB,NicholasHBJr(1997)GeneDoc:atoolforeditingandannotatingmultiplesequencealignments.
Distributedbytheauthors.
www.
psc.
edu/biomed/genedoc/AccessedOct-Dec2008Nole-WilsonS,TranbyTL,KrizekBA(2005)AINTEGUMENTA-like(AIL)genesareexpressedinyoungtissuesandmayspecifymeristematicordivision-competentstates.
PlantMolBiol57:613–628OhtoM,FischerRL,GoldbergRB,NakamuraK,HaradaJJ(2005)ControlofseedmassbyAPETALA2.
ProcNatlAcadSciUSA102:3123–3128OkamuroJK,CasterB,VillarroelR,VanMontaguM,JofukuKD(1997)TheAP2domainofAPETALA2denesalargenewfamilyofDNAbindingproteinsinArabidopsis.
ProcNatlAcadSciUSA94:7076–7081PassarinhoP,KetelaarT,XingM,vanArkelJ,MaliepaardC,HendriksMW,JoosenR,LammersM,HerdiesL,denBoerB,vanderGeestL,BoutilierK(2008)BABYBOOMtargetgenesprovidediverseentrypointsintocellproliferationandcellgrowthpathways.
PlantMolBiol68:225–237RiechmannJL,MeyerowitzEM(1998)TheAP2/EREBPfamilyofplanttranscriptionfactors.
BiolChem379:633–646RiechmannJL,HeardJ,MartinG,ReuberL,JiangC-Z,KeddieJ,AdamL,PinedaO,RatcliffeOJ,SamahaRR,CreelmanR,PilgrimM,BrounP,ZhangJZ,GhandehariD,ShermanBK,YuG–L(2000)Arabidopsistranscriptionfactors:genome-widecomparativeanalysisamongeukaryotes.
Science290:2105–2110SakumaY,LiuQ,DubouzetJG,AbeH,ShinozakiK,Yamaguchi-ShinozakiK(2002)DNA-bindingspecicityoftheERF/AP2domainofArabidopsisDREBs,transcriptionfactorsinvolvedindehydration-andcold-induciblegeneexpression.
BiochemBiophysResCommun290:998–1009SimmondsDH,DonaldsonPA(2000)Genotypescreeningforproliferativeembryogenesisandbiolistictransformationofshort-seasonsoybeangenotypes.
PlantCellRep19:485–490SrinivasanC,LiuZ,HeidmannI,SupenaENJ,FukuokaH,JoosenR,LambalkJ,AngenentG,ScorzaR,CustersJBM,BoutilierK(2007)HeterologousexpressionoftheBABYBOOMAP2/ERFtranscriptionfactorenhancestheregenerationcapacityoftobacco(NicotianatabacumL).
Planta225:341–351ThompsonJD,HigginsDG,GibsonTJ(1994)CLUSTALW:improvingthesensitivityofprogressivemultiplesequencealignmentthroughsequenceweighting,position-specicgappenaltiesandweightmatrixchoice.
NucleicAcidsRes22:4673–4680TsuwamotoR,YokoiS,TakahataY(2010)ArabidopsisEMBRY-OMAKERencodinganAP2domaintranscriptionfactorplaysakeyroleindevelopmentalchangefromvegetativetoembryonicphase.
PlantMolBiol.
doi:10.
1007/s11103-010-9634-3VerdeilJ-L,AlemannoL,NiemenakN,TranbargerTJ(2007)Pluripotentversustotipotentplantstemcells:dependenceversusautonomyTrendsPlantSci12:1360–1385Wu¨rschumT,Grob-HardtR,LauxT(2006)APETALA2regulatesthestemcellnicheintheArabidopsisshootmeristem.
PlantCell18:295–307ZhangG,ChenM,ChenX,XuZ,GuanS,LiL-C,LiA,GuoJ,MaoL,MaY(2008)Phylogeny,genestructures,andexpressionpatternsoftheERFgenefamilyinsoybean(GlycinemaxL.
).
JExpBot59:4095–4107326PlantMolBiol(2010)74:313–326123
ThisarticleispublishedwithopenaccessatSpringerlink.
comAbstractMembersoftheAP2familyoftranscriptionfactors,suchasBABYBOOM(BBM),playimportantrolesincellproliferationandembryogenesisinArabidopsisthaliana(AtBBM)andBrassicanapus(BnBBM)buthowthisoccursisnotunderstood.
WehaveisolatedthreeAP2genes(GmBBM1,GmAIL5,GmPLT2)fromsomaticembryoculturesofsoybean,Glycinemax(L.
)Merr,anddiscoveredGmBBM1tobehomologoustoAtBBMandBnBBM.
GmAIL5andGmPLT2werehomologoustoArabidopsisAINTEGUMENTA-like5(AIL5)andPLETH-ORA2(PLT2),respectively.
ConstitutiveexpressionofGmBBM1inArabidopsisinducedsomaticembryosonvegetativeorgansandotherpleiotropiceffectsonpost-germinativevegetativeorgandevelopment.
SequencecomparisonsofBBMorthologuesrevealedthepresenceoftensequencemotifsoutsideoftheAP2DNA-bindingdomains.
Oneofthemotifs,bbm-1,wasspecictotheBBM-likegenes.
Deletionanddomainswapanalysesrevealedthatbbm-1wasimportantforsomaticembryo-genesisandactedcooperativelywithatleastoneothermotif,euANT2,intheregulationofsomaticembryogene-sisandembryodevelopmentintransgenicArabidopsis.
TheresultsprovidenewinsightsintothemechanismsbywhichBBMgovernsembryogenesis.
SouadElOuakfaoui,JaimieSchnell,AshrafAbdeenandAdamColvillehavecontributedequally.
ElectronicsupplementarymaterialTheonlineversionofthisarticle(doi:10.
1007/s11103-010-9674-8)containssupplementarymaterial,whichisavailabletoauthorizedusers.
S.
ElOuakfaouiJ.
SchnellA.
AbdeenA.
ColvilleH.
LabbeS.
HanB.
BaumB.
Miki(&)AgricultureandAgri-FoodCanada,960CarlingAve,Ottawa,ONK1A0C6,Canadae-mail:mikib@agr.
gc.
caS.
ElOuakfaouie-mail:Souad.
ElOuakfaoui@ec.
gc.
caJ.
Schnelle-mail:Jaimie.
Schnell@inspection.
gc.
caA.
Abdeene-mail:Ashraf.
Abdeen@mail.
mcgill.
caA.
Colvillee-mail:Adam.
Colville@IOGEN.
caH.
Labbee-mail:labbehe@agr.
gc.
caS.
Hane-mail:hans@agr.
gc.
caB.
Baume-mail:bernard.
baum@agr.
gc.
caA.
ColvilleBiologyDepartment,CarletonUniversity,Ottawa,ONK1S5B6,CanadaS.
ElOuakfaouiS.
LabergeAgricultureandAgri-FoodCanada,2560HochelagaBlvd,Quebec,QCG1V2J3,Canadae-mail:laberges@agr.
gc.
caPresentAddress:S.
ElOuakfaouiBiotechnologySection,EmergingPrioritiesDivision,EnvironmentCanada,FontaineBuilding,7thoor,#775,200Sacre-Coeur,Gatineau,QuebecK1A0H3,CanadaPresentAddress:J.
SchnellPlantandBiotechnologyRiskAssessmentUnit,CanadianFoodInspectionAgency,1400MerivaleRoad,Ottawa,OntarioK1A0Y9,Canada123PlantMolBiol(2010)74:313–326DOI10.
1007/s11103-010-9674-8KeywordsAP2/ANTArabidopsisBABYBOOMEmbryogenesisSoybeanTranscriptionfactorIntroductionBABYBOOM(BBM)isamemberoftheAP2familyoftranscriptionfactorswhichhavediversefunctionsinplantdevelopment(Nole-Wilsonetal.
2005;FloydandBowman2007;Fengetal.
2005).
TheAP2familybelongstotheAP2/ERFsuperfamily.
Thisisoneofthelargestgroupsofplanttranscriptionfactorsandhasundergoneextensiveduplicationanddomainshufingduringitsevolution(Ri-echmannetal.
2000;Kimetal.
2006;Nakanoetal.
2006).
MembershavedoubleAP2/ERFdomainsintheAP2family,singleAP2/ERFDNA-bindingdomainsintheERFfamilyandsingleAP2/ERFdomainstogetherwithaB3DNA-bindingdomainintheRAVfamily(RiechmannandMeyerowitz1998).
Thesuperfamilyconsistsof147membersinArabidopsis,157membersinrice(Nakanoetal.
2006)and148membersinsoybean(Zhangetal.
2008).
Membersfunctionindiverseprocessesfundamentaltoplantgrowth,reproductionandenvironmentalinterac-tions(RiechmannandMeyerowitz1998;Fengetal.
2005;Nole-Wilsonetal.
