glyk12818.com
k12818.com 时间:2021-04-08 阅读:()
REVIEWARTICLEPhysiology,phylogeny,earlyevolution,andGAPDHWilliamF.
Martin1&RüdigerCerff2Received:7December2016/Accepted:22February2017/Publishedonline:6March2017#TheAuthor(s)2017.
ThisarticleispublishedwithopenaccessatSpringerlink.
comAbstractThechloroplastandcytosolofplantcellsharboranumberofparallelbiochemicalreactionsgermanetotheCalvincycleandglycolysis,respectively.
Thesereactionsarecatalyzedbynuclearencoded,compartment-specificiso-enzymesthatdifferintheirphysiochemicalproperties.
ThechloroplastcytosolisoenzymesofD-glyceraldehyde-3-phos-phatedehydrogenase(GAPDH)harborevidenceofmajoreventsinthehistoryoflife:theoriginofthefirstgenes,thebacterial-archaealsplit,theoriginofeukaryotes,theevolutionofproteincompartmentationduringeukaryoteevolution,theoriginofplastids,andthesecondaryendosymbiosisamongthealgaewithcomplexplastids.
ThereactionmechanismofGAPDHentailsphosphorolysisofathioestertoyieldanenergy-richacylphosphatebond,achemistrythatpointstoprimitivepathwaysofenergyconservationthatexistedevenbeforetheoriginofthefirstfree-livingcells.
Here,werecountthemaininsightsthatchloroplastandcytosolicGAPDHpro-videdintoendosymbiosisandphysiologicalevolution.
KeywordsEndosymbiosis.
Plastids.
Mitochondria.
Cellevolution.
PeterSitteIntroductionPeterSittewasavirtuosointheartofelectronmicroscopy.
Hedevotedhisscientificcareertounderstandingthenatureandevolutionarybasisofcompartmentationineukaryoticcellsandtherolethatendosymbiosisplayedtherein(Sitte2007).
Thanksmainlytoelectronmicroscopicstudiesinthe1960sand1970s,scientistsin2016recognizetwokindsofcells:theprokaryotictypeandtheeukaryotictype.
Themaindifferencethatdistinguishesthetwocelltypesisthenatureofinternalcompartmentationineukaryotes.
Thechromosomesineu-karyoticcellsareseparatedfromthecytoplasmbymembranesurroundingthecellnucleus,whilechromosomesinprokary-otesarefreelydispersedthroughoutinthecytoplasm.
Eukaryotestypicallypossessacomplexendomembranesys-tem,andmitochondria,plant,andalgalcellspossesschloro-plastsinaddition.
Bythemeasureofcompartmentation,themostcomplexcellsinnaturearefoundamongthealgaethatpossessplastidssurroundedbythreeorfourmembranes,plas-tidsthatareremnantsofevolutionarilyreducedeukaryoticcellsresidingwithinthecytosolofanothernucleus-bearingcell(StoebeandMaier2002;Gouldetal.
2008).
Thoughitwasnotalwaysthecase,today,biologistsrecognizethatcom-plexityineukaryoticcellsstemsfromendosymbiosis(Archibald2014).
EndosymbiotictheorytakesrootinMereschkowsky'sclas-sicalessayontheoriginofplastids(Mereschkowsky1905).
Ithasalongandturbulenthistory,asrecentlysummarizedelse-where(Martinetal.
2015).
Theelderofusfirstlearnedaboutendosymbiosisinthe1960sinPeterSitte'scellbiologylec-turesattheUniversityofFreiburg.
Endosymbiotictheory—theprospectthatmitochondriaandchloroplastsdescendedfromfreelivingprokaryotesthatenteredintoasymbioticre-lationshipwiththeirrespectivehostcellearlyineukaryotichistory—wasaveryexciting,almostrevolutionary,prospectThispaperisdedicatedtothememoryofPeterSitte(1929–2015).
HandlingEditor:UliKutschera*WilliamF.
Martinbill@hhu.
deRüdigerCerffcerff-ruediger@t-online.
de1InstituteofMolecularEvolution,UniversityofDüsseldorf,Universittsstr.
1,40225Düsseldorf,Germany2InstituteofGenetics,TechnicalUniversityofBraunschweig,Spielmannstr.
7,38106Braunschweig,GermanyProtoplasma(2017)254:1823–1834DOI10.
1007/s00709-017-1095-yincellevolutionthatopenedupfundamentallynewavenuesofpursuittoinvestigateandunderstandeukaryoticintracellu-larcompartmentation.
Oneaspectinparticularwasimportantforendosymbiotictheory:thecompartmentationofmetabo-lismineukaryotes.
Earlyon,endosymbiotictheoryhaditthatthecoremetabolicfunctionsofmitochondria(respiration)andchloroplasts(photosynthesis)weredirectinheritancesfromthebacterialancestorsoforganelles.
ItwasalsoclearfromelectronmicroscopythatorganellespossessedDNA(KowallikandHaberkorn1971),andthatorganellegenomesweremuchtoosmalltoencodealloftheproteinsthatunder-pinrespirationandphotosynthesis(Herrmannetal.
1975).
Asaconsequence,mostoftheproteinsthatsupportthephysio-logicalfunctionofchloroplastsandmitochondriahadtobeencodedinnuclearchromosomes,whichmeantthattherehadtohavebeensomeformofgenetransfergoingonfromendo-symbiontstothehost,orasWallinputitwithregardtomito-chondria,B.
.
.
bacterialorganismsmaydevelopanabsolutesymbiosiswithahigherorganismandinsomewayoranotherimpressanewcharacteronthefactorsofheredity.
Thesim-plestandmostreadilyconceivablemechanismbywhichthealterationtakesplacewouldbetheadditionofnewgenestothechromosomesfromthebacterialsymbiont.
^(Wallin1925;p.
144).
Chloroplastcytosolisoenzymesprovideduniqueopportu-nitiestotestcrucialpredictionsofendosymbiotictheorywithmolecularevolutionarystudies.
Ifendosymbiotictheorywascorrectaboutchloroplastsarisingfromcyanobacteria,itfollowedthatthenucleargeneforthechloroplastenzymeshouldultimatelystemfromthegenomeofthecyanobacterialancestorofplastids,whilethecytosolicenzymeshouldreflecttheevolutionofthehostcellthatacquiredtheplastid.
Asitappliedtoisoenzymes,thisspecificinferencewascalledtheproductspecificitycorollarytoendosymbiotictheory(Weeden1981).
Chloroplastcytosolisoenzymesforhigherplantglyceraldehyde-3-phosphatedehydrogenases(GAPDH)providedanexcellentsystemtotesttheprospectofgenetransferinendosymbiosis.
GAPDH:fromproteintoDNALikeendosymbiotictheory,GAPDHitselfhasalongandturbulenthistory.
TheenzymaticactivityofGAPDHgoeswayback,bothinplantmetabolismandinenzymologyitself.
Indeed,itwasamongthefirstenzymestobestudied.
EarlyworkonGAPDHfocusedontheenzymefrommuscleandyeast.
Inthe1930s,whenEmbden,Meyerhof,andParnasweremakingheadwayintosynthesisofadenylpyrophosphate(nowcalledATP)fromtheoxidationofglucose,DorothyNeedhamreportedthattheenergyreleasedintheoxidationoftriosephosphateindicatedBTheexistenceinmuscleextractofacoupledmechanismwherebysynthesisofadenylpyrophosphate(fromadenylicacidandfreephosphate)canaccompanyoxido-reduction^(NeedhamandPillai1937,p.
1850).
OttoWarburgreportedthecrystallizationoftheyeastenzyme(WarburgandChristian1939),whichhecalleddasoxydierendeGrungsferment.
BythetimethatCorietal.
(1948)reportedcrystallizationoftheanimalenzyme,itwascalledD-glyceraldehye-3-phosphatedehydrogenase.
Stumpf(1950)reportedGAPDHactivityinplants,notingthattheoxidationoffructose-1,6-bisphosphateinpeaseedlingsoccursbyaseriesofreactionthatBisapparentlysimilartothatinyeastandanimaltissues.
^AGAPDHactivitywasreportedinchlorophyll-containingplanttissuesthatrequiredNADPH(Gibbs1952).
Intothe1970s,itwasnotclearwhetheroneGAPDHenzymeexistedinplantsthatwasmodifiedforNADPH-dependentactivity(Melandrietal.
1970;Cerff1974)orwhetherisoenzymesexistedfortheNADH-andNADPH-dependentactivitiesand,ifthelattercaseweretrue,whethersuchisoenzymeswerespecifictotheGAPDHreactionsofglycolysisinthecytosolandtheCalvincycleofplastids.
Classicalenzymology—sep-aration,purification,andcharacterizationoftheplantenzymes(Cerff1978a,b;Cerff1982a)—demonstratedtheexistenceofcytosol-specific(NADHdependent)andchloroplast-specific(NADPHaccepting)GAPDHisoenzymesinphotosynthetictissuesofhigherplants(Cerff1979;CerffandChambers1979).
Theisoenzymesindeedhadverydistinctevolutionaryhistories(Melandrietal.
1970;Cerff1982a),andtheywereencodedbynucleargenes(Cerff1982b;CerffandKloppstech1982).
Intheearly1980s,therewasalotofexcitementsur-roundingthenewlydiscoveredabilityofmolecularse-quencecomparisonsandphylogenetictrees,notonlytotestendosymbiotictheorybuttoalsoreconstructearlyevolutionmoregenerally.
LynnMargulishadveryeffectivelyrevivedendosymbiotictheory(Sagan1967),butshenevermadethetransitiontotestingitspredictionswithmolecularphyloge-netics.
ItwasMargaretDayhoff(SchwartzandDayhoff1978)whousheredendosymbiotictheoryintotheeraofscientifictestingwithphylogenetictreescreatedfrompro-teinsequences.
Disappointingly,sequencesfortheplantGAPDHisoenzymescouldnotbeobtainedviastandardEdmandegradationproteinsequencingtechnologiesthathadworkedsowellforcytochromecorferredoxins(Dayhoff1965).
ThatmeantthatifonewantedtogettheaminoacidsequencesforplantGAPDHinordertoaddressthedecisiveevolutionaryissuesattheforefrontofthefield,onehadtoutilizethelatesttechnology:DNAsequencing.
Atthattime,theroadtoobtainingproteinsequencesfromDNAsequenceswentthroughcDNAsequencesandanti-bodies.
Methodsforseparatingandpurifyingtheisoen-zymes(Cerff1979)meantthatantiseraagainstthepurifiedproteinswereavailable(CerffandKloppstech1982).
Theavailabilityofantiserapermitteduseofanarchaic,experi-mentallydemanding,butofteneffectivetechniquecalled1824W.
F.
Martin,R.
Cerffhybridreleasetranslation.
Ifallhasgonewell,theresultofthehybridtranslationapproachtocloningdeliversthede-siredcDNAclones,andthechemicalmethodofMaxamandGilbertdeliverstheirsequences(MartinandCerff1986).
TheoriginofplastidsWiththecDNAsandderivedaminoacidsequences,wewereabletoshowthatthenuclearencodedchloroplastenzymewasmoresimilartoitshomologuesfrombacteriathanitwastohomologuesfromeukaryotes,andthatthenuclearencodedcytosolicenzymewasmoresimilartohomologuesfromani-malsandyeastthanitwastohomologuesfromprokaryotes(Fig.
1a).
Thisclearlyboreoutthepredictionsfromendosym-biotictheory,anovelandexcitingfind.
Intheprocessofnotgettingourpaperpublishedintwojournals,however,these-quencesofGAPDHfromEscherichiacolibecamepublished,andreferees,oneintimatelyfamiliarwithE.
coliGAPDH,weresuddenlydemandingthatweexplainwhyE.
coliGAPDHwasmoresimilartoeukaryoticsequencesthanitwastoGAPDHfromThermusaquaticusorBacillusstearothermophilus.
Wereasonedthatthiswasalsoprobablyacaseoflateralgenetransfer(ourworkinghypothesisentailedlateralgenetransferanyway),butnotfromprokaryotestoeukaryotesasinthecaseofthechloroplastenzyme,ratherfromeukaryotestoprokaryotes(MartinandCerff1986).
GAPDHthuspresentsoneoftheearliestreportsinthelitera-tureforsequence-basedinferenceoflateralgenetransfer,aconceptthatadecadelaterbecamequitepopular(toopopularinsomecirclesperhaps),andsequencesfromotherplantsourcesboreoutthepredictionsofendosymbiotictheorywithrespecttotheplantenzyme(Brinkmannetal.
1987).
ChloroplastGAPDHuncoveredadditionalsurprises.
TheNADPH-utilizingplastidenzymefromhigherplantswasknowntoexistintwoforms,anA4homotetramerandanA2B2heterodimer(CerffandChambers1979).
TheAandBsubunitswereshowntobetheresultofanucleargeneduplicationthattookplaceearlyintheevolutionofthegreenplantlineage,withtheBsubunithavingacquiredashortC-terminalextensionwithconservedcysteineresidues(Brinkmannetal.
1989).
TheC-terminalextensionofGapBwasacquiredatthebeginningoflandplantevolutionfromthenuclearencodedsmallredoxactiveproteinCP12,whichwasshowntointeractwiththeA2B2andA4formsofchlo-roplastGAPDHinadditiontophophoribulokinaseintheabsenceofNADP(H)(Wedeletal.
1997,WedelandSoll1998,Petersenetal.
2006a).
ThisinteractionblockedCO2fixationactivityinthedarkandpreventedfutilecyclingbetweenglycolysisandtheCalvincycle.
ItalsoexplainedwhythechloroplastenzymeaggregatedinthepresenceofNAD(H),whichwasthekeytoefficientseparationandpu-rificationoftheisoenzymes(Cerff1982a).
TheoriginofthefirstgenesIntheearly1980s,beforetheconceptofanRNAworld(Gilbert1986)hadbeenborn,peoplewerestillvigorouslydebatingtheissueofwhatcamefirst,protein,orDNA.
Oneofthebigpuzzleswashowthefirstlongopenreadingframesingenescametobe,andhowenzymesizedproteinsaroseintheabsenceofaccuratetemplatereplication.
Figuringpromi-nentlyinthatdebatewasWalterGilbert'sexontheoryofgenes(Gilbert1987),accordingtowhichintronswererelictsfromtheprimordialassemblyofgenesatlife'soriginandthatintronpositionsinmoderngenescorrespondedtotheboundariesbetweenstructuralmodulesofproteinfunctioncalleddo-mains.
Modules,beingshorterandeasiertoevolve,couldrecombineviaBexonshuffling^andexonscouldperhapsun-dergoalternativesplicing(Gilbert1978),therebypromotingancientenzymediversity.
AncientenzymessuchasGAPDHClostridiumpasteurianumaThermusaquaticusBacillusstearothermophilusAnabaenavariabilismaizepeatobaccomaizepeatobaccoGapAGapBChloroplastCyanobacteriumbFig.
1PhylogenetictreesofGAPDHaminoacidsequences.
aThetreewasconstructedbythemethodofFitchandMargoliash(1967)withtwointeractiveroundsoftopologicaladjustmenttoreducethesumofsquareddifferencesbetween(i)pairwisedistancescalculatedbetweenaminoacidssequencesusingtheminimummutationdistanceofDayhoff(1965)and(ii)pairwisedistancesmeasuredintheconstructedphyloge-netictree(Martin1985).