2005).
InArabidopsis,riceandsoybean(Sakumaetal.
2002;Gongetal.
2004;Nakanoetal.
2006;Zhangetal.
2008)thememberscanbegroupedbysequencesimilarityintothesamefamilyandsubfamilygroupings(FloydandBowman2007).
BBMclusterswithinoneofthesublineages,euANT,whichappearstohavespecializedinmeristemdifferentiationandmaintenance(FloydandBowman2007).
FunctionalstudiesoftheAP2familymembers,suchasAPETALA2(AP2),AINTEGUMENTA(ANT),BABYBOOM(BBM),PLETHORA1(PLT1),PLETHORA2(PLT2)andtheAINTEGUMENTA-like(AIL)genes,haverevealeddiversetranscriptionalnetworksanddevelop-mentalprocessesthatthefamilycontrolsaswellasredundanciesthatexistsamongandwithinthedifferentgroups.
APETALA2(AP2),therstmemberofthefamilytobereported,functionsindependentlyinthespecicationoforalorganidentity(Jofukuetal.
1994;Okamuroetal.
1997,Maes1999)andinthemaintenanceofthestemcellnicheoftheshootmeristem(Wu¨rschumetal.
2006).
AINTEGUMENTA(ANT)isrequiredforovuledevelop-mentandoralorgangrowth(Elliottetal.
1996,Klucheretal.
1996).
ANTcanactredundantlywithAP2inoraldevelopment(Krizeketal.
2000).
BABYBOOM(BBM)hasbeenimplicatedinthedifferentiationofembryonalstemcellsfromsomaticcells(Boutilieretal.
2002)andclusterswithinthesamecladeasPLETHORA(PLT1andPLT2)whichcontrolsrootstemcellidentityandmaintenance(Aidaetal.
2004).
PLT1,PLT2,BBMandPLT3/AIL6functionredundantlyinrootmeristemandembryodiffer-entiation(Galinhaetal.
2007).
TheyarealsocloselyrelatedtoanumberofotherAINTEGUMENTA-like(AIL)genes(Nole-Wilsonetal.
2005;Tsuwamotoetal.
2010)whicharegenerallyinvolvedinthespecicationofmer-istemsordivision-competentstates(Nole-Wilsonetal.
2005).
MembersoftheAP2familysharetwohighly-conservedAP2DNA-bindingdomainrepeatsseparatedbyalinkerregion;however,theN-terminalandC-terminalsequencesareverydistinct.
Thedomainswithintheseregionshavenotbeenstudiedbuttheyarelikelytobeimportantforthespecictranscriptionalactivities,proteininteractionsandnuclearlocalizationsthatconfertheuniquefunctionsassociatedwitheachmember(Nakanoetal.
2006).
SequencecomparisonsoftheAP2/ERFsuperfamilymem-bersfromsoybean,ArabidopsisandricehaverevealedthepresenceofmanyconservedmotifsoutsideoftheAP2/ERFDNAbindingdomainraisingthepossibilitythatsharedconservedmotifsmayformthebasisforfunctionalsimi-laritiesamongdifferentgroups(Zhangetal.
2008).
BABYBOOM(BnBBM)wasclonedfromBrassicanapusmicrosporeembryoculturesandwasshowntoinducesomaticembryoswhenectopicallyoverexpressedinArabidopsisorB.
napus(Boutilieretal.
2002).
Theacquisitionoftotipotencythroughthisprocesswasaccom-paniedbyanumberofpleiotropiceffectsonplantdevelop-ment(Boutilieretal.
2002).
Intransgenictobacco,BnBBMexpressioninducedpleiotropiceffectsonvegetativegrowthanddevelopmentbutdidnotinduceembryogenesis(Srinivasanetal.
2007)indicatingthatembryogenicpath-waysdifferamongspeciesorthatthedomainswithinBBMthatgovernembryogenesishavedivergedinsequenceamongplantsandwerenotrecognized.
ItiscurrentlybelievedthatBnBBMenhancescellproliferationsthatcanresultindifferentdevelopmentaloutcomesincludingorganogenesisorembryogenesis(Srinivasanetal.
2007).
Thevarietyofdifferentpleiotropiceffectsonplantdevel-opmentthatwereobservedwithectopically-expressedBnBBMmayindicatebroadredundanciesamongAP2familyPresentAddress:A.
AbdeenDepartmentofBiology,McGillUniversity,1205DocteurPeneld,RoomN5/2Montreal,QuebecH3A1B1,CanadaPresentAddress:A.
ColvilleIogenCorporation,400HuntClubRoad,Ottawa,OntarioK1V1C1,CanadaPresentAddress:S.
HanAgricultureandAgri-FoodCanada,1391SandfordStreet,London,OntarioN5V4T3,Canada314PlantMolBiol(2010)74:313–326123membersbeyondthosealreadydemonstratedwiththePLT-likeandBBMgenes(Galinhaetal.
2007)ortheAP2andANTgenes(Krizeketal.
2000).
InthisstudyweexaminedBBMorthologuesfromanon-cruciferousspecies,soybean,todetermineifthesamedevelopmentalpathwayswereinducedintransgenicplants;toidentifyconservedmotifsinBBM-likegenes;andtolocatethedeterminantsofembryogenesis.
GmBBM1wasidentiedasthefunctionalorthologueofAtBBMandBnBBMthroughbothstructuralandfunctionalstudies.
ThemodulardomainstructureofBBM-likegeneswasalsoanalyzedandrevealedmotifsthatwereimportantfortheirspecicityintheinductionofsomaticembryogenesis.
MaterialsandmethodsPlantmaterialEmbryoswereisolatedfromsoybean,Glycinemax(L.
)MerrilgenotypeX5,formerlycalledX2650-7-2-3(Sim-mondsandDonaldson2000).
Donorplantsweregrownundera16hphotoperiodinthegreenhouseaspreviouslyreported(SimmondsandDonaldson2000).
Embryogeniccultureswereestablishedaspreviouslydescribed(Finer1988;FinerandNagasawa1988).
Immaturecotyledons,4and5.
9mminsize,wereplacedabaxialside-downonMSD40medium(FinerandNagasawa1988)containingMSsalts,B5vitamins,6%(w:v)sucrose,40mg/l2,4-Dand0.
2%Gelrite,pH5.
8andculturedat27°Cunderaphoto-periodof16hwithalightintensityof60–70lmolm-2s-1providedbycool-whiteuorescentlamps.
Secondaryglob-ularembryosdevelopingontheculturedcotyledonsweretransferredto125mlErlenmeyeraskswith30mlof10A40Nmedium(FinerandNagasawa1988)containingmodiedMSsalts(withMSnitrogenreplacedby10mMNH4NO3and30mMKNO3),B5vitamins,6%sucrose,5mg/l2,4-D,and5mMasparagine,pH5.
8andculturedasaboveat30lmolm-2s-1onarotaryshakerat125rpm.
Embryogenictissue(30–75mg)wassubculturedtofreshmediumevery2–4weekstomaintainproliferation.
Prolif-erationwasstoppedandembryomaturationwasinitiatedbyculturingembryogenictissuefor4weeksonsolidOMSM6mediumwithoutcharcoalunderalightintensityof70lmolm-2s-1.
Embryosweredesiccatedin85%relativehumidityandgerminatedonB5mediumwith3%sucroseand0.
6%Phytagaror0.
2%Gelrite,pH5.
8.
Plantswithtwotrifoliateleavesweretransplantedtosoilandgrownundera16-hphotoperiodpriortotransfertoa12-hphotoperiodforseedformation.
Roottissueswereharvestedfrom1-weekoldseedlings.
Leafandstemtissueswerefrom3-weeksoldseedlings.
Flowerswereharvested2daysafterfullanthesis.
Developingseedsandpodswereharvestedfromdifferentstagesbetween6and24dayspostanthesis(dpa)at3daysintervals.
TostudytheexpressionofGmBBM1genesindevelopingseedstissuesfromseedcoat,embryoandpodswerecollectedseparatelyandRNAwasextractedusingtheRNeasyKit(Qiagen,Canada).
Tissuesfromsoybeanroots,leaves,stemsandowerswerealsocollected,homoge-nizedinliquidnitrogenandtotalRNAwasextractedusingTrizol(Invitrogen,Canada).
RT-PCRProbesgeneratedbyRT-PCRwereusedforlibraryscreeningandexpressionproling.
Primersspecictoeachofthethreesoybeangenes,GmBBM1,GmPLT2andGmAIL5,areshowninSupplementaryTable1.
TheSoy-Tub2genewasusedaspositivecontrol.
PCRconditionsforGmPLT2andGmAIL5wereasfollows:3minat95°C;30cycles[1min95°C,1minspecicannealingtemperature(SupplementaryTable1),1min72°C]and5minexten-sionat72°C.
PCRconditionsforGmBBM1andSoyTub2wereasfollows:31minat50°C,15minat95°C;30cycles(30s94°C,30sspecicannealingtemperature(Supple-mentaryTable1),30s72°C)and2minextensionat72°C.
LibraryconstructionTotalRNA(2mg/gfreshtissue)wasextractedfromculturedembryogenicsoybeansuspensionculturesandmRNA(0.
2%oftotalRNA)waspuriedusingtheQuikmRNAIsolationKit(Stratagene/VWR,Canada).
AcDNAlibrarywascon-structedusingLambda-ZAPIIcDNAsynthesiskit(Strata-gene/VWR,Canada).
Phages(1.
69106pfu)wereplatedaccordingtothemanufacturer'sinstructions.
TheampliedcDNAlibrarytitrewas2.
59109pfu.
ThecDNAlibraryphageplaqueswerehybridizedusingaprobecontainingtheAP2domainandlinkerusingprimersdesignedfromthealignmentofESTAW200688codingforasoybeanAP2proteinandBnBBM1fromBrassicanapusasdescribedinResults.
cDNAwereexcisedfromthelambda-ZAPIIvectorsfollowingtheconversionofthelambda-ZAPcloneplasmidspBluescriptSKDNA.
SequencesweredeterminedwiththeABIPRISMTMdyeterminatorcyclesequencingkit(PEAppliedBiosystems).
DatawasanalyzedusingDNAsis(HitachiCorporation2003)GenejumperprimerinsertionkitforsequencingversionB(Invitrogen,Canada)wasusedinordertobeabletosequencethefull-lengthclones.
ArabidopsistransformationandassessmentPlanttransformationswerecarriedoutaccordingtoCloughandBent(1998).
GmBBM1wasclonedintothepBINPLUSvectorandexpressionwasdrivenbythedouble35SPlantMolBiol(2010)74:313–326315123promoterandalfalfamosaicvirus(AMV)translationalenhancerasdescribedinBoutilieretal.
(2002).
Selectionwasperformedon50mg/lkanamycin.
Thousandsofseedswerescreeneduntilacollectionofapproximately35lineswithsomaticembryoswererecovered.
Ofthese,sixrep-resentativelineswerestudiedingreaterdetailoversuc-cessivegenerations.
BnBBM1mutantsweregeneratedbyinversePCRreactionsontheBnBBM1PUC19plasmid,usingExpandLongTemplatePCRSystem(Roche)andsyntheticprimerswith10-bpsequenceincludingaBamHIorXhoIsitesintheir50-endsthatreplacestheoriginalsequences.
ThefollowingprimerswereusedforeuANT2(N1BamHIF50-CGGGATCCCTGAGAAATCAACCCGTGGATAATG-30andN1BamHIR50-CTGGATCCGCCACCACCACCGTCTCCTCCTC-30)andforbbm-1(N2XhoIF50-CGGCTCGAGCCTTATGAACAAAATCACCATCG-30andN2XhoIR50-CCGCTCGAGGGTGGAAGTATTTGAAAGAAAT-30).
TheseconstructswereobtainedbyKpnI/SalIdigestionandclonedintothebinaryvectorpCAMBIA1300,alongwiththewild-typeBnBBM1geneasthecontrol.
Theywereintro-ducedintoAgrobacteriumtumefaciensGV3101forplanttransformationandselectionwasperformedwith30mg/Lhygromycin.
Approximately,3,000–4,000seedsintotalwerescreenedintriplicateexperimentstoassessthephe-notypesofthetransgeniclines.
Forthedomainswapanalysis,thePLT2,AIL5,andAIL7genesfromArabidopsisweresynthesizedwiththeBnBBM1bbm-1motif(SLGLSMIKTWLRNQP)asshowninFig.
2AbyGenScriptCorporation(Piscataway,NJ).
TheywereclonedintopCAMBIA1300undercontrolofthedouble35Spromoter.
Asabovetheconstructswereintro-ducedintoAgrobacteriumtumefaciensGV3103forplanttransformationandscreened.
Seedsfromeachtransgenicseedlineweresterilizedinasolutionof70%ethanolfor30sfollowedbyasolutionof25%javexplus0.
05%Tritonfor20–30min.
Theseedswereshakenfortheduration.
Theywerethenrinsed4timeswithsteriledistilledwaterandplacedina0.
1%solutionofaga-roseat4°Covernight.
Thenextday,theseedswereplatedonMSB5media(SigmaChemicalCo)with3%sucroseand0.
6%agarosealongwithselectableagents:50mg/lkana-mycinforpBINPLUSor30mg/lhygromycinforpCAM-BIA1300.
Seedlingswerethensurveyedforphenotypeovera2–3weekperiod.
At3–4weeksgrowththeyweretrans-ferredtosoilandbroughttooweringstage.
PhylogeneticanalysisSequencesofdoubleAP2domainproteinssimilartoAtBBMwereminedfromGenBank(NationalInstitutesofHealthgeneticsequencedatabaseatNationalCenterforBiotechnologyInformation)usingtheEntrezbrowser(availableat:http://www.
ncbi.
nlm.
nih.
gov/)andfromtheTIGRGeneIndices.
Proteinsthatwerenotfull-lengthwerenotincludedasweintendedtoeventuallyassesstheN-terminalandtheC-terminalsequencesalso.
ToidentifythosesequencesbelongingtotheeuANTsubgroup,sequenceswerescreenedforthe10aminoacidinsertionintherstAP2domaincharacteristicofthisgroupaswellasapredomainregionlongerthan127aminoacids,asdescribedbyKimetal.
(2006).
WRI1fromthebasa-lANTsubgroup,aseparateANTlineage,wasincludedasanoutgroup.
TheminedaminoacidsequenceswerealignedusingCLUSTALW(Thompsonetal.
1994)andfurtherimprovedbyvisualexaminationandeditingusingGeneDocVersion2.
6.
002(NicholasandNicholas1997).
TheAP2domainwasusedforalignmentasthefulllengthsequencesweretoodivergenttosuccessfullyalignallthesequencesretrievedfromGeneBankandTIGRGeneIndicesdatabase.
WelimitedtheboundariesofthedoubleAP2domaintothesequencesstarting5aminoacidsupstreamandapproximately10aminoacidsdownstreamoftheconserveddoubleAP2domains.
Intotal,53sequences(SupplementaryTable2)wereanalyzed.
Thealignedsequenceswererstsubjectedtoatoolfortheselectionofthebesttmodelfromamong112modelsofproteinevolution,usingProtTest(Abascaletal.
2005)version2.
2.
Afterthelikelihoodstatisticswerecompletedthreestatisticalframeworkswereselectedconsecutivelyoneatatimetodeterminewhichcandidatemodeltsthedatabest.
Althoughatreeusingthemodel-averagedesti-mateoftheparameterswasobtainedintheanalysisandexamined,wecarriedaseparateMaximumLikelihood(ML)phylogeneticanalysisusingthePROMLprogram(PROteinMaximumLikelihoodprogram)inthePHYLIPpackage(Felsenstein2008)withtheselectedevolutionarymodelalongwithitsspecicparameters,previouslyobtainedfromProtTest.
Intheanalysis100bootstraptreeswereproducedfromwhichamajorityruletreewascom-putedtoobtainbootstrapsupportforthebranchesontheMLtreeandastrictconsensustreewasalsocomputedforexamination.
Anothersimilarbootstrapanalysiswascar-riedwithMEGA4(NeiandKumar2000)with1,000repeats.
ConservedmotifswereidentiedthroughacombinationofClustalWalignment,MEMEversion4.
0.
0(BaileyandElkan1994),andBlockMaker(Henikoffetal.
1995).
TheEukaryoticLinearMotifresourceforFunctionalSitesinProtein(http://elm.
eu.
org/links.
html)wasusedtopredictmotiffunction.
316PlantMolBiol(2010)74:313–326123ResultsAP2familygenesclonedfromsoybeansomaticembryosSoybeancDNAswithsequencessimilartoBnBBM1,wereisolatedfromimmaturesomaticembryosofsoybean,G.
maxcvMerrilgenotypeX5(SimmondsandDonaldson2000).
TheprobeusedtoscreenthecDNAlibrarywasstrategicallydesignedusingPCRprimers(SupplementaryTable1)targetedtosequencesidentiedfromalignmentsofBnBBM1andapartialsoybeanputativeAP2proteinESTAW200688fromacDNAlibraryconstructedfromcotyledonsof3-and7-day-oldseedlingsofcultivarWilliams(http://www.
ncbi.
nlm.
nih.
gov/entrez/viewer.
fcgidb=nucest&val=6481417)(ShoemakerR,ThePublicSoybeanESTProject)(BiogeneticsServices,SD,USA)sequences.
Twoclones,GmBBM1(2.
5kb,707aa,GenBankAccessionHM775856)andGmAIL5(2.
3kb,558aa,GenBankAccessionHM775857),wereidentiedandiso-lated.