Senf,mustard;Hefe,yeast;Huhn,chicken;Mensch,human;Schwein,pig;Hummer,lobster;B.
,Bacillus,T.
,Thermus.
Numbersatbranchesaredistances.
bThenuclearencodedgenesfortheAandBsubunitsofhigherplantchloroplastGAPDH,anA2B2tetramer,branchwiththeCalvincyclehomologuefromcyanobacteria.
RedrawnfromMartinetal.
(1993)Physiology,phylogeny,earlyevolution,andGAPDH1825wereclearlywellsuitedtotestthoseideas,andindeed,wefoundintronpositionsthatwerepresentbetweenthesamenucleotidesinthesamehomologouscodoninthenucleargeneforchloroplastGAPDHandinanimalGAPDH(Quigleyetal.
1988).
ThecloserwelookedintoGAPDHgenes,themoreevidencewefoundforidenticalintronpositionsinancientlydivergedgenes(Liaudetal.
1990;Kersanachetal.
1994;Cerffetal.
1994).
Duringthoseinvestigations,however,ourviewsconcerningtheageofeukaryoticGAPDHgeneshadtoberevised,andlikesomanyotherthingsinevolution,cyanobacteriawereresponsibleforthechange.
EndosymbiosisandtheunexpectedoriginsofeukaryoticgenesTheendosymbiosisstoryfortheoriginofchloroplastGAPDHwasconceptuallysatisfying,butanimportantpiecewasmiss-ing.
Thepredictionfromendosymbiotictheorywas,namelythattheplastidenzymeshouldbemoresimilarnotjusttoprokaryotichomologuesingeneral,buttocyanobacterialho-mologuesinparticular.
Therefore,weembarkedtoobtainthesequenceofGAPDHfromcyanobacteria,whichwedid,find-ingmorethanweexpected.
Wefoundthatthecyanobacteriumthatwehadinvestigated,Anabaena,indeedhadaCalvincyclehomologueofGAPDHthat,inphylogenetictrees,branchedveryspecificallywiththenuclearencodedchloroplastGAPDHenzymesofhigherplants(Fig.
1b),aresoundingconfirmationofthepredictionfromendosymbiotictheory(Martinetal.
1993).
However,thatwasnotallthatAnabaenahadinstore,italsoharboredtwootherGAPDHgenes,oneofwhichsheddramaticlightonthepeskyE.
coliGAPDHthatwasannoyinglysimilartoeukaryoticGAPDH.
AnabaenahadacopyofthesameE.
coliGAPDHgenethat,upuntilthen,wehadthoughttobeaeukaryote-to-prokaryotetransfer.
Wehadbecomeprettygoodatinterpretingtrees,butthisobservationwasapuzzle.
Inordertomakesenseofit,wehadtopaycarefulattention(i)towhatReinhardHenselandhisteamhadbeenfindingforarchaealGAPDH(FabryandHensel1988;Zwickletal.
1990)and(ii)towhateveryonewassayingatthetimewithregardtoeukaryoteorigins.
Intheearly1990s,WoeseandKandler'srootedthreedomaintreewasasenvogueasitgetsitbiology(Woeseetal.
1990).
However,itwasatreeofribosomes.
Theassumptionbackthen,beforewehadgenomes,wasthattherRNAtreewasspeakingforthegenomeasawhole,andthatultimately,whenwehadallthedata,allgeneswouldtendtopaintroughlythesamepictureastherRNAtree,witharchaeaasthesisterstotheeukaryotesandwithmitochondrion-lackingeukaryotesbranchingearlyontheeukaryoticbranch,meaningthatmito-chondriacamelateineukaryoteevolutionandthateukaryotesare,forallpracticalpurposestobeseenasgrown-uparchaea.
Accordingly,eukaryoteGAPDHgenesshouldderivefromarchaealGAPDHgenes.
Thatwasnot,however,whatReinhardHenselwasfinding(FabryandHensel1988;Zwickletal.
1990).
TheywerefindingthatarchaealGAPDHwas,forallpracticalpurposes,unrelatedtoeukaryoteGAPDH,butitwasalsojustasunrelatedtobacterialGAPDH.
ThatindicatedasplitofarchaealGAPDHfromitshomo-loguesintheothertwodomainsattheearlieststagesofproteinevolution,whichinitselfwasnothugelysurprising.
However,fromourstandpoint,tryingtomakesenseofdatathatwasfundamentallydifferentfromwhateveryoneelsewasfinding(Fig.
2),therewasasubstantialsurprise.
Ifwethoughtthingsthroughinfull(whichissomethingwespentalotoftimedoinginthosedays),italsoindicatedthatalloftheGAPDHgenesthatwehadseenineukaryotesupuntilthenwere—justlikechloroplastGAPDH—acquiredfrombacteria.
Eukaryoticgenomeswerenotgenerallyarchaeal,theywerechimeric.
Becauseanimalsandfungihadneverpossessedplastids(sosaystheguidinglogicofendosymbiotictheory),thisledustopositthattheeukaryoticgeneforglycolyticGAPDHisalsoanacquisitionviaendosymbiosisandultimatelystemsfrommito-chondrion.
CytosolicGAPDH,acoreenzymeofcarbonandenergymetabolismandthekeyenergy-conservingstepofgly-colysis,reflectedtheevolutionofthehostthatacquiredtheplastid,butnottheevolutionofthehostthatacquiredthemi-tochondrion.
Endosymbiosishadmoretodowithcompartmen-talizedenergymetabolismineukaryotesthananyonehadsuspected.
Weconcludedbysaying:BTheevidenceforeubac-terialoriginofeukaryoticGAPC,anessentialglycolyticen-zyme,suggeststhateukaryoticgenomesaremorehighlychi-mericthanpreviouslyassumed.
Whereasmostorganellarpro-teinsarecurrentlyencodedinthenucleus,endosymbiontsmayhavedonatedmanygenestothenucleuswithoutorganellarreimportoftheprotein,therebyenrichingthegeneticandmet-abolicpotentialofthehost^(Martinetal.
1993;p.
8695).
Webegantonoticethatendosymbioticgenetransferandendosym-bioticgenereplacementwereprobablymorewidespreadthananyonehadsuspected;itwasanexcitingtime.
Notuntilmanyyearslater,withtheavailabilityofcompletegenomes,didpeopleseethatthevastmajorityofeukaryoticgenesstemfrombacteria,notfromarchaea(Esseretal.
2004;Pisanietal.
2007;CottonandMcInerney2010;Kuetal.
2015).
Today,weknowthateukaryotesarenotjustgrownuparchaea,theyaretheproductofendosymbiosis(Zimorskietal.
2014;Martinetal.
2015),butforsomereason,itisstillverypopulartosaythateukaryotesarosefromarchaeaortowriteabouttheBarchaealoriginofeukaryotes^(Williamsetal.
2013;Koonin2015),eventhoughthestatementis,ifwelookatthedataopenly,flatlywrongbecauseeukaryotespossessthreetimesmorebacterial-derivedgenesthattheypossessarchaealderivedgenes(Esseretal.
2004;Pisanietal.
2007;CottonandMcInerney2010;Kuetal.
2015).
Whatthedatasay(Zaremba-Niedzwiedzkaetal.
2017)isthatthehostfortheoriginofmitochondriahasanarchaealorigin(McInerney1826W.
F.
Martin,R.
Cerffetal.
2014).
AdemocraticgenomewithonegeneonevotewouldelecttheBbacterialoriginofeukaryotes.
^GAPDHwasjustthetipoftheiceberg.
TheevolutionofproteincompartmentationduringeukaryoteevolutionThefindingthatendosymbiontsmayhavedonatedmanygenestothenucleuswithoutorganellarreimportofthepro-tein(Martinetal.
1993)sharpenedourawarenesstoanissuethateventuallybecamequitepressing.
We(andeveryoneelse)hadbeenassumingthattheproductsofendosymbiont-derivedgeneswerespecifictotheorganellefromwhichthegenewasacquired;afterall,thiswaswhattheproductspecificitycorollarytoendosymbiotictheory,whichwehadbeentestingforadecadesaid.
CytosolicGAPDHwasanexception,andaplastid-targetedversionofcytosolicGAPDHwasalsoanexception(Meyer-Gauenetal.
1994;Petersenetal.
2003).
Whenwelookedaroundmoregenerally,almostalloftheenzymesofcentralcarbo-hydratemetabolisminplantswereanexception,anditwasquicklybecomingapparentthateukaryoteshadabacterialglycolyticpathway(Martin1996),whichwasalsotrueofeukaryotessuchasGiardiaorEntamoebathatlackedtypi-calmitochondria(Henzeetal.
1995),asshowninFig.
3.
Allthatprovidedcausetothinkabouttheprocessofendo-symbioticgenetransferasaprocessinrealtime.
Whenthebacterialancestorsofchloroplastsandmitochondriabecameendosymbionts,theydidnotbringalongtheirownpre-existingproteinimportmachinery.
Rather,boththeTiMandToMproteinimportmachineryofmitochondriaandtheTiCandToCproteinimportmachineryofplastidsareeukaryoticinventions,albeitconsistingofsomeprokaryotic-derivedba-siccomponents(SollandSchleiff2004),arosewithintheeukaryoticlineage,eachfollowingtheoriginoftherespectiveorganelle.
Theconsequenceofthatinsightisthatattheonsetofmitochondrialendosymbiosis,whenthefirstgeneswerebeingtransferredtothechromosomesofthehost,therewasnoplaceforthetranslatedproductsofendosymbiont-derivedgenestogoexcepttothecytosol(ortothehost'splasmamembrane).
Thatinturnmeansthatitshouldnotbeallthatsurprisingtoseeapredominanceofbacterialgenesineukary-otes.
Wewereassuming20yearsagothatwhichtoday'sdatasay(Zaremba-Niedzwiedzkaetal.
2017):Thehostthatac-quiredthemitochondrionwasanarchaeon,aprokaryote.
Genetransfersduringendosymbiosistransformedthearchaealhostfromwithinbecauseifthefirsttransferdidnotwork(thatis,ifitjustgaverisetoapseudogene),thenmaybethenexttransferwouldbesuccessful,ormaybethenextafterthat.
Attheoriginofeukaryotes,themitochondrialendosymbi-ont'sgenomewasacontinuousandpersistentsourceofnewgenesforthehost.
Attheonsetofthatsymbiosis,however,theproductsofthefirsttransferredgenesbecametargetedtothehost'scytosol.
OnlyafterthesymbioticconsortiumhadevolvedtheTiMandToMcomplexesinthetwomitochondri-almembranescouldtheproductsoftransferredgenesbereimported.
TheevolutionoftheTiMandToMcomplexesfosteredtheevolutionofN-terminaltransitpeptidesthatwoulddirectgeneproductsfaithfullytomitochondria.
GAPDHshedlightonthatprocess,too,andintronswereonceagaininvolved.
WalterGilbert'steamfoundacaseinwhichtheNAD-bindingdomainofcytosolicGAPDHhadbeenappendedtotheN-terminusofplantcytochromecviaexonshufflingtogenerateafunctionaltransitpeptide(Longetal.
1996).
Thus,whilechloroplastGAPDHpossessedtransitpep-tides,cytosolicGAPDHinplantsprovidedgeneticstartingmaterialforthegenerationoftransitpeptides,yetnotdenovo,ratherbyexonshuffling.
Thatwasallextremelyhelpfulforunderstandingthesitua-tion,butsomeoldimportantquestionsremainedandnewonesarose.
Amongtheoldimportantquestionswastheissueofwhy,withallthattransfergoingon,anygenesremainedinorganellesatall,whichJohnAllen'sCORRhypothesisexplainsbetterthananyothercompetingtheory,ithastodowiththeneedtomaintainredoxbalanceinbioenergeticmembranes,whichrequiresthatcomponentsoftheelectrontransportchainbeencodedintheorganelle(Allen2015).
Amongthenewquestionstounfoldfromthoseinsightswastheissueofwhyeukaryotes,whicheveryonethoughtdescendfromarchaea(sayribosomes),shouldhaveabacterialglycolyticpathwayinthefirstplaceinsteadoftheiroriginalarchaealglycolyticpathway.
Keepingthebiggerpictureinfocus—andremainingcriticalofthetheoreticalfoundationsuponwhichthewholeendosymbiosisendeavorwasresting—perhapstheanswerwasthatsomethingelseevenmorefunda-mentalwaswrongwithendosymbiotictheory.
WhatwaswrongAslongasglycolysishadbeenknownandforaslongEFGaEFTuVATPase16SrRNARNApolymeraseGAPDHLateraleubacterialtransferandsubstitutionofnucleargapCbEubacteriaEukaryotesArchaebacteriaEubacteriaEukaryotesArchaebacteriaEubacteriaEukaryotesArchaebacteriaFig.
2Domainrelationships.
aSchematicrepresentationofdomainrelationshipsforseveralmarkersusedatthetimethatshowedasistergrouprelationshipbetweeneukaryotesandarchaeacontrastedtothedomainrelationshipsdepictedbysequencesforglycolyticGAPDHsequences.
bInterpretationofthedifferencebetweendomainrelationshipsdepictedbythedatasets.
RedrawnafterMartinetal.
(1993)Physiology,phylogeny,earlyevolution,andGAPDH1827asanyoneeverthoughtthatmitochondriawereendosymbi-onts,everyonehadalwaysassumedthatthehostfortheoriginofthemitochondrionwasaheterotroph.
WhatifthehostwasanautotrophAnautotrophwouldactuallyrequireanewgly-colyticpathwayinordertosupplyitsmitochondriawithcar-bonsubstratesbecauseinautotrophs,carbonfluxisspecial-izedandoptimizedtorunfrompyruvatetoglucose.
Inordertofeedamitochondrioninthecytosolofanautotroph,atypicalbacterialEmbden-Meyerhofcataboliccarbonfluxwasneededasitoccursinalleukaryotes.
Stumpf's(1950)inferenceaboutglycolysisinplantsbeingBsimilartothatinyeastandanimaltissues^appliedtoalleukaryotesingeneral.
Glycolysiswaspresentintheeukaryoticcommonancestor,butwasitpresentinthehostAtthesametime,thereweretheanaerobicformsofmitochondriaoutthereforwhichendosymbiotictheoryalsodidnotaccount,hydrogenosomes(LindmarkandMüller1973;Müller1988).
Ifthehostwasanautotrophicarchaeon,thenitwouldbedependentuponH2,likemostarchaealautotrophsare(Fuchs2011).
IfitweredependentuponH2producedbythecommonancestorofmitochondriaandhydrogenosomes,thatwouldprovideaselectivepressuretoassociatetheendosymbionttoitshost(MartinandMüller1998).
Finally,wehadaversionofendosymbiotictheorywherethehostactuallyneededitsen-dosymbiont,butaverydifferentversionthanhadeverbeenoutthere,oneinwhichbygenetransfer,thesymbiont'sbac-terialglycolyticpathwayendedupinthecytosol,feedingthemitochondrion,andconvertingtheautotrophichostcellcom-partmentintoaheterotrophiccellcompartment(MartinandMüller1998).