AnothercDNAclone(2Kb,560aa),GmPLT2,wasrstidentiedastwo50-trunctedcDNAclonesthatlaterappearedasafulllengthclone(EU677381).
AllpossessedhighlevelsofsequencesimilarityinthedoubleAP2domainregion(97%)whichdecreasedintheN-andC-terminalregions(SupplementaryFig.
1A).
ThesoybeanGmBBM1sequencewassimilartothoseofthecruciferousgenes,BnBBM1andAtBBM,withmorethan91%simi-larityinthedoubleAP2domains(SupplementaryFig.
1B).
ThisdecreasedintheN-andC-terminalsequencesto48and30%respectively(SupplementaryFig.
1B).
PhylogeneticanalysisofGmBBM1,GmAIL5andGmPLT1ToexaminetherelationshipsofthethreesoybeangeneswithotherAP2familymembersallavailablegenesinTheAra-bidopsisInformationResource(http://www.
arabidopsis.
org/),GenBank,DFCI(DanaFarberCancerInstitute)GeneIndex(http://compbio.
dfci.
harvard.
edu/tgi/;formerlyTIGRgeneindex),andtheTIGRRiceGenomeAnnotationProjectDatabase(http://rice.
plantbiology.
msu.
edu)atthetimeofthestudywithsequencesimilaritytoBBMgeneswereretrievedusingthedoubleAP2domainregionoftheAtBBMsequence.
Onlyfull-lengthsequencesandthosewithatleast60%similarityinthedoubleAP2domainwereretained.
SequencesbelongingtotheeuANTsubgrouparecharacterizedbythepresenceofa10aminoacidinsertintherstAP2domainregionandarelativelylongpredomainregion(Kimetal.
2006).
Thesetwocharacteristicswereusedtoselect49sequencesbelongingtotheeuANTsubgroup.
WRI1,belongingtothebasalANTsubgroup,wasincludedasanoutgroup.
Atotalof53sequences(SupplementaryTable2)wereincludedinthephylogeneticanalysis.
ThealignmentwascarriedoutusingtheregionspanningthetwoAP2domains,includingthelinkerregion,starting5residuesupstreamoftherstconservedAP2domainandnishingapproximately10aminoacidsdownstreamofthesecondAP2domain.
TheevolutionarymodelthatbesttthisdatawastheJTTGmodel(i.
e.
theJones-Taylor-Thorntonmodel)underthethreestatisticalframeworksanalyzed,i.
e.
AIC,AICcandBIC,anditsparameterG=0.
558,i.
e.
gammashapewith4ratecategories.
Figure1showstherelationshipofthethreesoybeangenestotheeuANTsubgroupbasedontheAP2-linker-AP2sequences.
GmBBM1isclosesttotheotherlegumeBBMgene,MtBBM,fromMedicagotruncatulaandalsoclusterscloselywiththeBBMgenesfromtheBrassicaceaandPoaceaefamilies.
GmAIL5isgroupedwiththeoilpalmEgAp2-1geneandisclosetoAIL5fromArabidopsis.
GmPLT2isgroupedwiththeArabidopsisPLT1andPLT2genesalongwithagenefromVitisvinifera,allofwhichformthelargerPLT-likegroupingwithArabidopsisAIL6andAIL7andsequencesfromrice,Brassicarapa,andanothersequencefromVitisvinifera.
MotifcompositionandidenticationofaBBM-specicmotifTofurtherdistinguishthegenesintheBBM-likegroupingfromothereuANTgenes,themorevariableN-terminalandC-terminalproteinsequenceswerealignedtosearchforconservedmotifs(seeSupplementaryFig.
1C).
IntheN-terminalsequences,vemotifswereidentiedinmostmembersoftheBBM-likegrouping.
ThreeofthesemotifswerepreviouslyidentiedintheeuANTlineage(euANT2,3,and4;Kimetal.
2006)andwerefoundthroughoutmostofthesequencesanalyzed(Fig.
2A).
Thefourthmotif(bbm-1)wasspecictotheBBM-likegenesandabsentfromtheclosely-relatedPLT-likegenes(Fig.
2A).
ThismotifwasweaklyconservedinsomeoftheANT-likegenes,particularlyGbANTL1,PtANTL1,andCAO45633.
ItwasalsofoundinOs01g67410,whichwasnotassociatedwithanyoftheidentiedgroupings.
Thefthmotif(euANT5)wasalsoidentiedinthemajorityofthemembersoftheBBM-likegroupingbutwasabsentintheBBM-likegenesfromCenchrusciliarisandPennisetumsquamulatum(Fig.
2A).
ThismotifisalsoconservedintheANT-likegenesaswellastheAIL1-likegenes,andrepre-sentativescontainingthismotifcanalsobefoundamongstthePLT-likegenes,althoughitiscompletelyabsentfromtheAIL5-likegenes.
Interestinglythebbm-1andeuANT5motifsboresomesimilarity,bothincludingthesequenceLSM.
IntheC-terminalsequencesveothermotifswereidentied(Fig.
2B).
Amotif(euANT6)wasidentiedinallPlantMolBiol(2010)74:313–326317123oftheBBM-likegenesimmediatelyfollowingthesecondAP2domain.
ItwasidentiedinrepresentativesfromallofthemajoreuANTlineages,althoughthedegreeofcon-servationvariedconsiderably(Fig.
2B).
Thismotifcon-tainsanumberofLysandArgresidues,suggestingthatitmayatleastpartiallyfunctionasanuclearlocalizationsignal,ashasbeensuggestedforthePLTgenes(Aidaetal.
2004).
ThefouradditionalmotifsintheC-terminalsequencesoftheBBM-likegeneswereconservedwithalowdegreeofspecicity.
Thebbm-2motifwasonlyweaklyconservedintheArabidopsisandB.
napusBBMgenesandbbm-3wascompletelyabsent.
SomeofthesemotifswereweaklyconservedinsequencesoutsideoftheBBM-likegroupingparticularlybbm-3andbbm-4insomeofthePLT-likesequences.
Aswiththebbm-1motifs,Os01g67410alsocontainedthebbm-2andbbm-4motifs,althoughthebbm-3motifwasonlyweaklyconservedandthebbm-5motifwasabsent.
Severalofthemotifs,includingeuANT2,bbm-1,euANT5,andeuANT6,containconsensussequencesforphosphorylation.
euANT5alsocontainsaconsensussequenceforsumoylation.
TheeuANT2motifalsocon-formstoaTRFHdomaindockingmotifandaWWligandmotif.
ExpressionproleofGmBBM1,GmAIL5andGmPLT1Insoybeanplants,theexpressionofGmBBM1paralleledthatofAtBBMinArabidopsis(Boutilieretal.
2002).
Itwasselectivelyexpressedinsoybeanembryosandyoungroots(Fig.
3).
GmAIL5alsopossessedthesameexpressionpat-ternasAIL5inArabidopsis(Nole-Wilsonetal.
2005).
Itwasexpressedinallofthesoybeanorgansundergoinggrowthanddevelopmentthatwereexamined(Fig.
3).
GmPLT2hadthesameexpressionpatternasPLT1andPLT2inArabidopsis(Aidaetal.
2004)andM.
truncatula(Iminetal.
2007;Holmesetal.
2008).
Itwasexpressedpredominantlyinsoybeanrootsandtoalesserextentinsoybeanembryos(Fig.
3).
Thedatasupportedtheassign-mentofGmBBM1asaBBMhomologue,GmAIL5asanFig.
1MaximumlikelihoodtreeoftheeuANTsubgroupofthedoubleAP2family318PlantMolBiol(2010)74:313–326123Fig.
2MotifsintheN-terminal(A)orC-terminal(B)sequencesoftheBBM-likegenesandtheirconservationintheeuANTsubgroupoftheAP2family.
Residuesthatmatchtheconsensussequenceareinred.
Sequencesaregroupedintocladesasindicatedinthephylogenetictree.
ThesequencesoftheeuANT2andbbm-1motifsintheBnBBM1sequencethatweredeletedtotesttheirfunctionareunderlined.
Forthedomainswapanalysis,thesiteofinsertionofthebbm-1motiffromBnBBM1(SLGLSMIKTWLRNQP)inPLT2,AIL5,andAIL7areindicatedwitharrowheadsPlantMolBiol(2010)74:313–326319123AIL5homologueandGmPLT2asaPLThomologuebasedonthegenephylogeny,conservationoftheN-terminalandC-terminalsequencesandtheexpressionpatterns.
PhenotypesgeneratedbyBBM-likegenesintransgenicArabidopsisAcomparisonoftheembryogenicphenotypeandpleio-tropiceffectsconferredbyGmBBM1andBnBBM1wascarriedoutintransgenicArabidopsisusingthepBINPLUSvectorsystemusedpreviouslyforectopicoverexpressionofBnBBM1(Boutilieretal.
2002)(Fig.
4).
Weconrmedpreviousdescriptions(Boutilieretal.