Notonlywouldabacterialglycolyticpathwayineukaryotesmakesense,butsowouldthewidespreaddis-tributionofanaerobicpathwaysinmitochondria(Mülleretal.
2012).
Intermsofphysiology,andintermsofvisiblesyntrophicmicrobialinteractions(FenchelandFinlay1995),anautotrophichostfortheoriginofmitochondriaworksmuchbetterthanthephagocytosinghostthateveryonehadalwaysassumed.
Whilemicrobiologiststendedtoliketheidea(Madiganetal.
2012),inevolutionarycircles,thepriceforaphysiolog-icallybasedunderstandingoftheprokaryote-to-eukaryotetransitionwassteep.
Itmeantthat(i)allofMargulis'sversionsofendosymbiotictheory(Margulisetal.
2006)werewrong,thatdeDuve'sinterpretationsofhydrogenosomeswerewrong(deDuve2007),thatWoese'sthreedomaintree(Woeseetal.
1990)waswrong,andthatDoolittle's(1998)viewofunre-strictedandcontinuousinterdomainlateralgenetransferwasalsowrong.
Thatwasalotofauthoritybeingwrongallatonce.
However,theydidnotagreeonanythingamongthem-selveseither,andourintentwastogetthescienceright,nottoproveparticularscientistsright.
Lookingatthematteropenly,everyonewastryingtoaccountfortheirownfavoriteexplananda(flagella,peroxisomes,onebranchinonetree,andmanybranchesinmanytrees).
Weweretryingtoaccountforphysiology,thewaythatcellsstayaliveintheirenviron-ment,onthesimplebutrobustpremisethatchloroplastsandmitochondriawereoncefree-livingbacteria.
Theobjectofourundertakingwastoobtainsomeinsightsintocellevolutionfromthestartingpointofanimportantenzymecentraltocar-bonandenergymetabolism.
Naturedoesnotdivulgesecretsaboutearlyevolutionlightly,regardlessofwhetheronesetstheleveronphysiology,orelsewhere.
SecondaryendosymbiosisamongthealgaewithcomplexplastidsWelearnedfromGAPDHinteraliathatwhenitcomestoendosymbioticgenetransfer,thereisnohomingdevicethatdirectstheproductoftransferredgenesbacktotheorganellefromwhichthegenestems,ratheritisuptonaturalvariationandnaturalselectiontodeterminewherethegeneproductwillendup.
Therewasonceadebateaboutproteintargetinginiti-atedbydeDuve,whoarguedthatperoxisomeswereonceendosymbiontslikemitochondria,becausehowelse(sohisargument)couldonegetanentirepathwayintoanewcom-partmentFollowingdeDuve'sreasoningforthesymbioticoriginofperoxisomes(deDuve1969),MichelsandOpperdoes(1991)explainedwhyspecifictargetingofanen-tirepathwaytoaneworganelledoesindeedseemveryunlike-lyatfirstsight,itisbecauseofthis:Geneduplicationandmutationcanreadilyleadtothesignalneededtodirectoneenzymetoanewcompartment,butbyitself,thenewenzymeisuseless,forlackofsubstrate,forlackofdownstreamprod-uctconversion,orboth;hence,thenewtargetingvariantwillbeaburdenforthecellandwillbelost.
However,ifweconsiderthepossibilitythatproteintargetingwithineukaryot-iccellsmightnotalwaysbe100%specific,withminorZeaChondrusGallusDrosophilaEntamoebaGiardiaChlamydomonasSaccharomycesChlorophytaRhodophytaMetazoaMetazoaRhizopoda(nomt)Metamonada(nomt)ChlorophytaAscomycotaGapCGapCGapCGapCGapCGapCGapCGapCFig.
3LineagerelationshipsforGAPDHfromeukaryoteslackingtypicalmitochondria(abbreviatedasBnomt^).
RedrawnafterHenzeetal.
(1995)1828W.
F.
Martin,R.
Cerffamountsofmanyproteinsconstantlybeingimportedintothewrongorganelle,theneventheretargetingofentirepathwaysduringeukaryoteevolutiondoesnotseemsodifficult(Martin2010)becauseaverysmallamountofenzymecangoalongwayintermsofactivity,andaverysmallamountofanentirepathwayprovidesaunitoffunctionuponwhichselectioncanact.
Nowhereinnatureistheissueofcellcompartmentationmorecomplexthaninalgae.
TheconceptofsecondaryendosymbiosisgoesbacktoRobertE.
Lee(1977)andtoSarahGibbs(1978).
Itpositsthat—touseSitte'sterm—thecomplexplastidsofalgaede-scendfromtheengulfmentofaeukaryoticalgabyaeukary-otichost;suchplastidsaresurroundedbythreeorfourmem-branes.
GeneslikethatforchloroplastGAPDHthatstemfromthecyanobacterialancestorofplastidsshouldalsoreflectplas-tidphylogeny,buthavingbeentransferredtwiceduringevo-lution:onceduringprimaryendosymbiosisfromthecyanobacterialancestorofplastidstothenucleusofthehostandasecondtimefromthenucleusofthealgalendosymbionttothenucleusofitshost.
GAPDHgenesfromredalgae(Liaudetal.
1994)wereextremelyinsightfulinthisrespectastheytracedtheredalgalancestryofplastidsacrossthebreadthofthephotosyntheticeukaryoteshavingredcomplexplastids(Liaudetal.
1997;Liaudetal.
2000).
GAPDHgeneshelpeddiscriminatebetweencompetinghypothesisforthehighlyde-batednumberofsecondarysymbiosesinvolvingredalgalendosymbiontsthatoccurredduringeukaryoticevolution(Petersenetal.
2006b),andtheyhelpedtoclarifytheevolu-tionaryoriginoflandplantsthemselves(Petersenetal.
2006a).
RoundingoutthefamilyofplantGAPDH,therewasalsoanon-phosphorylatingformoftheenzyme,calledGAPN,thatturnedouttobeamemberofthealdehydedehy-drogenasefamily,whichdidhavehomologuesamongar-chaea,providingadditionalinsightsintotheearlydiversifica-tionofgenesinthelastuniversalcommonancestorofalllife(Habenichtetal.
1997).
ThereasonforbringingGAPDHoutofthecoldroomandintotherealmofmolecularbiologyinthefirstplacewastodiscriminatebetweencompetinghypotheses.
Amongtheal-gaewithcomplexplastids,thenumbersofpossibleendosym-bioticeventsandgenetransfersthatonecaninferfromGAPDHgenephylogeniescanbecomebewildering.
Intryingtoreconciletheobservedphyleticpatternswiththebiologyoftheorganisms—theexplanandumoftheintellectualexer-cise—itiseasytolosesightoftheforest(thebiology)forthegenetrees.
Genetreesarehelpfulwhenitcomestohy-pothesistesting,andtheyneitherhavethesameweight,asobservationsinnature,noraretheyeveninthesamecategory.
ThealgaepossessingGAPDHgenesandtheGAPDHgenesthemselvesarerealinthesensethattheyexistinnature.
Bycontrast,branchesingenetreesarenotreal.
Branchesintreescomefromcomputerprograms(orinthecaseofFig.
1,fromapencil,pen,sliderule,andLetraset).
Inordertobringthebranchingpatternsingenetreesintologicalagreementwithwhatwethinktruespeciesrelationshipsshouldbe,oneonlyneedstoaddfouringredients:phylogeneticerror,genelosses,endosymbioses,and(sinceour1986paperanyway)lateralgenetransfer.
Intheabsenceofaprioriknowledgeaboutthesubjectofinquiry,forwhosestudythetreewasconstructed,thereisnosimplewaytodecidehowmuchofwhichingredienttoadd.
Thatcallsforasoundphilosophicalfoundationforthescien-tificenquiryathand,somethinginwhichtheelderofusamplyreceivedformaltrainingduringadecadeofresearchinFreiburg.
ThephilosophyofscienceasitappliestopracticalempiricalinquiryissomethingaboutwhichweoftendiscussedinHannoverandBraunschweig,especiallywhenitwastimetothinkthingsthroughinfull,inthehopeofextractingbelastbare[Ger:sturdyinthesenseofbeingabletosupportsomething]insightsintoearlyevolutionfrompro-teinsequencedata.
Withoutgoingintojustification,detailordigression,twosetsofguidelinesalwaysseemedhelpful.
OneistheprincipleofOccam'srazorassplendidlydefinedinWebster's9thCollegiateDictionary:Bascientificandphil-osophicrulethatentitiesshouldnotbemultipliedunnecessar-ilywhichisinterpretedasrequiringthatthesimplestofcom-petingtheoriesbepreferredtothemorecomplexorthatex-planationsofunknownphenomenabesoughtfirstintermsofknownquantities.
^Thelast12wordsarethemostimportant.
TheysetOccam'srazormilesapartfrommereparsimony,andinpracticalterms,theytranslatetoausefuldoctrineforinfer-ence:Donotmakeupanythingunlessitisabsolutelyneces-saryandworkwithobservationsinnature.
AnothersetofreasoningguidelinescanbefoundinalittlenotedpaperbyLeeaboutthenumberandnatureofplastidsymbioses:BAnyevolutionaryschemeshouldadheretothefollowingthreeprinciples.
(1)Amonophyleticoriginofanyorganism,chemicalcompoundorcytoplasmicstructurehasthegreateststatisticalprobabilityofbeingcorrect.
(2)Thelossofanon-essentialstructurecanrequirejustthemutationofasinglegenebuttheacquisitionofastructuregenerallyrequiresmanymutationsandaconsiderableamountoftime.
(3)Mostorganismsinevolutionarysequenceswouldhavebeenlost,yetinpostulatingphylogeneticeventstheplausibilityofthetheorycanbeenhancedbytheexistenceoforganismssimilartothoseintheproposedscheme.
^(Lee1972,p.
44).
Manyoftoday'spapersaboutlateralgenetransferamongeukaryotescouldbenefitfromLee'srules,no.
2inparticular.
Thioesters,energy,andchemicalrelictsfromlife'soriginGAPDHcatalyzesawell-studiedchemicalreaction;themechanismofwhichisofinterest(SegalandBoyer1953,Bieseckeretal.
1977).
Intheoxidativedirection,thealdehydecarboninD-glyceraldehyde-3-phosphateisattackedbythethiolofCys149(BacillusstearothermophilusresiduePhysiology,phylogeny,earlyevolution,andGAPDH1829numbering)formingacovalentlyenzyme-boundhemithioacetal.
HydrideremovalbyNAD+oxidizesthesub-strate'sthiol-bondedcarbonatomtogenerateathioester,whichharborsanenergy-richbond(Lipmann1941)withafreeenergyofhydrolysisof32kJmol1(BuckelandEggerer1965).
Thesubstrateishowevernotremovedfromtheenzymebyhydrolysis,whichwouldreleasetheenergyasheat,ratherphosphorolysisreleases1,3-bisphospho-D-glycerate.
Theacylphosphatebondof1,3-bisphospho-D-glycerateconservestheenergy;ithasafreeenergyofhydro-lysisontheorderof52kJmol1(Meyerhof1951,Thaueretal.
1977)sufficienttoreadilyphosphorylateADPtoATP(freeenergyofhydrolysis31kJ·mol1,Thaueretal.
1977)viasubstratelevelphosphorylation(Fig.
4a).
Preciselythissequenceofreactionsfromthioestertoacylphosphate(amixedanhydride)totheβ-γphosphoanhydridebondinATPhaslongbeenthoughttobearelicfromtheverybeginningofmetabolism.
DeDuve(1991)suggestedthatsuchathioester–acylphosphate–ATPconversion,asitoccursinglycolysis,mightrepresenttheprimordialpathwayofenergyconservation,buthewaslookingattheproblemofearlyevo-lutionfromthestandpointofsugaroxidations.
Theproblemwithhisproposalisnotthebasicchemistry.
Rather,theprob-lemisthesurroundingenvironmentsupportingsuchareactionsequence.
Likegenerationsofscientistsbeforehim(Haldane1929;Wald1964),deDuvewasassumingthatlifestartedoffwithfermentations.
Thatseemslikeaharmlessassumption,butifwethinkitthroughinfullonthebasisofwhatweknowtodayaboutmicrobialphysiology,itturnsouttobealtogetheruntenable(Schnheitetal.
2016).
Forprimordialenergycon-servationintheoxidativedirectionoftheGAPDHreaction,themainproblemsaretwofold.
First,ontheearlyEarth,thereisnoevidencetosuggesttheexistenceofanyspontaneouslysynthesizedreservoirofanyparticularsugarisomer,noristherereasontoassumethatsuchamountainofsugarevenexisted.
ThisisthesameproblemencounteredwhentryingtoharnessenergyinsomemannerfromorganicsdeliveredtoEarthfromspace:Theorganiccompoundsincarbonaceouschondritesarenotonlyextremelyheterogeneousinstructure(Sephton2002)sothatnospecificisomercouldbecomethefocusofbiologicalenergyconservation,buttheyarealsounfermentablesubstrates(Schnheitetal.
2016),meaningthattheonlywaytoharnessenergyfromthemisthroughoxida-tion,whichrequiresanoxidant,anelectronacceptorlikeO2orFe3+orsimilar.
Theneedforanoxidantbringsustothesecondproblem.
Theinvocationofoxidants,suchasFe3+astheoriginalelec-tronacceptorsfortheoxidativesynthesisofthioestersviaareactionsimilartothatcatalyzedbyGAPDH(deDuve1991),hasanoftenoverlookednegativeconsequenceofgeneralsig-nificance.
InthepresenceofstrongoxidantslikeFe3+,carbonequilibriumliesonthesideofCO2ratherthanonthesideofreducedcarboncompounds,thebuildingblocksoflife(Sousaetal.
2013,AmendandMcCollom2009).
Fe3+isastrongoxidant.
Themidpointpotential,E0',oftheFe3+/Fe2+coupleis+772mV,thatoftheO2/H2Ocoupleis+818mV(Thaueretal.
1977).
Attheonsetofbiochemicalenergyharnessing(orattheoriginoflife,asoneprefers),thefirstandforemosthurdleisthegenerationandaccumulationofreducedorganiccompoundscomprisingthesubstanceofcells,nottheircon-versiontoCO2.
Neitherdisproportionations(fermentations)noroxidationofreducedorganiccompoundsofferatenablepathtogetstartedatbiochemicalorigins.
Whennavigatingthewatersofearlyevolution,physiologyisagoodcompass.
Seenasachemicalprocess,theenergymetaboliccomponentofphysiologyentailsfocusingtheflowofa(modestly)specificspectrumofsubstratesthoughincreas-inglynarrowchannelstowardsveryspecificexergonicreac-tionswhereenergyisconserved.
OntheearlyEarth,themainformofcarbonwasCO2(Sleepetal.
2011).
WecansaythatbecausetheearlyEarthwentthroughaphasewheretheplanetwasmoltenrock,whichishotterthan1000°C.
CarbonincontactwithaglobaloceanofmagmaexistsasCO2,notasglucoseoranythingsimilar.
WhentheEarthcooled,CO2remainedasthemaincarbonspecies.