2002)thatshowedBnBBM1expressioninArabidopsisinducedsomaticembryosandcotyledon-likestructuresonpost-germinationorgansoftransgenicseedlingsaswellasauniquesetofpleiotropiceffectsonboththevegetativeandreproductiveorgans(Table1,BnBBM1lines1–3).
Approximately35independenttransgenicArabidopsislinesexpressingGmBBM1embryosonthecotyledonswereexaminedandsixrepresentativelineswerefollowedintotheT2genera-tions(Fig.
4A–C).
Table1showthedatafor3oftheGmBBM1lines.
Somaticembryogenesisgenerallyoccur-redatalowerfrequencythanwithBnBBM1(Table1).
Thehighestfrequencywas28%intheGmBBM1line1,fol-lowedby17%intheGmBBM1line2and5%intheGmBBM1line3(n[100foreachline).
Embryosalsoemergedfromtheshootapicalmeristem(Fig.
4D)andhypocotyls(Fig.
4E).
PleiotropiceffectsondevelopmentthatweretypicalofBnBBM1overexpressionwereagainevidentwithGmBBM1overexpression.
Theseincludedshortroots(80%penetrance,GmBBM1line3;n=100),Fig.
3AccumulationofGmBBM1,GmAIL5,GmPLT1andtubulin(accessionnumberM21297)mRNAinsoybeancvMerrilgenotypeX5detectedbyRT-PCR.
Rootsat1week(R),stemat3weeks(S),leafat3weeks(L),owersat2dpa(F),seedpodat21dpa(SP),seedcoatsat21dpa(SC),embryosat21dpa(E)Fig.
4PhenotypesinducedbyexpressionofGmBBM1intransgenicArabidopsisseedlings.
Somaticembryosdifferentiatingfromthetipsofcotyledons(A,B,C).
Ectopicrootsdifferentiatingfromtheembryogenictissues(A,C).
Somaticembryosdifferentiatingfromtheshootapicalmeristem(D)andhypocotyls(E).
Alterationsinseedlingdevelopmentcharacterizedbyshortroots,swollenhypocotylsandelongatedcotyledons(E,F).
Un-transformedcontrolseedlingsforcomparison(G)320PlantMolBiol(2010)74:313–326123shorthypocotyls(greaterthan80%penetranceinallGmBBM1linestested),elongatedcotyledons(lowpene-trancephenotype\25%inallGmBBM1lines)onseed-lings(Fig.
4E,F).
Laterindevelopmentarangeofpleiotropiceffectswerenotedincludingalteredleafmor-phologiesandthickenedoralstems.
Increasednumbersofinorescenceswerealsonoted(datanotshown).
Duringthereproductivephaseofgrowth,GmBBM1andBnBBM1bothinduceddelayed-oweringtime;increasedoralorgansize;thickenedandincreasednumbersofinores-cences;decreasedsiliquesize;anddecreasedseedpro-duction(Table1).
AlthoughmanyofthephenotypesweresimilarforBnBBM1andGmBBM1someinterestingdifferenceswereobserved.
Forexample,inGmBBM1transgeniclinesectopicrootsoccassionallydevelopedoncotyledonsalongwithembryos(Fig.
4A,C),afeaturethatwasnotreportedwithBnBBM1(Boutilieretal.
2002).
Furthermore,elon-gatedrootswereseenonplantletsoftheGmBBM1lineincontrasttotheshort-rootphenotypeobservedinBnBBM1plants(Table1).
Functionofthebbm-1motifinembryogenesisThesignicanceofthebbm-1sequencemotiftoBnBBM1functionwasexaminedbycreatinga9-aminoaciddeletionmutationintheBnBBM1gene(Fig.
2A)andassessingfunctionalityintransgenicArabidopsis(Fig.
5).
Forcom-parison,adeletionofmotifeuANT2(Fig.
2A)wasalsocreatedsinglyandtogetherwithadeletionofbbm-1.
Theinductionofsomaticembryosoncotyledonsbywild-typeBnBBM1wasrstconrmedin22transgeniclinesascontrolsusingthepCAMBIA1300vector(Fig.
5C).
Shootssubsequentlydevelopedinculture(Fig.
5D)andplantletsregeneratedasdescribedpreviouslybyBoutilieretal.
2002(datanotshown).
SomaticembryogenesiswasnoteliminatedbydeletionofeuANT2(Fig.
5G,H)oreuANT2andbbm-1together(Fig.
5I,J).
However,themutantphe-notypesdifferedinthattheembryo-likestructurestendedtoproliferateandshootsdidnotemergeovertime(Fig.
5H,J)aswithwild-typeBnBBM1(Fig.
5D).
Thisphenotypewasconrmedin14and25transgeniclines,respectivelyandwasinstrikingcontrasttothephenotypegeneratedbydeletionofthebbm-1motifalonewhichwascharacterizedbythecompletelossofsomaticembryogenesisoncotyle-donsaswellastheaccompanyingpleiotropiceffectsonseedlings(Fig.
5E,F).
Pleiotropiceffectsassociatedwiththismutationemergedatlaterstagesofplantdevelopment.
SomeweresimilartotheclassIIpleiotropiceffectsdescribedforBnBBM1byBoutilieretal.
(2002).
Forexample,owerswithshortsepalsandpetalsrelativetocarpelswerefrequentlyobserved(Fig.
6A)comparedwithwildtypeowers(Fig.
6B).
Thechangesinleafmorphol-ogydescribedforBnBBM1(Boutilieretal.
2002)wereinfrequentlyobserved;however,serratedmarginswereoccasionallyobserved(Fig.
6C)inplantsexpressingthedeletionmutantbutnotwildtypeplants(Fig.
6D).
Thebbm-1motifwasinsertedintoArabidopsisPLT2,AIL5andAIL7(Fig.
2A)todetermineifothereuANTmemberspossessedthecomplementaryelementsneededforsomaticembryogenesisintransgenicArabidopsis.
AIL7bbm-1andAIL5bbm-1failedtogenerateectopicembryosinexperimentsparalleltothoseabove(datanotshown).
TheectopicPLT2phenotypewaspre-viouslycharacterizedbytheproliferationofectopicrootsandroothairsongerminatingembryosorseedlings(Aidaetal.
2004).
PLT2bbm-1generatedarangeofpheno-typesinvedifferenttransgeniclines.
Rapidandstrongtissueproliferationfollowedbycopiousembryodifferen-tiationwasobservedin4ofthe5lines(Fig.
7A,B).
ThisphenotyperesembledtheBBMphenotype(BenScheresandcolleagues,personalcommunication;Fig.
7).
SmallerTable1PhenotypesofindividualtransgeniclinesexpressingBnBBM1andGmBBM1constitutivelyintransgenicArabidopsisDescriptionofphenotypeTransgenicBnBBM1linesTransgenicGmBBM1linesLine1Line2Line3Line1Line2Line3SomaticembryoformationoncotyledonsShortroots,brousrootsElongatedrootsThick,shorthypocotylsElongatedcotyledonsDelayedoweringtimeReducedseedsetIncreasedoralorgansizeThephenotypesinboldattheseedlingstageofdevelopmentandusedfortheinitialscreeningoflines.
Thefrequencyofoccurrenceofthephenotypewasrecordedforeachlineusingatleast100plantlets([0\25%),(25–50%50–75%75–100%)PlantMolBiol(2010)74:313–326321123undenedgreentissueshadalsobeendescribedaspartofthePLT2ectopicphenotype(Aidaetal.
2004)In2ofthelines,bothBBMandPLT2phenotypeswereexpressedsimultaneously.
Intheselinesembryodifferentiationwaslessprolicandrootdifferentiationwasveryabundant(Fig.
7C,D).
Fig.
5Phenotypeofun-transformedArabidopsisseedlingsat14days(A)and21–28days(B)postgerminationcomparedwiththephenotypeof14dayand21–28daytransgenicArabidopsisectopicallyexpressingwild-typeBnBBM1(C,Drespectively),theBnBBM1deletionmutantwithoutbbm-1(E,Frespectively),theBnBBM1deletionmutantwithouteuANT2(G,Hrespectively),theBnBBM1deletionmutantwithouteuANT2andbbm-1(I,Jrespectively)322PlantMolBiol(2010)74:313–326123Fig.
6PleiotropiceffectsontransgenicArabidopsisower(A)andleaf(C)morphologiesgeneratedbytheBnBBM1deletionmutantthatlacksthebbm-1motifcomparedwithwildtypeowers(B)andleaf(D)Fig.
7PhenotypesoftransgenicArabidopsisseedlings,inwhichthebbm-1motiffromBnBBM1wasinsertedintothecorrespondingpositionofthePLT2gene,at11days(A,B),13days(C)and17days(D)postgerminationPlantMolBiol(2010)74:313–326323123DiscussionThephenotypesassociatedwiththeAP2/ANTfamilyfrequentlyinvolveundifferentiatedcellproliferationanddifferentiationofstemcellnicheswithinmeristems(Nole-Wilsonetal.