Thatisgoodinaphys-iologicalsensebecauseitprovidesaveryspecificsubstrateforcorephysiologicalreactions.
Thesurprisingaspectformanyobserversisthatunderstrictlyanaerobicconditions,whichexistedontheearlyEarth,biologicalsystemscanactuallyharnessenergyfromCO2,withoutlight,whenusingH2astheelectronsource:Acetogens(SchuchmannandMüller2014)and,undersomeconditions,methanogenssuchasMethanosarcinaacetovoransgrowingonCO(RotherandMetcalf2004)cou-pletheexergonicreductionofCO2withH2tothesynthesisofATP.
ThatexergonicprocessofATPsynthesisentailsthesamesequenceofsubstratelevelphosphorylationinvolvingthioester–acylphosphate–ATPconversionasdeDuvehadinmind.
However,neitherthesubstratelevelphosphorylationsthatoccursinacetogensandmethanogensnortheenzymesinvolvedhaveanythingtodowithglycolysis(Fig.
4b).
EnergyisreleasedinthereductivedirectionbecauseinthereactionofH2withCO2,thechemicalequilibriumliesonthesideofreducedcarboncompounds(ShockandBoyd2015).
TheH2requiredtodrivesuchreactionsinthedirectionofCO2reductionwaspresentontheearlyEarth,inabun-dance,suppliedthroughageochemicalprocesscalledserpentinization,whichhasbeengeneratingH2inhydrother-malsystemsinmillimolesperliteramountssincetherewaswateronEarth(Sleepetal.
2004),andstilldoessotoday(McCollomandSeewald2013).
Acylphosphatesmighthaveevenbeentheancestralenergycurrencyoflife,moreancientthanATP(Martin2012).
Indeed,theirsynthesisfrommethylgroupsandCOinanaerobicautotrophsentailsreactionscata-lyzedbytransitionmetalsonly(Sousaetal.
2013),pointingtoaveryancientrolefortransitionmetalsinbiochemical1830W.
F.
Martin,R.
Cerffevolution,somethingthatbiologistshavealwayssuspected(EckandDayhoff1966).
Insupportofthatview,reconstruc-tionsofearlyphysiologicalevolutionfrominvestigationofancientgenes(Weissetal.
2016)providegenome-basedevi-dencethatthecommonancestorofalllifeconservedenergyfromthetransitionmetal-dependentreductionofCO2withelectronsfromH2.
Thus,fromtheperspectiveofthermodynamics,naturalH2-producinggeochemicalprocessescouldhaveenablereactionsinvolvingthesynthesisofsmallmolecularweightcarboncompoundsfromCO2tobecoupledtotheexergonicsynthesisofhigh-energyphosphatebonds.
Areactionsequenceofthattypecouldhaveresidedattheoriginofphysiologicalenergyconservation.
Ifso,thioesterswouldhaveprecededacylphos-phatesinevolutionandacylphosphateswouldhaveprecededATPastheuniversalenergycurrency.
ThatwouldmakesensebecauseitmeansthatthesimplestformofATPsynthesis,substratelevelphosphorylation,wouldhaveprecededchemi-osmoticATPsynthesis,whichrequiresproteins.
Fromthatwouldfollowthattheearlyhistoryofbiologicalenergycon-servationisrecapitulatedduringeverysinglecatalyticcycleoftheGAPDHreactionintheoxidativedirection,wherebyinearlyenergyconservationthereductivesynthesisofthioestersfromH2andCO2andtheirconversiontoacylphosphateswouldhavebeenexergonic,withacouplingofenergymetab-olismtoautotrophy.
EnvoiEarlyevolutionwillalwaysholdadeepelementoffascination.
Afterall,itisalmostimpossibletoimaginehowanythingascomplicatedaslifecouldgetstartedfromanythingassimpleasCysSHCOHRaCysSCOHRHNAD+NADH+H+CysSCORHPO4=CysSCORHPOOO––OCoASCOCH3HPO4=COPOOO––ONiCCH3CH3+SSONiFeCoASHNiCOCH3SSNiFeCoASHNiCCH3SSONiFeNiSSNiFeCoCOb+electronselectronsCoFig.
4Energyconservationashigh-energyphosphatebonds.
aMechanismoftheGAPDHreactionintheoxidativedirection,redrawnafterSegalandBoyer(1953)andBieseckeretal.
(1977),R=CHOHCH2OPO32.
Theverticalarrowunderscorestheoxidativenatureofthereversiblereactionwhendrawnfromlefttoright.
Forsustainedfluxinthatdirectiontoallowsustainedsynthesisofhigh-energyphosphatebonds,suitableelectronacceptorsmustbeavailableintheenvironment.
Theseacceptorscouldnothavebeengeneratedfermentativelyduringmetabolismoforganicsfromspacebecauseor-ganicsfromspaceareunfermentablesubstrates(Schnheitetal.
2016).
Assumingthe(debateable)existenceofaprimordialsourceofsugars,oxidativeenergyconservationrequirestheexistenceofoxidantsaselec-tronacceptors;oxidantsarehoweverthermodynamicallyunfavorablefortheaccumulationofthebuildingblocksoflife(seetext).
Underphysio-logicalconditions,theacylphosphatebondintherightmostcompound,1,3-bisphosphoglycerate,hasafreeenergyofhydrolysisof52kJ/mol(Thaueretal.
1977).
bReductivesynthesisofacylphosphate(acetylphosphate)fromCOandamethylgroupasitoccursintheacetyl-CoApathwayofacetogensin(Fuchs2011;SchuchmannandMüller2014)andinsomemethanogenswhengrownonCO(RotherandMetcalf2004).
ThereactionsaredrawnfromdatacompiledinSvetlitchnaiaetal.
(2006)ANDinRagsdale(2009)particularlyFigs.
S5andS6,inFuchs(2011),andinSchuchmannandMüller(2014).
Theverticalarrowindicatesthattheexergonicnatureofthereactionsequence,henceitsabilityforsustainedsynthesisofacylphosphate,entailsthecontinuousreductionofCO2andhencerequirestheenvironmentalavailabilityofasuitablereductantsuchasH2,whichwasabundantontheearlyEarth(Sleepetal.
,2011)andisstillabundanttodayinhydrothermalvents(McCollomandSeewald2013).
ElectronsfromtheH2/H+couple,whichhasastandardmidpointpotentialof414kJ/molatpH7(Thaueretal.
1977),areusedbyhydrogenotrophicacetogensandmethanogenstosynthesizeCOandthemethylgroupfromCO2(Fuchs2011,SchuchmannandMüller2014).
Underphysiologicalconditions,theacylphosphatebondintheacetylphosphatehasafreeenergyofhydrolysisof45kJ/mol(Thaueretal.
1977).
Inmicrobialinmetabolism,both1,3-bisphosphoglycerateandacetylphosphatetypicallyphosphorylateADPtoATP,whichhasfreeenergyofhydrolysisof32kJ/mol(Thaueretal.
1977),viasubstratelevelphosphorylation.
Somereadersmightobjecttotheuseofthetermsenergy-richbondorhigh-energybond,butthetermsareveryusefulandthelabilityofthebondsinquestionisfoundedinthecircumstancethatthevalenceelectronsofPandSareinthethirdshell,whichcanacceptfurtherelectronsinthedorbital,andinthecircumstancethatcorrespondingbondshavesubstantialbondlengths,offeringampleopportunityforat-tackbymoleculessuchaswaterthatpossessloneelectronpairs(Wald1964)Physiology,phylogeny,earlyevolution,andGAPDH1831rocks,water,andCO2ontheearlyEarth.
Peoplelookatearlyevolutionfrommanydifferentangles,fromchemicaltracesforearlylifetoreplication,togenetics,toself-organization,andfinallytotheoriginsoffree-livingprokaryotesthatalmost2billionyearsagogaverisetoeukaryotes—ourancestors—andtruebiologicalcomplexityviaendosymbiosis.
Whatmanyob-serversfailtonoteisthatlifeisachemicalreaction,anenergy-releasingchemicalreaction.
Thesubstanceoflife,andweour-selves,areultimatelyjustsideproductsofthatreaction,whosemeaningistokeepthereactiongoingandwhoseprerogativeistopondertheoriginofitall.
AcknowledgementsWorkinthelaboratoryofRCwasfundedbytheDFG,andworkinthelaboratoryofWFMisfundedbytheERC.
WethankKlausV.
Kowallikformanydiscussionsandcriticalcommentsonthetext,andwethankVerenaZimorskiforthehelpinpreparingthemanuscript.
OpenAccessThisarticleisdistributedunderthetermsoftheCreativeCommonsAttribution4.
0InternationalLicense(http://creativecommons.
org/licenses/by/4.
0/),whichpermitsunrestricteduse,distribution,andreproductioninanymedium,providedyougiveappro-priatecredittotheoriginalauthor(s)andthesource,providealinktotheCreativeCommonslicense,andindicateifchangesweremade.
ReferencesAllenJF(2015)Whychloroplastsandmitochondriaretaintheirowngenomesandgeneticsystems:colocationforredoxregulationofgeneexpression.
ProcNatlAcadSciUSA112:10231–10238.
doi:10.
1073/pnas.
1500012112AmendJP,McCollomTM(2009)EnergeticsofbiomoleculesynthesisonearlyEarth.
In:ZaikowskiL,FriedrichJM,RussellSeidelS(eds)ChemicalevolutionII:fromtheoriginsoflifetomodernsociety.
AmericanChemicalSociety,Washington,DC,pp63–94ArchibaldJM(2014)Oneplusoneequalsone:symbiosisandtheevolu-tionofcomplexlife.
OxfordUniversityPress,Oxford,UKBieseckerG,HarrisJI,ThierryJC,WalkerJE,WonacottAJ(1977)SequenceandstructureofD-glyceraldehyde-3-phosphatedehydro-genasefromBacillusstearothermophilus.
Nature266:328–333BrinkmannH,MartinezP,QuigleyF,MartinW,CerffR(1987)Endosymbioticoriginandcodonbiasofthenucleargeneforchlo-roplastglyceraldehyde-3-phosphatedehydrogenasefrommaize.
JMolEvol26:320–328.
doi:10.
1007/BF02101150BrinkmannH,CerffR,SalomonM,SollJ(1989)CloningandsequenceanalysisofcDNAsencodingthecytosolicprecursorsofsubunitsGapAandGapBofchloroplastglyceraldehyde-3-phosphatedehy-drogenasesfrompeaandspinach.
PlantMolBiol13:81–94BuckelW,EggererH(1965)Ontheopticaldeterminationofcitratesyn-thaseandacetyl-coenzymea.
BiochemZ343:29–43CerffR(1974)Inhibitor-dependentreciprocalchangesintheactivitiesofglyceraldehyde3-phosphatedehydrogenasesinSinapisalbacotyle-dons.
ZPflanzenphysiol73:109–118CerffR(1978a)Glyceraldehyde-3-phosphatedehydrogenase(NADP)fromSinapisalbaL.
:reversibleassociationofenzymewithapro-teinfactorascontrolledbypyridinenucleotidesinvitro.
PlantPhysiol61:369–372.
doi:10.
1104/pp.
61.
3.
369CerffR(1978b)Glyceraldehyde-3-phosphatedehydrogenase(NADP)fromSinapisalbaL.
:steady-statekinetics.
Phytochemistry17:2061–2067.
doi:10.
1016/S0031-9422(00)89281-XCerffR(1979)Quaternarystructureofhigherplantglyceraldehyde-3-phosphatedehydrogenases.
EurJBiochem94:243–247.
doi:10.
1111/j.
1432-1033.
1979.
tb12891.
xCerffR(1982a)SeparationandpurificationofNAD-andNADP-linkedglyceraldehyde-3-phosphatedehydrogenasesfromhigherplants.
In:EdelmanM,HallickRB,ChuaN-H(eds)Methodsinchloroplastmolecularbiology.
ElsevierBiomedicalPress,Amsterdam,pp683–694CerffR(1982b)Evolutionarydivergenceofchloroplastandcytosolicglyceraldehyde-3-phosphatedehydrogenasesfromangiosperms.
EurJBiochem126:513–515.
doi:10.
1111/j.
1432-1033.
1982.
tb06810.
xCerffR,ChambersSE(1979)Subunitstructureofhigherplantglyceraldehyde-3-phosphatedehydrogenase(EC1.
2.
1.
12andEC1.
2.
1.
12).
JBiolChem254:6094–6098CerffR,KloppstechK(1982)Structuraldiversityanddifferentiallightcontrolofmessenger-RNAscodingforangiospermglyceraldehyde-3-phosphatedehydrogenases.
ProcNatlAcadSciUSA79:7624–7628.
doi:10.
1073/pnas.
79.
24.
7624CerffR,MartinW,BrinkmannH(1994)Originofintrons-earlyorlate.
Nature369:527–528CoriGT,SleinMW,CoriCF(1948)CrystallineD-glyceraldehyde-3-phosphatedehydrogenasefromrabbitmuscle.
JBiolChem173:605–618CottonJA,McInerneyJO(2010)Eukaryoticgenesofarchaebacterialoriginaremoreimportantthanthemorenumerouseubacterialgenes,irrespectiveoffunction.
ProcNatlAcadSciUSA107:17252–17255.
doi:10.
1073/pnas.
1000265107DayhoffMO(1965)Atlasofproteinsequenceandstructure.
NationalBiomedicalResearchFoundation,MarylanddeDuveC(1969)Evolutionoftheperoxisome.
AnnNYAcadSci168:369–381.
doi:10.
1111/j.
1749-6632.
1969.
tb43124.
xdeDuveC(1991)Blueprintforacell:thenatureandoriginoflife.
NeilPattersonPublishers,Burlington,NorthCarolinadeDuveC(2007)Theoriginofeukaryotes:areappraisal.
NatRevGenet8:395–403.
doi:10.
1038/nrg2071DoolittleWF(1998)Youarewhatyoueat:agenetransferratchetcouldaccountforbacterialgenesineukaryoticnucleargenomes.
TrendsGenet14:307–311EckRV,DayhoffMO(1966)Evolutionofthestructureofferredoxinbasedonlivingrelicsofprimitiveaminoacidsequences.
Science152:363–366EsserC,AhmadinejadN,WiegandC,RotteC,SebastianiF,Gelius-DietrichG,HenzeK,KretschmannE,RichlyE,LeisterD,BryantD,SteelMA,LockhartPJ,PennyD,MartinW(2004)Agenomephylogenyformitochondriaamongalpha-proteobacteriaandapre-dominantlyeubacterialancestryofyeastnucleargenes.
MolBiolEvol21:1643–1660.
doi:10.
1093/molbev/msh160FabryS,HenselR(1988)Primarystructureofglyceraldehyde-3-phosphatedehydrogenasededucedfromthenucleotidesequenceofthethermophilicarchaebacteriumMethanothermusfervidus.
Gene64:189–197.
doi:10.
1016/0378-1119(88)90334-4FenchelT,FinlayBJ(1995)Ecologyandevolutioninanoxicworlds.
OxfordUniversityPress,OxfordFitchWM,MargoliashE(1967)Constructionofphylogenetictrees.
Science155:279–284FuchsG(2011)Alternativepathwaysofcarbondioxidefixation:insightsintotheearlyevolutionoflifeAnnuRevMicrobiol65:631–658.
doi:10.
1146/annurev-micro-090110-102801GibbsM(1952)Triosephosphatedehydrogenaseandglucose-6-phosphatedehydrogenaseinthepeaplant.
Nature170:164–165.
doi:10.
1038/170164a0GibbsSP(1978)ThechloroplastsofEuglenamayhaveevolvedfromsymbioticgreenalgae.
CanJBot56:2883–2889GilbertW(1978)WhygenesinpiecesNature271:501.
doi:10.
1038/271501a01832W.
F.
Martin,R.
CerffGilbertW(1986)Originoflife:theRNAworld.
Nature319:618.
doi:10.
1038/319618a0GilbertW(1987)Theexontheoryofgenes.
ColdSpringHarbSympQuantBiol52:901–905GouldSB,WallerRF,McFaddenGI(2008)Plastidevolution.
AnnuRevPlantBiol59:491–517.
doi:10.
1146/annurev.
arplant.
59.
032607.
092915HabenichtA,QuesadaA,CerffR(1997)Sequenceofthenon-phosphorylatingglyceraldehyde-3-phosphatedehydrogenasefromNicotianaplumbaginifoliaandphylogeneticoriginofthegenefam-ily.
Gene198:237–243.
doi:10.
1016/S0378-1119(97)00320-XHaldaneJBS(1929)Theoriginoflife.
RationalistAnnual1929:3–10HenzeK,BadrA,WetternM,CerffR,MartinW(1995)AnucleargeneofeubacterialorigininEuglenagracilisreflectscrypticendosymbi-osesduringprotistevolution.
ProcNatlAcadSciUSA92:9122–9126.
doi:10.
1073/pnas.
92.
20.
9122HerrmannRG,BohnertHJ,KowallikKV,SchmittJM(1975)Size,con-formationandpurityofchloroplastDNAofhigherplants.
BiochimBiophysActa378:305–317.
doi:10.
1016/0005-2787(75)90118-5KersanachR,BrinkmannH,LiaudMF,ZhangDX,MartinW,CerffR(1994)Fiveidenticalintronpositionsinancientduplicatedgenesofeubacterialorigin.
Nature367:387–389.
doi:10.
1038/367387a0KooninEV(2015)Originofeukaryotesfromwithinarchaea,archaealeukaryome,andburstsofgenegain:eukaryogenesisjustmadeeas-ierPhilTransRSocLondBBiolSci370:20140333.
doi:10.
1098/rstb.
2014.
0333KowallikKV,HaberkornG(1971)TheDNA-structuresofthechloro-plastofProrocentrummicans(Dinophyceae).
ArchMicrobiol80:252–261KuC,Nelson-SathiS,RoettgerM,SousaFL,LockhartPJ,BryantD,Hazkani-CovoE,McInerneyJO,LandanG,MartinWF(2015)Endosymbioticoriginanddifferentiallossofeukaryoticgenes.
Nature524:427–432.
doi:10.
1038/nature14963LeeRE(1972)Originofplastidsandthephylogenyofalgae.
Nature237:44–46LeeRE(1977)Evolutionofalgalflagellateswithchloroplastendoplas-micreticulumfromtheciliates.
SAfrJSci73:179–182LiaudMF,ZhangDX,CerffR(1990)Differentialintronlossandendo-symbiotictransferofchloroplastglyceraldehyde-3-phosphatedehy-drogenasegenestothenucleus.
ProcNatlAcadSciUSA87:8918–8922LiaudMF,ValentinC,MartinW,BougetFY,KloaregB,CerffR(1994)Theevolutionaryoriginofredalgaeasdeducedfromthenucleargenesencodingcytosolicandchloroplastglyceraldehyde-3-phosphatedehydrogenasesfromChondruscrispus.
JMolEvol38:319–327.
doi:10.
1007/BF00163149LiaudMF,BrandtU,ScherzingerM,CerffR(1997)Evolutionaryoriginofcryptomonadmicroalgae:twonovelchloroplast/cytosol-specificGAPDHgenesaspotentialmarkersofancestralendosymbiontandhostcellcomponents.
JMolEvol44:S28–S37.
doi:10.
1007/PL00000050LiaudMF,LichtléC,AptK,MartinW,CerffR(2000)Compartment-specificisoformsofTPIandGAPDHareimportedintodiatommitochondriaasafusionprotein:evidenceinfavorofamitochon-drialoriginoftheeukaryoticglycolyticpathway.
MolBiolEvol17:213–223LindmarkDG,MüllerM(1973)Hydrogenosome,acytoplasmicorgan-elleoftheanaerobicflagellateTritrichomonasfoetus,anditsroleinpyruvatemetabolism.
JBiolChem248:7724–7728LipmannF(1941)Metabolicgenerationandutilizationofphosphatebondenergy.
AdvEnzymol1:99–162LongM,DeSouzaS,RosenbergC,GilbertW(1996)Exonshufflingandtheoriginofthemitochondrialtargetingfunctioninplantcyto-chromec1precusor.
ProcNatlAcadSciUSA93:7727–7731MadiganMT,MartinkoJM,BenderKS,BuckleyDH,StahlDA(2012)Brockbiologyofmicroorganisms,13thedn.
BenjaminCummings,BostonMargulisL,ChapmanM,GuerreroR,HallJ(2006)Thelasteukaryoticcommonancestor(LECA):acquisitionofcytoskeletalmotilityfromaerotolerantspirochetesintheProterozoicEon.
ProcNatlAcadSciUSA103:13080–13085.
doi:10.
1073/pnas.
0604985103MartinW(1985)Klonierung,IdentifizierungundSequenzierungvoncDNAsfürNAD+-undNADP+-spezifischeGAPDHsausSinapisalba.
DiplomarbeitamFachbereichBiologiederTechnischenUniversittHanoverunterderAnleitungvonR.
CerffMartinWF(1996)IssomethingwrongwiththetreeoflifeBioEssays18:523–527.
doi:10.
1002/bies.
950180702MartinW(2010)Evolutionaryoriginsofmetaboliccompartmentaliza-tionineukaryotes.
PhilosTransRSocLondB365:847–855.
doi:10.
1098/rstb.
2009.
0252MartinWF(2012)Hydrogen,metals,bifurcatingelectrons,andprotongradients:theearlyevolutionofbiologicalenergyconservation.
FEBSLett9:485–493MartinW,CerffR(1986)Prokaryoticfeaturesofanucleus-encodedenzyme:cDNAsequencesforchloroplastandcytosolicglyceraldehyde-3-phosphatedehydrogenasesfrommustard(Sinapisalba).
EurJBiochem159:323–331.
doi:10.
1111/j.
1432-1033.
1986.
tb09871.
xMartinW,MüllerM(1998)Thehydrogenhypothesisforthefirsteukary-ote.
Nature392:37–41.
doi:10.
1038/32096MartinW,BrinkmannH,SavonaC,CerffR(1993)Evidenceforachi-mericnatureofnucleargenomes:eubacterialoriginofeukaryoticglyceraldehyde-3-phosphatedehydrogenasegenes.
ProcNatlAcadSciUSA90:8692–8696.
doi:10.
1073/pnas.
90.
18.
8692MartinW,GargS,ZimorskiV(2015)Endosymbiotictheoriesforeu-karyoteorigin.
PhilosTransRSocLondB370:20140330.
doi:10.
1098/rstb.
2014.
0330McCollomTM,SeewaldJS(2013)Serpentinites,hydrogen,andlife.
Elements9:129–134.
doi:10.
2113/gselements.
9.
2.
129McInerneyJO,O'ConnellM,PisaniD(2014)ThehybridnatureoftheEukaryotaandaconsilientviewoflifeonEarth.
NatRevMicrobiol12:449–455.
doi:10.
1038/nrmicro3271MelandriBA,PupilloP,Baccarini-MelandriA(1970)D-glyceraldehyde-3-phosphatedehydrogenaseinphotosyntheticcells.
I.
Thereversiblelight-inducedactivationinvivoofNADP-dependentenzymeanditsrelationshiptoNAD-dependentactivities.
BiochimBiophysActa220:178–189.
doi:10.
1016/0005-2744(70)90004-5MereschkowskyC(1905)berNaturundUrsprungderChromatophorenimPflanzenreiche.
BiolCentralbl25:593–604[Englishtranslationin:MartinW,KowallikK(1999)AnnotatedEnglishtranslationofMereschkowsky's1905paper.
EurJPhycol34:287–295]Meyer-GauenG,SchnarrenbergerC,CerffR,MartinW(1994)Molecularcharacterizationofanovel,nuclear-encoded,NAD+-de-pendentglyceraldehyde-3-phosphatedehydrogenaseinplastidsofthegymnospermPinussylvestrisL.
PlantMolBiol26:1155–1166.
doi:10.
1007/BF00040696MeyerhofO(1951)Phosphorusmetabolism.
AmSci39:682–687MichelsPAM,OpperdoesFR(1991)Theevolutionaryoriginofglycosomes.
ParasitolToday7:105–109.
doi:10.
1016/0169-4758(91)90167-MMüllerM(1988)Energymetabolismofprotozoawithoutmitochondria.
AnnuRevMicrobiol42:465–488.
doi:10.
1146/annurev.
mi.
42.
100188.
002341MüllerM,MentelM,vanHellemondJJ,HenzeK,WoehleC,GouldSB,YuRY,vanderGiezenM,TielensAGM,MartinWF(2012)Biochemistryandevolutionofanaerobicenergymetabolismineu-karyotes.
MicrobiolMolBiolRev76:444–495.
doi:10.
1128/MMBR.
05024-11Physiology,phylogeny,earlyevolution,andGAPDH1833NeedhamDM,PillaiRK(1937)Thecouplingofoxido-reductionsanddismutationswithesterificationofphosphateinmuscle.
BiochemJ31:1837–1851PetersenJ,BrinkmannH,CerffR(2003)Origin,evolution,andmetabol-icroleofanovelglycolyticGAPDHenzymerecruitedbylandplantplastids.
JMolEvol57:16–26.
doi:10.
1007/s00239-002-2441-yPetersenJ,TeichR,BeckerB,CerffR,BrinkmannH(2006a)TheGapA/Bgeneduplicationmarkstheoriginofstreptophyta(Charophytesandlandplants).
MolBiolEvol23:1109–1118.
doi:10.
1093/molbev/msj123PetersenJ,TeichR,BrinkmannH,CerffR(2006b)ABgreen^phosphoribulokinaseincomplexalgaewithredplastids:evidenceforasinglesecondaryendosymbiosisleadingtohaptophytes,cryptophytes,heterokonts,anddinoflagellates.
JMolEvol62:143–157.
doi:10.
1007/s00239-004-0305-3PisaniD,CottonJA,McInerneyJO(2007)Supertreesdisentanglethechimericaloriginofeukaryoticgenomes.
MolBiolEvol24:1752–1760.
doi:10.
1093/molbev/msm095QuigleyF,MartinW,CerffR(1988)Intronconservationacrosstheprokaryote-eukaryoteboundry:structureofthenucleargeneforchloroplastglyceraldehyde-3-phosphatedehydrogenasefrommaize.
ProcNatlAcadSciUSA85:2672–2676RagsdaleSW(2009)Nickel-basedenzymesystems.
JBiolChem284:18571–18575RotherM,MetcalfWW(2004)AnaerobicgrowthofMethanosarcinaacetivoransC2Aoncarbonmonoxide:anunusualwayoflifeforamethanogenicarchaeon.
ProcNatlAcadSciUSA101:16929–16934.
doi:10.
1073/pnas.
0407486101SaganL(1967)Ontheoriginofmitosingcells.
JTheoretBiol14:225–274.
doi:10.
1016/0022-5193(67)90079-3SchnheitP,BuckelW,MartinWF(2016)Ontheoriginofheterotrophy.
TrendsMicrobiol24:12–25.
doi:10.
1016/j.
tim.
2015.
10.
003SchuchmannK,MüllerV(2014)Autotrophyatthethermodynamiclimitoflife:amodelforenergyconservationinacetogenicbacteria.
NatRevMicrobiol12:809–821.
doi:10.
1038/nrmicro3365SchwartzRM,DayhoffMO(1978)Originsofprokaryotes,eukaryotes,mitochondria,andchloroplasts.
Science199:395–403.
doi:10.
1126/science.
202030SegalHL,BoyerPD(1953)TheroleofsulfhydrylgroupsintheactivityofD-glyceraldehyde-3-phosphatedehydrogenase.
JBiolChem204:265–281SephtonMA(2002)Organiccompoundsincarbonaceousmeteorites.
NatProdRep19:292–311.
doi:10.
1039/b103775gShockEL,BoydES(2015)Geomicrobiologyandmicrobialgeochemis-try:principlesofgeobiochemistry.
Elements11:389–394.
doi:10.
2113/gselements.
11.
6.
395SitteP(2007)EndosymbioseinderZell-undLebensevolution.
In:StorchV,WelschU,WinkM(eds)Evolutionsbiologie,2.
Auflage.
Springer,Berlin,Heidelberg,NewYork,pp227–238SleepNH,MeibomA,FridrikssonT,ColemanRG,BirdDK(2004)H2-richfluidsfromserpentinization:geochemicalandbioticimplica-tions.
ProcNatlAcadSciUSA101:12818–12823.
doi:10.
1073/pnas.
0405289101SleepNH,BirdDK,PopeEC(2011)Serpentiniteandthedawnoflife.
PhilosTransRSocLondSerBBiolSci366:2857–2869.
doi:10.
1098/rstb.
2011.
0129SollJ,SchleiffE(2004)Proteinimportintochloroplasts.
NatRevMolCellBiol5:198–208.
doi:10.
1038/nrm1333SousaFL,ThiergartT,LandanG,Nelson-SathiS,PereiraIAC,AllenJF,LaneN,MartinWF(2013)Earlybioenergeticevolution.
PhilosTransRSocLondB368:20130088.
doi:10.
1098/rstb.
2013.
0088StoebeB,MaierUG(2002)One,two,three:nature'stoolboxforbuildingplastids.
Protoplasma219:123–130.
doi:10.
1007/s007090200013StumpfPK(1950)Carbohydratemetabolisminhigherplants.
III.
Breakdownoffructosediphosphatebypeaextracts.
JBiolChem182:261–272SvetlitchnaiaT,SvetlitchnyiV,MeyerO,DobbekH(2006)Structuralinsightsintomethyltransferreactionsofacorrinoidiron-sulfurpro-teininvolvedinacetyl-CoAsynthesis.
ProcNatlAcadSciUSA103:14331–14336ThauerRK,JungermannK,DeckerK(1977)Energyconservationinchemotrophicanaerobicbacteria.
BacteriolRev41:100–180WaldG(1964)Theoriginsoflife.
ProcNatlAcadSciUSA52:595–611WallinIE(1925)Onthenatureofmitochondria.
IX.
Demonstrationofthebacterialnatureofmitochondria.
AmJAnat36:131–139.
doi:10.
1002/aja.
1000360106WarburgO,ChristianW(1939)IsolierungundKrystallisationdesProteinsdesoxydierendenGarungsferments.