2005).
Recently,aroleinembryogenesishasalsoemerged.
MostoftheinformationhasbeengeneratedbytheinductionofsomaticembryogenesisbyectopicoverexpressionofBnBBM1inArabidopsisandBnapus(Boutilieretal.
2002;Passarinhoetal.
2008).
GmBBM1wasshownheretoinducesimilardevelopmentaleventsinArabidopsis.
Comparedwithmeristematiccelltypes,thedifferentiationofembryonalstemcellsfromsomaticcellsispoorlyunderstood.
Studieshaveshownthatsomaticembryogenesismaybeactivatedthroughanumberofdifferentpathwaysandbyanumberofdifferentgenes(Passarinhoetal.
2008;reviewedbyVerdeiletal.
2007).
Inthisstudy,wehaveidentiedspecicfeaturesoftheBBM-likehomologuesthatareresponsiblefortheinductionofembryogenesisandthatmayseparatetheBBM-likegenesfromcloselyrelatedmembersoftheAP2/ANTfamilythatfunctioninvegetativedevelopmentyetactredundantlyinembryodevelopment.
TounderstandhowBBM-likegenesfunctionintheconversionofsomaticcellstoembryonalcellsandthedevelopmentoftheresultingembryoitmaybeimportanttounderstandtherelationshipsamongtheAP2/ANTfamilymembers.
ThemembersoftheeuANTlineage,suchastheBBM-like,PLT-likeandAIL5-likegenes,areverycloselyrelatedphylogeneticallyandstructurally.
Ouranalysishasrevealeddomainconservationamongthembutalsodomainspecicity.
Forexample,thebbm-1motifwasspecictoalloftheBBM-likegenes.
Thedifferentmembersareknowntohavespecicrolesindevelopmentbutevidenceforredundancyisalsoemerging.
Forexample,themutantphenotypesofbbmandplt2haveshownthatAtBBMandPLT2functionasredundantpartnersinearlyArabidopsisembryogenesis(Galinhaetal.
2007).
AtBBM,PLT1,PLT2andPLT3/AIL6areexpressedinrootprimordiawheretheyfunctionasredundantpartnersinrootstemcelldifferen-tiationinembryosandrootdifferentiation(Aidaetal.
2004;Galinhaetal.
2007).
Thedemonstrationthattheadditionofthebbm-1motiftoPLT2enhancedthecapacitytoinducesomaticembryogenesisconrmedthecloserelationshipbetweenthesegenesandmayhaverevealedakeystructuralfeatureofBBMimportantforitsabilitytofunctioninembryogenesis.
ThePLT2ectopicphenotypewasnotlostwhentheBnBBM1ectopicphenotypewasexpressedbutthestrengthofthePLT2andBnBBM1ectopicphenotypeswereinverselyrelatedindicatingthattheembryogenicandrootmeristematicpathwayscouldbecompetinginthesetransgeniclines.
HowAIL5functions,particularlyinembryogenesis,isunclear.
AIL5appearstobeinvolvedincellproliferationactivitiesinmanyorgansandmaygenerateenlargedorgans,forexamplelargeowers,whenectopicallyexpressed(Nole-Wilsonetal.
2005).
ThisphenotypeissimilartothatofANT,whichgenerateslargeowers(Nole-Wilsonetal.
2005;MizukamiandFischer2000)andAP2whichgenerateslargeembryos(Ohtoetal.
2005)whenectopicallyexpressed.
SinceAIL5mutantshavenoapparentphenotypeitisbelievedthatitactsredundantlywithotherrelatives(Nole-Wilsonetal.
2005).
AIL5andBBMdoublemutantsappeartolackdefectsinembryoorrootmeristemdevelopmentthereforeadditionalunidentiedpartnersmayexistiftheyactinthesamepathways(Tsuwamotoetal.
2010).
Thendingthatectopicover-expressionofAIL5yieldedphenotypessimilartoBBMover-expression,includingsomaticembryogenesis(Tsuwamotoetal.
2010),indicatesthatitcouldbeinvolvedintheproliferationofthetotipotentsomaticcells.
TheembryonicstructuresthatemergedexhibitelevatedexpressionofAGL15,LEC1,LEC2,FUS3andBBM(Tsuwamotoetal.
2010)allofwhichhavethecapacitytoinducesomaticembryogenesiswhenectopicallyexpressed.
Interesting,theoilpalmAIL5-likegene,EgAp2-1,isalsoexpressedinproliferatingtis-sues,especiallyembryonictissuesbutinducescallusgrowthandshootorganogenesisratherthanembryosinArabidopsis(Morcilloetal.
2007).
BnBBM1alsoinducesshootorganogenesisintobaccoratherthanembryogenesisrevealingthatBBM-likegeneshavethecapacitytoinducebothshootmeristemactivityaswellasembryogenesisdependingonthegeneticandcellularenvironmentoftheproliferatingcells(Srinivasanetal.
2007).
UnlikePLT2,theadditionofthebbm-1motiftoAIL5didnotgeneratethecapacitytoinduceembryogenesisintransgenicArabidopsisinthisstudy.
WhetherthisalterationdisruptedthecapacityofAIL5togeneratesomaticembryosasdescribedbyTsuwamotoetal.
(2010)isunknownandpointstocomplexrolesforAIL5inplantdevelopment.
Conservedsequencemotifshavebeenrecognizedamongsoybean,riceandArabidopsisAP2/ANTgenesandsoybean-specicmotifsareamongthem(Zhangetal.
2008).
Previousstudieshavesuggestedthatwithinsub-familiesconservedsequencemotifsareabundantandlikelyreectsharedfunctions(Nakanoetal.
2006).
Unlikesomeofthewell-characterizedmotifsoftheERFfamily,sequencemotifswithintheAP2/ANTfamilyhavenotyetbeenassignedspecicrolesinthetranscriptionalregula-tionofdevelopmentalprocesses(Nakanoetal.
2006;Zhangetal.
2008).
ThecharacterizationoftheGmBBM1genehereallowedustoaligntheBBM-likegenesfromdivergentspecieswhichareknowntogenerateectopicembryosinArabidopsisandthusprovideafunctionaldata324PlantMolBiol(2010)74:313–326123setthatwasusedtoidentify10conservedsequencemotifsoutsideoftheAP2-linker-AP2region.
AmongtheBBM-like,PLT-likeandAIL5-likegenesthemotifcompositionwasverysimilar,however,thebbm-1motifwasthemostspecictotheBBM-likegenesintheN-terminalregion.
IntheC-terminalregionsequenceswithalowerdegreeofconservationamongtheBBM-likegenesexist.
Ofthesebbm-2,bbm-3andbbm-4arepoorlyconservedinthecruciferousgenesandthusunlikelytobeessentialforembryogenesis.
Themotif,bbm-5,isalsopoorlyconservedrelativetobbm-1.
Bothdeletionanddomainswapanalysesrevealedthatthebbm-1motifwasneededforBnBBM1toinducesomaticembryogenesisinArabidopsis.
Furtherexaminationrevealedthatbbm-1activityisintimatelylinkedtotheactivitiesofothermotifswithinthesetranscriptionfactors,especiallyeuANT2,whichisfoundinalmostallothergenesintheeuANTlineageincludingPLT2andAIL5.
DeletionofeuANT2singlyoreuANT2andbbm-1simultaneouslypre-ventedthesomaticembryosfromgeneratingshootsevenafterprolongedtimesinculture.
Amongthemanypossibleexplanationsforbbm-1functionisthepossibilitythattheeuANT2motiffunctionsinvegetativepathwaysofdevel-opmentandthatbbm-1actstorestrainthevegetativeactivityofBnBBM1.
Ifcorrect,thenamechanismmustexisttorelievetherestraintsoneuANT2inBnBBM1duringthelaterstagesofembryodevelopmentwhentheshootandrootmeristemsdifferentiateanddevelop.
Furthermore,ifasingledomain,bbm-1,canseparatetheectopicphenotypesofredundantpartnerssuchasBBMandPLT2thenitwouldseempossiblethatthesharedmotifscouldprovidethebasisforfunctionalredundancy.
AhierarchicalprocessmustexistinthecellularenvironmenttorecruitandcoordinatethedifferentAP2/ANTmemberstothedifferentdevelopmentalpathwaysinanorderlymanner.
Asboththebbm-1andeuANT2motifshavetheconsensussequenceforphosp-orylationpost-translationalprocessesmaybeinvolvedintheregulationofsuchinteractions.
ForPLT-likegenes,expressiongradientsappeartoplayanimportantinthetransitionfromrootmeristeminitiationtorootdevelopment(Galinhaetal.
2007).
AnygeneralmodelfortheroleofAP2/ANTtranscriptionfactorsinsomaticembryogenesisshouldconsidertheobservationsthatBBM-likegenesmayplaydistinctrolesatdifferentstagesofdevelopment;interactwithredundantpartners;andinducedifferentpathwaysofdevelopmentdependingonthegeneticandcellularenvironment.
Thebbm-1motifofBBM-likegenesappearstoplayanearlyroleintheconversionofsomaticcellstoneoplasticembryonalcells.
Thismayoccurbysuppressingtheactivityofothermotifs,suchaseuANT2,whichfunctioninvegetativecelldifferentiation.
ThesharedmotifsamongtheAP2/ANTmembersmayprovideamechanismforBBMtorecruittheactivitiesofmemberswithmeristematicactivitiesessentialforthedevelopmentoftheembryoonceembryogenesisisestablished.
Asnewmembersarerecruitedtothedevel-opingembryodilutionofthebbm-1motifmayreleasethecellsfromtheneoplasticstateandallowthemeristemstodevelop.
SustainedectopicexpressionofBBMgenesdur-ingplantmaturationresultsinarangeofpleiotropiceffects.
ThismayoccurthroughsimilarunintendedinteractionswithAP2/ANTproteinsthroughsharedmotifsthatdisrupttheirnormalpatternsofactivity.
ThedifferentpleiotropiceffectsobservedforGmBBM1andBnBBM1mayrevealdifferencesintheefcienciesofinteractionsresultingfromsequencedivergence.
IfBBM-likegenesfunctionbyuti-lizingredundancieswithotherAP2/ANTtranscriptionfactorstorecruitdevelopmentalpathwaysneededforembryogenesisandembryodevelopmentinseedsthenstrictspatialcontrolofBBM-likegeneexpressionwouldbeessentialfornormalplantdevelopment.
AsshownhereandinallotherstudiesofBBM–likegeneexpression(Boutilieretal.
2002;Nole-Wilsonetal.
2005;Iminetal.
2007)expressioninseedsisunderstrictspatialanddevelopmentalregulationasaretheotherAP2/ANTfamilymembers(Fengetal.
2005).
AcknowledgmentsTheauthorsareverygratefultoLiWangforArabidopsistransformation,MichelleLopezforthetransgenicstud-ies,WayneStanglefortheconstruct,DainaSimmonds,SherylHubbardandSuquinZhengforthesoybeantissues,RejeanDesa-gagneandDavidGagnefortheirsupportandcooperationandBenScheresandhislabforexaminationofourdatapriortopublication.
TheresearchwasfundedbytheCanadianBiotechnologyStrategy.
OpenAccessThisarticleisdistributedunderthetermsoftheCreativeCommonsAttributionNoncommercialLicensewhichper-mitsanynoncommercialuse,distribution,andreproductioninanymedium,providedtheoriginalauthor(s)andsourcearecredited.
ReferencesAbascalF,ZardoyaR,Posada,D(2005)ProtTestselectionofbesttmodelsofproteinevolution.
Bioinformatics21:2104–2105.
http://darwin.
uvigo.
es/software/prottest.
htmlVersion2.
2acces-sed28Sept.
2008AidaM,BeisD,HeidstraR,WillemsenV,BliloumI,GalinhamC,NussaumemL,NohmY-S,AmasinomR,ScheresB(2004)ThePLETHORAgenesmediatepatterningofthearabidopsisrootstemcellniche.
Cell119:109–120BaileyTL,ElkanC(1994)Fittingamixturemodelbyexpectationmaximizationtodiscovermotifsinbiopolymers.
In:AltmanR,BrutlagD,KarpP,LathropR,SearlsD(eds)Proceedingsofthesecondinternationalconferenceonintelligentsystemsformolecularbiology.
AmericanAssociationforArticialIntelli-gencePress,MenloPark,CA,pp28–36BoutilierK,OffringaR,SharmaVK,KieftH,OuelletT,ZhangL,HattoriJ,LiuCM,vanLammerenAA,MikiBL,CustersJB,vanLookerenCampagneMM(2002)EctopicexpressionofBABYBOOMtriggersaconversionfromvegetativetoembryonicgrowth.
PlantCell14:1737–1749PlantMolBiol(2010)74:313–326325123CloughSJ,BentAF(1998)Floraldip:asimpliedmethodforAgrobacterium-mediatedtransformationofArabidopsisthali-ana.
PlantJ16:735–743ElliottRC,BetznerAS,HuttnerE,OakesMP,TuckerWQJ,GerentesD,PerezP,SmythDR(1996)AINTEGUMENTA,anAPET-ALA2-likegeneofarabidopsiswithpleiotropicrolesinovuledevelopmentandoralorgangrowth.
PlantCell8:155–168FelsensteinJ(2008)PHYLIP,PhylogenyInferencePackage,version3.
68.
Availableathttp://evolution.
gs.
washington.
edu/phylip.
html.
AccessedNovember2008FengJ-X,LiuD,PanY,GongW,MaL-G,LuoJ-C,DengXW,ZhuY-X(2005)AnannotationupdateviacDNAsequenceanalysisandcomprehensiveprolingofdevelopmental,hormonalorenvironmentalresponsivenessoftheArabidopsisAP2/EREBPtranscriptionfactorgenefamily.
PlantMolBiol59:853–868FinerJJ(1988)Apicalproliferationofembryogenictissueofsoybean[Glycinemax(L.
)Merrill].
PlantCellRep7:238–241FinerJJ,NagasawaA(1988)Developmentofanembryogenicsuspensioncultureofsoybean[Glycinemax(L.
)Merrill].
PlantCellTissueOrgCult15:125–136FloydSK,BowmanJL(2007)Theancestraldevelopmentaltoolkitoflandplants.
IntJPlantSci168:1–35GalinhaC,HofhuisH,LuijtenM,WillemsenV,BlilouI,HeidstraR,ScheresB(2007)PLETHORAproteinsasdose-dependentmasterregulatorsofArabidopsisrootdevelopment.
Nature449:1053–1056GongW,ShenY-P,MaL-G,PanY,DuY-L,WangD-H,YangJ-Y,HuL-D,LiuX-F,DongC-X,MaL,ChenY-H,YangX-Y,GaoY,ZhuD,TanX,MuJ-Y,ZhangD-B,LiuY-L,Dinesh-KumarSP,LiY,WangX-P,GuH-Y,QuL-J,BaiS-N,LuY-T,LiJ-Y,ZhaoJ-D,ZuoJ,HuangH,DengXW,ZhuY-X(2004)Genome-wideORFeomecloningandanalysisofarabidopsistranscriptionfactorgenes.
PlantPhysiol135:773–782HenikoffS,HenikoffJG,AlfordWJ,PietrokovskiS(1995)Auto-matedconstructionandgraphicalpresentationofproteinblocksfromunalignedsequences.
Gene163:GC17–GC26HolmesP,GoffardN,WeillerGF,RolfeBG,IminN(2008)TranscriptionalprolingofMedicagotruncatulameristematicrootcells.
BMCPlantBiol8:21IminN,NizamidinM,WuT,RolfeBG(2007)FactorsinvolvedinrootformationinMedicagotruncatula.
JExpBot58:439–451JofukuKD,BoerB,MontaguMV,OkamuroJK(1994)ControlofarabidopsisowerandseeddevelopmentbythehomeoticgeneAPETALA2.
PlantCell6:1211–1225KimS,SoltisPS,WallK,SoltisDE(2006)PhylogenyanddomainevolutionintheAPETALA2-likegenefamily.
MolBiolEvol23:107–120KlucherKM,ChowH,ReiserL,FischerRL(1996)TheAINTE-GUMENTAgeneofarabidopsisrequiredforovuleandfemalegametophytedevelopmentisrelatedtotheoralhomeoticgeneAPETALA2.
PlantCell8:137–153KrizekBA,ProstV,MaciasA(2000)AINTEGUMENTApromotespetalidentityandactsasanegativeregulatorofAGAMOUS.
PlantCell12:1357–1366MaesT(1999)TheinorescencearchitectureofPetuniahybridaismodiedbytheArabidopsisthalianaAp2gene.
DevGenet25:199–208MizukamiY,FischerRL(2000)Plantorgansizecontrol:AINTE-GUMENTAregulatesgrowthandcellnumbersduringorgano-genesis.
ProcNatlAcadSciUSA97:942–947MorcilloF,GallardA,PillotM,JouannicS,Aberlene-BertossiF,CollinM,VerdeilJL,TregearJW(2007)EgAP2–1,andAINTEGUMENTA-like(AIL)geneexpressedinmeristematicandproliferatingtissuesofembryosinoilpalm.
Planta226:1353–1362NakanoT,SuzukiK,FujimuraT,ShinshiH(2006)Genome-wideanalysisoftheERFgenefamilyinArabidopsisandrice.
PlantPhysiol140:411–432NeiM,KumarS(2000)MolecularEvolutionandPhylogenetics.