BiochemZ303:40–68WedelN,SollJ(1998)EvolutionaryconservedlightregulationofCalvincycleactivitybyNADPH-mediatedreversiblephosphoribulokinase/CP12/glyceraldehyde-3-phosphatedehydro-genasecomplexdissociation.
ProcNatlAcadSciUSA95:9699–9704WedelN,SollJ,PaapB(1997)CP12providesanewmodeoflightregulationofCalvincycleactivityinhigherplants.
ProcNatlAcadSciUSA94:10479–10484WeedenNF(1981)Geneticandbiochemicalimplicationsoftheendo-symbioticoriginofthechloroplast.
JMolEvol17:133–139.
doi:10.
1007/BF01733906WeissMC,SousaFL,MrnjavacN,NeukirchenS,RttgerM,Nelson-SathiS,MartinWF.
Thephysiologyandhabitatofthelastuniversalcommonancestor.
NatureMicrobiology1:16116(2016)WilliamsTA,FosterPG,CoxCJ,EmbleyTM(2013)Anarchaealoriginofeukaryotessupportsonlytwoprimarydomainsoflife.
Nature504:231–236.
doi:10.
1038/nature12779WoeseCR,KandlerO,WheelisML(1990)Towardsanaturalsystemoforganisms:proposalforthedomainsarchaea,bacteria,andEucarya.
ProcNatlAcadSciUSA87:4576–4579.
doi:10.
1073/pnas.
87.
12.
4576Zaremba-NiedzwiedzkaK,CaceresEF,SawJH,BckstrmD,JuzokaiteL,VancaesterE,SeitzKW,AnantharamanK,StarnawskiP,KjeldsenKU,StottMB,NunouraT,BanfieldJF,SchrammA,BakerBJ,SpangA,EttemaTJG(2017)Asgardarchaeailluminatetheoriginofeukaryoticcellularcomplexity.
Nature541:353–358.
doi:10.
1038/nature21031ZimorskiV,KuC,MartinWF,GouldSB(2014)Endosymbiotictheoryfororganelleorigins.
CurrOpinMicrobiol22:38–48.
doi:10.
1016/j.
mib.
2014.
09.
008ZwicklP,FabryS,BogedianC,HaasA,HenselR(1990)Glyceraldehyde-3-phosphatedehydrogenasefromthehyperthermo-philicarchaebacteriumPyrococcuswoesei:characterisationoftheenzyme,cloningandsequencingofthegene,andexpressioninEscherichiacoli.
JBacteriol172:4329–43381834W.
F.
Martin,R.
Cerff
Martin1&RüdigerCerff2Received:7December2016/Accepted:22February2017/Publishedonline:6March2017#TheAuthor(s)2017.
ThisarticleispublishedwithopenaccessatSpringerlink.
comAbstractThechloroplastandcytosolofplantcellsharboranumberofparallelbiochemicalreactionsgermanetotheCalvincycleandglycolysis,respectively.
Thesereactionsarecatalyzedbynuclearencoded,compartment-specificiso-enzymesthatdifferintheirphysiochemicalproperties.
ThechloroplastcytosolisoenzymesofD-glyceraldehyde-3-phos-phatedehydrogenase(GAPDH)harborevidenceofmajoreventsinthehistoryoflife:theoriginofthefirstgenes,thebacterial-archaealsplit,theoriginofeukaryotes,theevolutionofproteincompartmentationduringeukaryoteevolution,theoriginofplastids,andthesecondaryendosymbiosisamongthealgaewithcomplexplastids.
ThereactionmechanismofGAPDHentailsphosphorolysisofathioestertoyieldanenergy-richacylphosphatebond,achemistrythatpointstoprimitivepathwaysofenergyconservationthatexistedevenbeforetheoriginofthefirstfree-livingcells.
Here,werecountthemaininsightsthatchloroplastandcytosolicGAPDHpro-videdintoendosymbiosisandphysiologicalevolution.
KeywordsEndosymbiosis.
Plastids.
Mitochondria.
Cellevolution.
PeterSitteIntroductionPeterSittewasavirtuosointheartofelectronmicroscopy.
Hedevotedhisscientificcareertounderstandingthenatureandevolutionarybasisofcompartmentationineukaryoticcellsandtherolethatendosymbiosisplayedtherein(Sitte2007).
Thanksmainlytoelectronmicroscopicstudiesinthe1960sand1970s,scientistsin2016recognizetwokindsofcells:theprokaryotictypeandtheeukaryotictype.
Themaindifferencethatdistinguishesthetwocelltypesisthenatureofinternalcompartmentationineukaryotes.
Thechromosomesineu-karyoticcellsareseparatedfromthecytoplasmbymembranesurroundingthecellnucleus,whilechromosomesinprokary-otesarefreelydispersedthroughoutinthecytoplasm.
Eukaryotestypicallypossessacomplexendomembranesys-tem,andmitochondria,plant,andalgalcellspossesschloro-plastsinaddition.
Bythemeasureofcompartmentation,themostcomplexcellsinnaturearefoundamongthealgaethatpossessplastidssurroundedbythreeorfourmembranes,plas-tidsthatareremnantsofevolutionarilyreducedeukaryoticcellsresidingwithinthecytosolofanothernucleus-bearingcell(StoebeandMaier2002;Gouldetal.
2008).
Thoughitwasnotalwaysthecase,today,biologistsrecognizethatcom-plexityineukaryoticcellsstemsfromendosymbiosis(Archibald2014).
EndosymbiotictheorytakesrootinMereschkowsky'sclas-sicalessayontheoriginofplastids(Mereschkowsky1905).
Ithasalongandturbulenthistory,asrecentlysummarizedelse-where(Martinetal.
2015).
Theelderofusfirstlearnedaboutendosymbiosisinthe1960sinPeterSitte'scellbiologylec-turesattheUniversityofFreiburg.
Endosymbiotictheory—theprospectthatmitochondriaandchloroplastsdescendedfromfreelivingprokaryotesthatenteredintoasymbioticre-lationshipwiththeirrespectivehostcellearlyineukaryotichistory—wasaveryexciting,almostrevolutionary,prospectThispaperisdedicatedtothememoryofPeterSitte(1929–2015).
HandlingEditor:UliKutschera*WilliamF.
Martinbill@hhu.
deRüdigerCerffcerff-ruediger@t-online.
de1InstituteofMolecularEvolution,UniversityofDüsseldorf,Universittsstr.
1,40225Düsseldorf,Germany2InstituteofGenetics,TechnicalUniversityofBraunschweig,Spielmannstr.
7,38106Braunschweig,GermanyProtoplasma(2017)254:1823–1834DOI10.
1007/s00709-017-1095-yincellevolutionthatopenedupfundamentallynewavenuesofpursuittoinvestigateandunderstandeukaryoticintracellu-larcompartmentation.
Oneaspectinparticularwasimportantforendosymbiotictheory:thecompartmentationofmetabo-lismineukaryotes.
Earlyon,endosymbiotictheoryhaditthatthecoremetabolicfunctionsofmitochondria(respiration)andchloroplasts(photosynthesis)weredirectinheritancesfromthebacterialancestorsoforganelles.
ItwasalsoclearfromelectronmicroscopythatorganellespossessedDNA(KowallikandHaberkorn1971),andthatorganellegenomesweremuchtoosmalltoencodealloftheproteinsthatunder-pinrespirationandphotosynthesis(Herrmannetal.
1975).
Asaconsequence,mostoftheproteinsthatsupportthephysio-logicalfunctionofchloroplastsandmitochondriahadtobeencodedinnuclearchromosomes,whichmeantthattherehadtohavebeensomeformofgenetransfergoingonfromendo-symbiontstothehost,orasWallinputitwithregardtomito-chondria,B.
.
.
bacterialorganismsmaydevelopanabsolutesymbiosiswithahigherorganismandinsomewayoranotherimpressanewcharacteronthefactorsofheredity.
Thesim-plestandmostreadilyconceivablemechanismbywhichthealterationtakesplacewouldbetheadditionofnewgenestothechromosomesfromthebacterialsymbiont.
^(Wallin1925;p.
144).
Chloroplastcytosolisoenzymesprovideduniqueopportu-nitiestotestcrucialpredictionsofendosymbiotictheorywithmolecularevolutionarystudies.
Ifendosymbiotictheorywascorrectaboutchloroplastsarisingfromcyanobacteria,itfollowedthatthenucleargeneforthechloroplastenzymeshouldultimatelystemfromthegenomeofthecyanobacterialancestorofplastids,whilethecytosolicenzymeshouldreflecttheevolutionofthehostcellthatacquiredtheplastid.
Asitappliedtoisoenzymes,thisspecificinferencewascalledtheproductspecificitycorollarytoendosymbiotictheory(Weeden1981).
Chloroplastcytosolisoenzymesforhigherplantglyceraldehyde-3-phosphatedehydrogenases(GAPDH)providedanexcellentsystemtotesttheprospectofgenetransferinendosymbiosis.
GAPDH:fromproteintoDNALikeendosymbiotictheory,GAPDHitselfhasalongandturbulenthistory.
TheenzymaticactivityofGAPDHgoeswayback,bothinplantmetabolismandinenzymologyitself.
Indeed,itwasamongthefirstenzymestobestudied.
EarlyworkonGAPDHfocusedontheenzymefrommuscleandyeast.
Inthe1930s,whenEmbden,Meyerhof,andParnasweremakingheadwayintosynthesisofadenylpyrophosphate(nowcalledATP)fromtheoxidationofglucose,DorothyNeedhamreportedthattheenergyreleasedintheoxidationoftriosephosphateindicatedBTheexistenceinmuscleextractofacoupledmechanismwherebysynthesisofadenylpyrophosphate(fromadenylicacidandfreephosphate)canaccompanyoxido-reduction^(NeedhamandPillai1937,p.
1850).
OttoWarburgreportedthecrystallizationoftheyeastenzyme(WarburgandChristian1939),whichhecalleddasoxydierendeGrungsferment.
BythetimethatCorietal.
(1948)reportedcrystallizationoftheanimalenzyme,itwascalledD-glyceraldehye-3-phosphatedehydrogenase.
Stumpf(1950)reportedGAPDHactivityinplants,notingthattheoxidationoffructose-1,6-bisphosphateinpeaseedlingsoccursbyaseriesofreactionthatBisapparentlysimilartothatinyeastandanimaltissues.
^AGAPDHactivitywasreportedinchlorophyll-containingplanttissuesthatrequiredNADPH(Gibbs1952).
Intothe1970s,itwasnotclearwhetheroneGAPDHenzymeexistedinplantsthatwasmodifiedforNADPH-dependentactivity(Melandrietal.
1970;Cerff1974)orwhetherisoenzymesexistedfortheNADH-andNADPH-dependentactivitiesand,ifthelattercaseweretrue,whethersuchisoenzymeswerespecifictotheGAPDHreactionsofglycolysisinthecytosolandtheCalvincycleofplastids.
Classicalenzymology—sep-aration,purification,andcharacterizationoftheplantenzymes(Cerff1978a,b;Cerff1982a)—demonstratedtheexistenceofcytosol-specific(NADHdependent)andchloroplast-specific(NADPHaccepting)GAPDHisoenzymesinphotosynthetictissuesofhigherplants(Cerff1979;CerffandChambers1979).
Theisoenzymesindeedhadverydistinctevolutionaryhistories(Melandrietal.
1970;Cerff1982a),andtheywereencodedbynucleargenes(Cerff1982b;CerffandKloppstech1982).
Intheearly1980s,therewasalotofexcitementsur-roundingthenewlydiscoveredabilityofmolecularse-quencecomparisonsandphylogenetictrees,notonlytotestendosymbiotictheorybuttoalsoreconstructearlyevolutionmoregenerally.
LynnMargulishadveryeffectivelyrevivedendosymbiotictheory(Sagan1967),butshenevermadethetransitiontotestingitspredictionswithmolecularphyloge-netics.
ItwasMargaretDayhoff(SchwartzandDayhoff1978)whousheredendosymbiotictheoryintotheeraofscientifictestingwithphylogenetictreescreatedfrompro-teinsequences.
Disappointingly,sequencesfortheplantGAPDHisoenzymescouldnotbeobtainedviastandardEdmandegradationproteinsequencingtechnologiesthathadworkedsowellforcytochromecorferredoxins(Dayhoff1965).
ThatmeantthatifonewantedtogettheaminoacidsequencesforplantGAPDHinordertoaddressthedecisiveevolutionaryissuesattheforefrontofthefield,onehadtoutilizethelatesttechnology:DNAsequencing.
Atthattime,theroadtoobtainingproteinsequencesfromDNAsequenceswentthroughcDNAsequencesandanti-bodies.
Methodsforseparatingandpurifyingtheisoen-zymes(Cerff1979)meantthatantiseraagainstthepurifiedproteinswereavailable(CerffandKloppstech1982).
Theavailabilityofantiserapermitteduseofanarchaic,experi-mentallydemanding,butofteneffectivetechniquecalled1824W.
F.
Martin,R.
Cerffhybridreleasetranslation.
Ifallhasgonewell,theresultofthehybridtranslationapproachtocloningdeliversthede-siredcDNAclones,andthechemicalmethodofMaxamandGilbertdeliverstheirsequences(MartinandCerff1986).
TheoriginofplastidsWiththecDNAsandderivedaminoacidsequences,wewereabletoshowthatthenuclearencodedchloroplastenzymewasmoresimilartoitshomologuesfrombacteriathanitwastohomologuesfromeukaryotes,andthatthenuclearencodedcytosolicenzymewasmoresimilartohomologuesfromani-malsandyeastthanitwastohomologuesfromprokaryotes(Fig.
1a).
Thisclearlyboreoutthepredictionsfromendosym-biotictheory,anovelandexcitingfind.
Intheprocessofnotgettingourpaperpublishedintwojournals,however,these-quencesofGAPDHfromEscherichiacolibecamepublished,andreferees,oneintimatelyfamiliarwithE.
coliGAPDH,weresuddenlydemandingthatweexplainwhyE.
coliGAPDHwasmoresimilartoeukaryoticsequencesthanitwastoGAPDHfromThermusaquaticusorBacillusstearothermophilus.
Wereasonedthatthiswasalsoprobablyacaseoflateralgenetransfer(ourworkinghypothesisentailedlateralgenetransferanyway),butnotfromprokaryotestoeukaryotesasinthecaseofthechloroplastenzyme,ratherfromeukaryotestoprokaryotes(MartinandCerff1986).
GAPDHthuspresentsoneoftheearliestreportsinthelitera-tureforsequence-basedinferenceoflateralgenetransfer,aconceptthatadecadelaterbecamequitepopular(toopopularinsomecirclesperhaps),andsequencesfromotherplantsourcesboreoutthepredictionsofendosymbiotictheorywithrespecttotheplantenzyme(Brinkmannetal.
1987).
ChloroplastGAPDHuncoveredadditionalsurprises.
TheNADPH-utilizingplastidenzymefromhigherplantswasknowntoexistintwoforms,anA4homotetramerandanA2B2heterodimer(CerffandChambers1979).