OxfordUniversityPress,NewYorkNicholasKB,NicholasHBJr(1997)GeneDoc:atoolforeditingandannotatingmultiplesequencealignments.
Distributedbytheauthors.
www.
psc.
edu/biomed/genedoc/AccessedOct-Dec2008Nole-WilsonS,TranbyTL,KrizekBA(2005)AINTEGUMENTA-like(AIL)genesareexpressedinyoungtissuesandmayspecifymeristematicordivision-competentstates.
PlantMolBiol57:613–628OhtoM,FischerRL,GoldbergRB,NakamuraK,HaradaJJ(2005)ControlofseedmassbyAPETALA2.
ProcNatlAcadSciUSA102:3123–3128OkamuroJK,CasterB,VillarroelR,VanMontaguM,JofukuKD(1997)TheAP2domainofAPETALA2denesalargenewfamilyofDNAbindingproteinsinArabidopsis.
ProcNatlAcadSciUSA94:7076–7081PassarinhoP,KetelaarT,XingM,vanArkelJ,MaliepaardC,HendriksMW,JoosenR,LammersM,HerdiesL,denBoerB,vanderGeestL,BoutilierK(2008)BABYBOOMtargetgenesprovidediverseentrypointsintocellproliferationandcellgrowthpathways.
PlantMolBiol68:225–237RiechmannJL,MeyerowitzEM(1998)TheAP2/EREBPfamilyofplanttranscriptionfactors.
BiolChem379:633–646RiechmannJL,HeardJ,MartinG,ReuberL,JiangC-Z,KeddieJ,AdamL,PinedaO,RatcliffeOJ,SamahaRR,CreelmanR,PilgrimM,BrounP,ZhangJZ,GhandehariD,ShermanBK,YuG–L(2000)Arabidopsistranscriptionfactors:genome-widecomparativeanalysisamongeukaryotes.
Science290:2105–2110SakumaY,LiuQ,DubouzetJG,AbeH,ShinozakiK,Yamaguchi-ShinozakiK(2002)DNA-bindingspecicityoftheERF/AP2domainofArabidopsisDREBs,transcriptionfactorsinvolvedindehydration-andcold-induciblegeneexpression.
BiochemBiophysResCommun290:998–1009SimmondsDH,DonaldsonPA(2000)Genotypescreeningforproliferativeembryogenesisandbiolistictransformationofshort-seasonsoybeangenotypes.
PlantCellRep19:485–490SrinivasanC,LiuZ,HeidmannI,SupenaENJ,FukuokaH,JoosenR,LambalkJ,AngenentG,ScorzaR,CustersJBM,BoutilierK(2007)HeterologousexpressionoftheBABYBOOMAP2/ERFtranscriptionfactorenhancestheregenerationcapacityoftobacco(NicotianatabacumL).
Planta225:341–351ThompsonJD,HigginsDG,GibsonTJ(1994)CLUSTALW:improvingthesensitivityofprogressivemultiplesequencealignmentthroughsequenceweighting,position-specicgappenaltiesandweightmatrixchoice.
NucleicAcidsRes22:4673–4680TsuwamotoR,YokoiS,TakahataY(2010)ArabidopsisEMBRY-OMAKERencodinganAP2domaintranscriptionfactorplaysakeyroleindevelopmentalchangefromvegetativetoembryonicphase.
PlantMolBiol.
doi:10.
1007/s11103-010-9634-3VerdeilJ-L,AlemannoL,NiemenakN,TranbargerTJ(2007)Pluripotentversustotipotentplantstemcells:dependenceversusautonomyTrendsPlantSci12:1360–1385Wu¨rschumT,Grob-HardtR,LauxT(2006)APETALA2regulatesthestemcellnicheintheArabidopsisshootmeristem.
PlantCell18:295–307ZhangG,ChenM,ChenX,XuZ,GuanS,LiL-C,LiA,GuoJ,MaoL,MaY(2008)Phylogeny,genestructures,andexpressionpatternsoftheERFgenefamilyinsoybean(GlycinemaxL.
).
JExpBot59:4095–4107326PlantMolBiol(2010)74:313–326123
- 2.5bnb998.com相关文档
- diabnb998.com
- 宁波bnb998.com
- 公司bnb998.com
- D76G57Ebnb998.com
- frequencybnb998.com
- N9CObnb998.com
2021年国内/国外便宜VPS主机/云服务器商家推荐整理
2021年各大云服务商竞争尤为激烈,因为云服务商家的竞争我们可以选择更加便宜的VPS或云服务器,这样成本更低,选择空间更大。但是,如果我们是建站用途或者是稳定项目的,不要太过于追求便宜VPS或便宜云服务器,更需要追求稳定和服务。不同的商家有不同的特点,而且任何商家和线路不可能一直稳定,我们需要做的就是定期观察和数据定期备份。下面,请跟云服务器网(yuntue.com)小编来看一下2021年国内/国...

阿里云香港 16核32G 20M 999元/月
阿里云香港配置图提速啦是成立于2012年的十分老牌的一个商家这次给大家评测的是 阿里云香港 16核32G 20M 这款产品,单单说价格上就是十分的离谱原价8631元/月的现价只要 999元 而且还有个8折循环优惠。废话不多说直接进入正题。优惠时间 2021年8月20日-2021年9月20日 优惠码 wn789 8折优惠阿里云香港BGP专线 16核32G 10M带宽 优惠购买 399元购买链接阿里云...
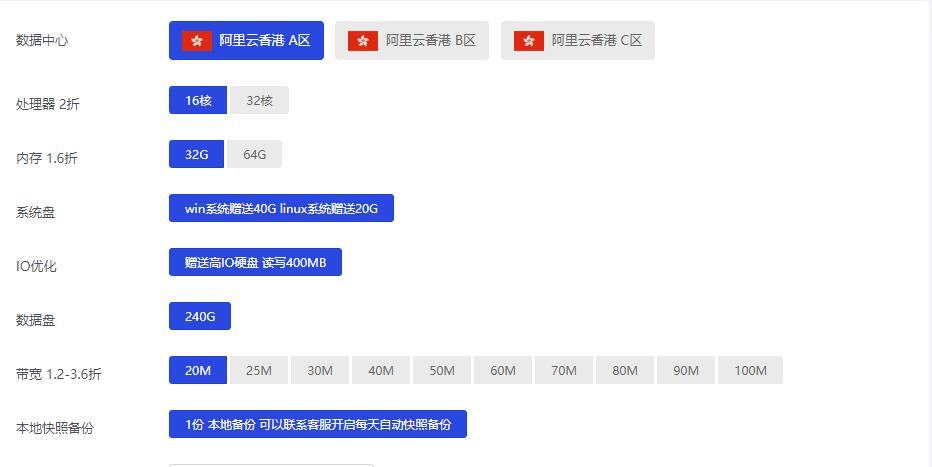
阿里云年中活动最后一周 - ECS共享型N4 2G1M年付59元
以前我们在参与到云服务商促销活动的时候周期基本是一周时间,而如今我们会看到无论是云服务商还是电商活动基本上周期都要有超过一个月,所以我们有一些网友习惯在活动结束之前看看商家是不是有最后的促销活动吸引力的,比如有看到阿里云年中活动最后一周,如果我们有需要云服务器的可以看看。在前面的文章中(阿里云新人福利选择共享性N4云服务器年79.86元且送2月数据库),(LAOZUO.ORG)有提到阿里云今年的云...
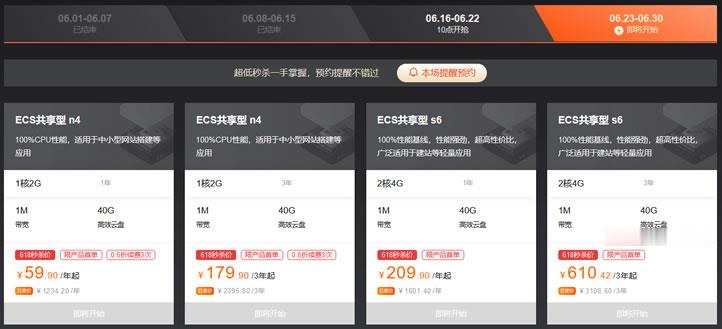
bnb998.com为你推荐
-
2020双十一成绩单2020双十一尾款如何合并付款?嘀动网动网和爱动网各自的优势是什么?百度关键词分析怎样对关键词进行分析和选择www.zhiboba.com上什么网看哪个电视台直播NBAwww.ijinshan.com好电脑要用什么样的软件www.ca800.com西门子plc仿真软件有什么功能www.147.qqq.com谁有147清晰的视频?学习学习45gtv.comLETSCOM是什么牌子?梦遗姐昨晚和姐姐和她朋友一起吃晚饭,我们都喝了酒,我迷糊着回到家的,早上我回想起我好像发生关系射过,会不会是我姐姐,如果是这样我怎么办关键词分析如何进行关键词指数分析