TheAandBsubunitswereshowntobetheresultofanucleargeneduplicationthattookplaceearlyintheevolutionofthegreenplantlineage,withtheBsubunithavingacquiredashortC-terminalextensionwithconservedcysteineresidues(Brinkmannetal.
1989).
TheC-terminalextensionofGapBwasacquiredatthebeginningoflandplantevolutionfromthenuclearencodedsmallredoxactiveproteinCP12,whichwasshowntointeractwiththeA2B2andA4formsofchlo-roplastGAPDHinadditiontophophoribulokinaseintheabsenceofNADP(H)(Wedeletal.
1997,WedelandSoll1998,Petersenetal.
2006a).
ThisinteractionblockedCO2fixationactivityinthedarkandpreventedfutilecyclingbetweenglycolysisandtheCalvincycle.
ItalsoexplainedwhythechloroplastenzymeaggregatedinthepresenceofNAD(H),whichwasthekeytoefficientseparationandpu-rificationoftheisoenzymes(Cerff1982a).
TheoriginofthefirstgenesIntheearly1980s,beforetheconceptofanRNAworld(Gilbert1986)hadbeenborn,peoplewerestillvigorouslydebatingtheissueofwhatcamefirst,protein,orDNA.
Oneofthebigpuzzleswashowthefirstlongopenreadingframesingenescametobe,andhowenzymesizedproteinsaroseintheabsenceofaccuratetemplatereplication.
Figuringpromi-nentlyinthatdebatewasWalterGilbert'sexontheoryofgenes(Gilbert1987),accordingtowhichintronswererelictsfromtheprimordialassemblyofgenesatlife'soriginandthatintronpositionsinmoderngenescorrespondedtotheboundariesbetweenstructuralmodulesofproteinfunctioncalleddo-mains.
Modules,beingshorterandeasiertoevolve,couldrecombineviaBexonshuffling^andexonscouldperhapsun-dergoalternativesplicing(Gilbert1978),therebypromotingancientenzymediversity.
AncientenzymessuchasGAPDHClostridiumpasteurianumaThermusaquaticusBacillusstearothermophilusAnabaenavariabilismaizepeatobaccomaizepeatobaccoGapAGapBChloroplastCyanobacteriumbFig.
1PhylogenetictreesofGAPDHaminoacidsequences.
aThetreewasconstructedbythemethodofFitchandMargoliash(1967)withtwointeractiveroundsoftopologicaladjustmenttoreducethesumofsquareddifferencesbetween(i)pairwisedistancescalculatedbetweenaminoacidssequencesusingtheminimummutationdistanceofDayhoff(1965)and(ii)pairwisedistancesmeasuredintheconstructedphyloge-netictree(Martin1985).
Senf,mustard;Hefe,yeast;Huhn,chicken;Mensch,human;Schwein,pig;Hummer,lobster;B.
,Bacillus,T.
,Thermus.
Numbersatbranchesaredistances.
bThenuclearencodedgenesfortheAandBsubunitsofhigherplantchloroplastGAPDH,anA2B2tetramer,branchwiththeCalvincyclehomologuefromcyanobacteria.
RedrawnfromMartinetal.
(1993)Physiology,phylogeny,earlyevolution,andGAPDH1825wereclearlywellsuitedtotestthoseideas,andindeed,wefoundintronpositionsthatwerepresentbetweenthesamenucleotidesinthesamehomologouscodoninthenucleargeneforchloroplastGAPDHandinanimalGAPDH(Quigleyetal.
1988).
ThecloserwelookedintoGAPDHgenes,themoreevidencewefoundforidenticalintronpositionsinancientlydivergedgenes(Liaudetal.
1990;Kersanachetal.
1994;Cerffetal.
1994).
Duringthoseinvestigations,however,ourviewsconcerningtheageofeukaryoticGAPDHgeneshadtoberevised,andlikesomanyotherthingsinevolution,cyanobacteriawereresponsibleforthechange.
EndosymbiosisandtheunexpectedoriginsofeukaryoticgenesTheendosymbiosisstoryfortheoriginofchloroplastGAPDHwasconceptuallysatisfying,butanimportantpiecewasmiss-ing.
Thepredictionfromendosymbiotictheorywas,namelythattheplastidenzymeshouldbemoresimilarnotjusttoprokaryotichomologuesingeneral,buttocyanobacterialho-mologuesinparticular.
Therefore,weembarkedtoobtainthesequenceofGAPDHfromcyanobacteria,whichwedid,find-ingmorethanweexpected.
Wefoundthatthecyanobacteriumthatwehadinvestigated,Anabaena,indeedhadaCalvincyclehomologueofGAPDHthat,inphylogenetictrees,branchedveryspecificallywiththenuclearencodedchloroplastGAPDHenzymesofhigherplants(Fig.
1b),aresoundingconfirmationofthepredictionfromendosymbiotictheory(Martinetal.
1993).
However,thatwasnotallthatAnabaenahadinstore,italsoharboredtwootherGAPDHgenes,oneofwhichsheddramaticlightonthepeskyE.
coliGAPDHthatwasannoyinglysimilartoeukaryoticGAPDH.
AnabaenahadacopyofthesameE.
coliGAPDHgenethat,upuntilthen,wehadthoughttobeaeukaryote-to-prokaryotetransfer.
Wehadbecomeprettygoodatinterpretingtrees,butthisobservationwasapuzzle.
Inordertomakesenseofit,wehadtopaycarefulattention(i)towhatReinhardHenselandhisteamhadbeenfindingforarchaealGAPDH(FabryandHensel1988;Zwickletal.
1990)and(ii)towhateveryonewassayingatthetimewithregardtoeukaryoteorigins.
Intheearly1990s,WoeseandKandler'srootedthreedomaintreewasasenvogueasitgetsitbiology(Woeseetal.
1990).
However,itwasatreeofribosomes.
Theassumptionbackthen,beforewehadgenomes,wasthattherRNAtreewasspeakingforthegenomeasawhole,andthatultimately,whenwehadallthedata,allgeneswouldtendtopaintroughlythesamepictureastherRNAtree,witharchaeaasthesisterstotheeukaryotesandwithmitochondrion-lackingeukaryotesbranchingearlyontheeukaryoticbranch,meaningthatmito-chondriacamelateineukaryoteevolutionandthateukaryotesare,forallpracticalpurposestobeseenasgrown-uparchaea.
Accordingly,eukaryoteGAPDHgenesshouldderivefromarchaealGAPDHgenes.
Thatwasnot,however,whatReinhardHenselwasfinding(FabryandHensel1988;Zwickletal.
1990).
TheywerefindingthatarchaealGAPDHwas,forallpracticalpurposes,unrelatedtoeukaryoteGAPDH,butitwasalsojustasunrelatedtobacterialGAPDH.
ThatindicatedasplitofarchaealGAPDHfromitshomo-loguesintheothertwodomainsattheearlieststagesofproteinevolution,whichinitselfwasnothugelysurprising.
However,fromourstandpoint,tryingtomakesenseofdatathatwasfundamentallydifferentfromwhateveryoneelsewasfinding(Fig.
2),therewasasubstantialsurprise.
Ifwethoughtthingsthroughinfull(whichissomethingwespentalotoftimedoinginthosedays),italsoindicatedthatalloftheGAPDHgenesthatwehadseenineukaryotesupuntilthenwere—justlikechloroplastGAPDH—acquiredfrombacteria.
Eukaryoticgenomeswerenotgenerallyarchaeal,theywerechimeric.
Becauseanimalsandfungihadneverpossessedplastids(sosaystheguidinglogicofendosymbiotictheory),thisledustopositthattheeukaryoticgeneforglycolyticGAPDHisalsoanacquisitionviaendosymbiosisandultimatelystemsfrommito-chondrion.
CytosolicGAPDH,acoreenzymeofcarbonandenergymetabolismandthekeyenergy-conservingstepofgly-colysis,reflectedtheevolutionofthehostthatacquiredtheplastid,butnottheevolutionofthehostthatacquiredthemi-tochondrion.
Endosymbiosishadmoretodowithcompartmen-talizedenergymetabolismineukaryotesthananyonehadsuspected.
Weconcludedbysaying:BTheevidenceforeubac-terialoriginofeukaryoticGAPC,anessentialglycolyticen-zyme,suggeststhateukaryoticgenomesaremorehighlychi-mericthanpreviouslyassumed.
Whereasmostorganellarpro-teinsarecurrentlyencodedinthenucleus,endosymbiontsmayhavedonatedmanygenestothenucleuswithoutorganellarreimportoftheprotein,therebyenrichingthegeneticandmet-abolicpotentialofthehost^(Martinetal.
1993;p.
8695).
Webegantonoticethatendosymbioticgenetransferandendosym-bioticgenereplacementwereprobablymorewidespreadthananyonehadsuspected;itwasanexcitingtime.
Notuntilmanyyearslater,withtheavailabilityofcompletegenomes,didpeopleseethatthevastmajorityofeukaryoticgenesstemfrombacteria,notfromarchaea(Esseretal.
2004;Pisanietal.
2007;CottonandMcInerney2010;Kuetal.
2015).
Today,weknowthateukaryotesarenotjustgrownuparchaea,theyaretheproductofendosymbiosis(Zimorskietal.
2014;Martinetal.
2015),butforsomereason,itisstillverypopulartosaythateukaryotesarosefromarchaeaortowriteabouttheBarchaealoriginofeukaryotes^(Williamsetal.
2013;Koonin2015),eventhoughthestatementis,ifwelookatthedataopenly,flatlywrongbecauseeukaryotespossessthreetimesmorebacterial-derivedgenesthattheypossessarchaealderivedgenes(Esseretal.
2004;Pisanietal.
2007;CottonandMcInerney2010;Kuetal.
2015).
Whatthedatasay(Zaremba-Niedzwiedzkaetal.
2017)isthatthehostfortheoriginofmitochondriahasanarchaealorigin(McInerney1826W.
F.
Martin,R.
Cerffetal.
2014).
AdemocraticgenomewithonegeneonevotewouldelecttheBbacterialoriginofeukaryotes.
^GAPDHwasjustthetipoftheiceberg.
TheevolutionofproteincompartmentationduringeukaryoteevolutionThefindingthatendosymbiontsmayhavedonatedmanygenestothenucleuswithoutorganellarreimportofthepro-tein(Martinetal.
1993)sharpenedourawarenesstoanissuethateventuallybecamequitepressing.
We(andeveryoneelse)hadbeenassumingthattheproductsofendosymbiont-derivedgeneswerespecifictotheorganellefromwhichthegenewasacquired;afterall,thiswaswhattheproductspecificitycorollarytoendosymbiotictheory,whichwehadbeentestingforadecadesaid.
CytosolicGAPDHwasanexception,andaplastid-targetedversionofcytosolicGAPDHwasalsoanexception(Meyer-Gauenetal.
1994;Petersenetal.
2003).
Whenwelookedaroundmoregenerally,almostalloftheenzymesofcentralcarbo-hydratemetabolisminplantswereanexception,anditwasquicklybecomingapparentthateukaryoteshadabacterialglycolyticpathway(Martin1996),whichwasalsotrueofeukaryotessuchasGiardiaorEntamoebathatlackedtypi-calmitochondria(Henzeetal.
1995),asshowninFig.
3.
Allthatprovidedcausetothinkabouttheprocessofendo-symbioticgenetransferasaprocessinrealtime.
Whenthebacterialancestorsofchloroplastsandmitochondriabecameendosymbionts,theydidnotbringalongtheirownpre-existingproteinimportmachinery.
Rather,boththeTiMandToMproteinimportmachineryofmitochondriaandtheTiCandToCproteinimportmachineryofplastidsareeukaryoticinventions,albeitconsistingofsomeprokaryotic-derivedba-siccomponents(SollandSchleiff2004),arosewithintheeukaryoticlineage,eachfollowingtheoriginoftherespectiveorganelle.
Theconsequenceofthatinsightisthatattheonsetofmitochondrialendosymbiosis,whenthefirstgeneswerebeingtransferredtothechromosomesofthehost,therewasnoplaceforthetranslatedproductsofendosymbiont-derivedgenestogoexcepttothecytosol(ortothehost'splasmamembrane).
Thatinturnmeansthatitshouldnotbeallthatsurprisingtoseeapredominanceofbacterialgenesineukary-otes.
Wewereassuming20yearsagothatwhichtoday'sdatasay(Zaremba-Niedzwiedzkaetal.
2017):Thehostthatac-quiredthemitochondrionwasanarchaeon,aprokaryote.
Genetransfersduringendosymbiosistransformedthearchaealhostfromwithinbecauseifthefirsttransferdidnotwork(thatis,ifitjustgaverisetoapseudogene),thenmaybethenexttransferwouldbesuccessful,ormaybethenextafterthat.
Attheoriginofeukaryotes,themitochondrialendosymbi-ont'sgenomewasacontinuousandpersistentsourceofnewgenesforthehost.
Attheonsetofthatsymbiosis,however,theproductsofthefirsttransferredgenesbecametargetedtothehost'scytosol.
OnlyafterthesymbioticconsortiumhadevolvedtheTiMandToMcomplexesinthetwomitochondri-almembranescouldtheproductsoftransferredgenesbereimported.
TheevolutionoftheTiMandToMcomplexesfosteredtheevolutionofN-terminaltransitpeptidesthatwoulddirectgeneproductsfaithfullytomitochondria.
GAPDHshedlightonthatprocess,too,andintronswereonceagaininvolved.
WalterGilbert'steamfoundacaseinwhichtheNAD-bindingdomainofcytosolicGAPDHhadbeenappendedtotheN-terminusofplantcytochromecviaexonshufflingtogenerateafunctionaltransitpeptide(Longetal.
1996).
Thus,whilechloroplastGAPDHpossessedtransitpep-tides,cytosolicGAPDHinplantsprovidedgeneticstartingmaterialforthegenerationoftransitpeptides,yetnotdenovo,ratherbyexonshuffling.
Thatwasallextremelyhelpfulforunderstandingthesitua-tion,butsomeoldimportantquestionsremainedandnewonesarose.
Amongtheoldimportantquestionswastheissueofwhy,withallthattransfergoingon,anygenesremainedinorganellesatall,whichJohnAllen'sCORRhypothesisexplainsbetterthananyothercompetingtheory,ithastodowiththeneedtomaintainredoxbalanceinbioenergeticmembranes,whichrequiresthatcomponentsoftheelectrontransportchainbeencodedintheorganelle(Allen2015).
Amongthenewquestionstounfoldfromthoseinsightswastheissueofwhyeukaryotes,whicheveryonethoughtdescendfromarchaea(sayribosomes),shouldhaveabacterialglycolyticpathwayinthefirstplaceinsteadoftheiroriginalarchaealglycolyticpathway.
Keepingthebiggerpictureinfocus—andremainingcriticalofthetheoreticalfoundationsuponwhichthewholeendosymbiosisendeavorwasresting—perhapstheanswerwasthatsomethingelseevenmorefunda-mentalwaswrongwithendosymbiotictheory.
WhatwaswrongAslongasglycolysishadbeenknownandforaslongEFGaEFTuVATPase16SrRNARNApolymeraseGAPDHLateraleubacterialtransferandsubstitutionofnucleargapCbEubacteriaEukaryotesArchaebacteriaEubacteriaEukaryotesArchaebacteriaEubacteriaEukaryotesArchaebacteriaFig.
2Domainrelationships.
aSchematicrepresentationofdomainrelationshipsforseveralmarkersusedatthetimethatshowedasistergrouprelationshipbetweeneukaryotesandarchaeacontrastedtothedomainrelationshipsdepictedbysequencesforglycolyticGAPDHsequences.
bInterpretationofthedifferencebetweendomainrelationshipsdepictedbythedatasets.
RedrawnafterMartinetal.
(1993)Physiology,phylogeny,earlyevolution,andGAPDH1827asanyoneeverthoughtthatmitochondriawereendosymbi-onts,everyonehadalwaysassumedthatthehostfortheoriginofthemitochondrionwasaheterotroph.
WhatifthehostwasanautotrophAnautotrophwouldactuallyrequireanewgly-colyticpathwayinordertosupplyitsmitochondriawithcar-bonsubstratesbecauseinautotrophs,carbonfluxisspecial-izedandoptimizedtorunfrompyruvatetoglucose.
Inordertofeedamitochondrioninthecytosolofanautotroph,atypicalbacterialEmbden-Meyerhofcataboliccarbonfluxwasneededasitoccursinalleukaryotes.
Stumpf's(1950)inferenceaboutglycolysisinplantsbeingBsimilartothatinyeastandanimaltissues^appliedtoalleukaryotesingeneral.
Glycolysiswaspresentintheeukaryoticcommonancestor,butwasitpresentinthehostAtthesametime,thereweretheanaerobicformsofmitochondriaoutthereforwhichendosymbiotictheoryalsodidnotaccount,hydrogenosomes(LindmarkandMüller1973;Müller1988).
Ifthehostwasanautotrophicarchaeon,thenitwouldbedependentuponH2,likemostarchaealautotrophsare(Fuchs2011).
IfitweredependentuponH2producedbythecommonancestorofmitochondriaandhydrogenosomes,thatwouldprovideaselectivepressuretoassociatetheendosymbionttoitshost(MartinandMüller1998).
Finally,wehadaversionofendosymbiotictheorywherethehostactuallyneededitsen-dosymbiont,butaverydifferentversionthanhadeverbeenoutthere,oneinwhichbygenetransfer,thesymbiont'sbac-terialglycolyticpathwayendedupinthecytosol,feedingthemitochondrion,andconvertingtheautotrophichostcellcom-partmentintoaheterotrophiccellcompartment(MartinandMüller1998).
Notonlywouldabacterialglycolyticpathwayineukaryotesmakesense,butsowouldthewidespreaddis-tributionofanaerobicpathwaysinmitochondria(Mülleretal.
2012).
Intermsofphysiology,andintermsofvisiblesyntrophicmicrobialinteractions(FenchelandFinlay1995),anautotrophichostfortheoriginofmitochondriaworksmuchbetterthanthephagocytosinghostthateveryonehadalwaysassumed.
Whilemicrobiologiststendedtoliketheidea(Madiganetal.
2012),inevolutionarycircles,thepriceforaphysiolog-icallybasedunderstandingoftheprokaryote-to-eukaryotetransitionwassteep.
Itmeantthat(i)allofMargulis'sversionsofendosymbiotictheory(Margulisetal.
2006)werewrong,thatdeDuve'sinterpretationsofhydrogenosomeswerewrong(deDuve2007),thatWoese'sthreedomaintree(Woeseetal.
1990)waswrong,andthatDoolittle's(1998)viewofunre-strictedandcontinuousinterdomainlateralgenetransferwasalsowrong.
Thatwasalotofauthoritybeingwrongallatonce.
However,theydidnotagreeonanythingamongthem-selveseither,andourintentwastogetthescienceright,nottoproveparticularscientistsright.
Lookingatthematteropenly,everyonewastryingtoaccountfortheirownfavoriteexplananda(flagella,peroxisomes,onebranchinonetree,andmanybranchesinmanytrees).
Weweretryingtoaccountforphysiology,thewaythatcellsstayaliveintheirenviron-ment,onthesimplebutrobustpremisethatchloroplastsandmitochondriawereoncefree-livingbacteria.
Theobjectofourundertakingwastoobtainsomeinsightsintocellevolutionfromthestartingpointofanimportantenzymecentraltocar-bonandenergymetabolism.
Naturedoesnotdivulgesecretsaboutearlyevolutionlightly,regardlessofwhetheronesetstheleveronphysiology,orelsewhere.
SecondaryendosymbiosisamongthealgaewithcomplexplastidsWelearnedfromGAPDHinteraliathatwhenitcomestoendosymbioticgenetransfer,thereisnohomingdevicethatdirectstheproductoftransferredgenesbacktotheorganellefromwhichthegenestems,ratheritisuptonaturalvariationandnaturalselectiontodeterminewherethegeneproductwillendup.
Therewasonceadebateaboutproteintargetinginiti-atedbydeDuve,whoarguedthatperoxisomeswereonceendosymbiontslikemitochondria,becausehowelse(sohisargument)couldonegetanentirepathwayintoanewcom-partmentFollowingdeDuve'sreasoningforthesymbioticoriginofperoxisomes(deDuve1969),MichelsandOpperdoes(1991)explainedwhyspecifictargetingofanen-tirepathwaytoaneworganelledoesindeedseemveryunlike-lyatfirstsight,itisbecauseofthis:Geneduplicationandmutationcanreadilyleadtothesignalneededtodirectoneenzymetoanewcompartment,butbyitself,thenewenzymeisuseless,forlackofsubstrate,forlackofdownstreamprod-uctconversion,orboth;hence,thenewtargetingvariantwillbeaburdenforthecellandwillbelost.
However,ifweconsiderthepossibilitythatproteintargetingwithineukaryot-iccellsmightnotalwaysbe100%specific,withminorZeaChondrusGallusDrosophilaEntamoebaGiardiaChlamydomonasSaccharomycesChlorophytaRhodophytaMetazoaMetazoaRhizopoda(nomt)Metamonada(nomt)ChlorophytaAscomycotaGapCGapCGapCGapCGapCGapCGapCGapCFig.
3LineagerelationshipsforGAPDHfromeukaryoteslackingtypicalmitochondria(abbreviatedasBnomt^).
RedrawnafterHenzeetal.
(1995)1828W.
F.
Martin,R.
Cerffamountsofmanyproteinsconstantlybeingimportedintothewrongorganelle,theneventheretargetingofentirepathwaysduringeukaryoteevolutiondoesnotseemsodifficult(Martin2010)becauseaverysmallamountofenzymecangoalongwayintermsofactivity,andaverysmallamountofanentirepathwayprovidesaunitoffunctionuponwhichselectioncanact.
Nowhereinnatureistheissueofcellcompartmentationmorecomplexthaninalgae.
TheconceptofsecondaryendosymbiosisgoesbacktoRobertE.
Lee(1977)andtoSarahGibbs(1978).
Itpositsthat—touseSitte'sterm—thecomplexplastidsofalgaede-scendfromtheengulfmentofaeukaryoticalgabyaeukary-otichost;suchplastidsaresurroundedbythreeorfourmem-branes.
GeneslikethatforchloroplastGAPDHthatstemfromthecyanobacterialancestorofplastidsshouldalsoreflectplas-tidphylogeny,buthavingbeentransferredtwiceduringevo-lution:onceduringprimaryendosymbiosisfromthecyanobacterialancestorofplastidstothenucleusofthehostandasecondtimefromthenucleusofthealgalendosymbionttothenucleusofitshost.
GAPDHgenesfromredalgae(Liaudetal.
1994)wereextremelyinsightfulinthisrespectastheytracedtheredalgalancestryofplastidsacrossthebreadthofthephotosyntheticeukaryoteshavingredcomplexplastids(Liaudetal.
1997;Liaudetal.
2000).
GAPDHgeneshelpeddiscriminatebetweencompetinghypothesisforthehighlyde-batednumberofsecondarysymbiosesinvolvingredalgalendosymbiontsthatoccurredduringeukaryoticevolution(Petersenetal.
2006b),andtheyhelpedtoclarifytheevolu-tionaryoriginoflandplantsthemselves(Petersenetal.
2006a).
RoundingoutthefamilyofplantGAPDH,therewasalsoanon-phosphorylatingformoftheenzyme,calledGAPN,thatturnedouttobeamemberofthealdehydedehy-drogenasefamily,whichdidhavehomologuesamongar-chaea,providingadditionalinsightsintotheearlydiversifica-tionofgenesinthelastuniversalcommonancestorofalllife(Habenichtetal.
1997).
ThereasonforbringingGAPDHoutofthecoldroomandintotherealmofmolecularbiologyinthefirstplacewastodiscriminatebetweencompetinghypotheses.
Amongtheal-gaewithcomplexplastids,thenumbersofpossibleendosym-bioticeventsandgenetransfersthatonecaninferfromGAPDHgenephylogeniescanbecomebewildering.
Intryingtoreconciletheobservedphyleticpatternswiththebiologyoftheorganisms—theexplanandumoftheintellectualexer-cise—itiseasytolosesightoftheforest(thebiology)forthegenetrees.
Genetreesarehelpfulwhenitcomestohy-pothesistesting,andtheyneitherhavethesameweight,asobservationsinnature,noraretheyeveninthesamecategory.
ThealgaepossessingGAPDHgenesandtheGAPDHgenesthemselvesarerealinthesensethattheyexistinnature.
Bycontrast,branchesingenetreesarenotreal.
Branchesintreescomefromcomputerprograms(orinthecaseofFig.
1,fromapencil,pen,sliderule,andLetraset).
Inordertobringthebranchingpatternsingenetreesintologicalagreementwithwhatwethinktruespeciesrelationshipsshouldbe,oneonlyneedstoaddfouringredients:phylogeneticerror,genelosses,endosymbioses,and(sinceour1986paperanyway)lateralgenetransfer.
Intheabsenceofaprioriknowledgeaboutthesubjectofinquiry,forwhosestudythetreewasconstructed,thereisnosimplewaytodecidehowmuchofwhichingredienttoadd.
Thatcallsforasoundphilosophicalfoundationforthescien-tificenquiryathand,somethinginwhichtheelderofusamplyreceivedformaltrainingduringadecadeofresearchinFreiburg.
ThephilosophyofscienceasitappliestopracticalempiricalinquiryissomethingaboutwhichweoftendiscussedinHannoverandBraunschweig,especiallywhenitwastimetothinkthingsthroughinfull,inthehopeofextractingbelastbare[Ger:sturdyinthesenseofbeingabletosupportsomething]insightsintoearlyevolutionfrompro-teinsequencedata.
Withoutgoingintojustification,detailordigression,twosetsofguidelinesalwaysseemedhelpful.
OneistheprincipleofOccam'srazorassplendidlydefinedinWebster's9thCollegiateDictionary:Bascientificandphil-osophicrulethatentitiesshouldnotbemultipliedunnecessar-ilywhichisinterpretedasrequiringthatthesimplestofcom-petingtheoriesbepreferredtothemorecomplexorthatex-planationsofunknownphenomenabesoughtfirstintermsofknownquantities.
^Thelast12wordsarethemostimportant.
TheysetOccam'srazormilesapartfrommereparsimony,andinpracticalterms,theytranslatetoausefuldoctrineforinfer-ence:Donotmakeupanythingunlessitisabsolutelyneces-saryandworkwithobservationsinnature.
AnothersetofreasoningguidelinescanbefoundinalittlenotedpaperbyLeeaboutthenumberandnatureofplastidsymbioses:BAnyevolutionaryschemeshouldadheretothefollowingthreeprinciples.
(1)Amonophyleticoriginofanyorganism,chemicalcompoundorcytoplasmicstructurehasthegreateststatisticalprobabilityofbeingcorrect.
(2)Thelossofanon-essentialstructurecanrequirejustthemutationofasinglegenebuttheacquisitionofastructuregenerallyrequiresmanymutationsandaconsiderableamountoftime.
(3)Mostorganismsinevolutionarysequenceswouldhavebeenlost,yetinpostulatingphylogeneticeventstheplausibilityofthetheorycanbeenhancedbytheexistenceoforganismssimilartothoseintheproposedscheme.
^(Lee1972,p.
44).
Manyoftoday'spapersaboutlateralgenetransferamongeukaryotescouldbenefitfromLee'srules,no.
2inparticular.
Thioesters,energy,andchemicalrelictsfromlife'soriginGAPDHcatalyzesawell-studiedchemicalreaction;themechanismofwhichisofinterest(SegalandBoyer1953,Bieseckeretal.
1977).
Intheoxidativedirection,thealdehydecarboninD-glyceraldehyde-3-phosphateisattackedbythethiolofCys149(BacillusstearothermophilusresiduePhysiology,phylogeny,earlyevolution,andGAPDH1829numbering)formingacovalentlyenzyme-boundhemithioacetal.
HydrideremovalbyNAD+oxidizesthesub-strate'sthiol-bondedcarbonatomtogenerateathioester,whichharborsanenergy-richbond(Lipmann1941)withafreeenergyofhydrolysisof32kJmol1(BuckelandEggerer1965).
Thesubstrateishowevernotremovedfromtheenzymebyhydrolysis,whichwouldreleasetheenergyasheat,ratherphosphorolysisreleases1,3-bisphospho-D-glycerate.
Theacylphosphatebondof1,3-bisphospho-D-glycerateconservestheenergy;ithasafreeenergyofhydro-lysisontheorderof52kJmol1(Meyerhof1951,Thaueretal.
1977)sufficienttoreadilyphosphorylateADPtoATP(freeenergyofhydrolysis31kJ·mol1,Thaueretal.
1977)viasubstratelevelphosphorylation(Fig.
4a).
Preciselythissequenceofreactionsfromthioestertoacylphosphate(amixedanhydride)totheβ-γphosphoanhydridebondinATPhaslongbeenthoughttobearelicfromtheverybeginningofmetabolism.
DeDuve(1991)suggestedthatsuchathioester–acylphosphate–ATPconversion,asitoccursinglycolysis,mightrepresenttheprimordialpathwayofenergyconservation,buthewaslookingattheproblemofearlyevo-lutionfromthestandpointofsugaroxidations.
Theproblemwithhisproposalisnotthebasicchemistry.
Rather,theprob-lemisthesurroundingenvironmentsupportingsuchareactionsequence.
Likegenerationsofscientistsbeforehim(Haldane1929;Wald1964),deDuvewasassumingthatlifestartedoffwithfermentations.
Thatseemslikeaharmlessassumption,butifwethinkitthroughinfullonthebasisofwhatweknowtodayaboutmicrobialphysiology,itturnsouttobealtogetheruntenable(Schnheitetal.
2016).
Forprimordialenergycon-servationintheoxidativedirectionoftheGAPDHreaction,themainproblemsaretwofold.
First,ontheearlyEarth,thereisnoevidencetosuggesttheexistenceofanyspontaneouslysynthesizedreservoirofanyparticularsugarisomer,noristherereasontoassumethatsuchamountainofsugarevenexisted.
ThisisthesameproblemencounteredwhentryingtoharnessenergyinsomemannerfromorganicsdeliveredtoEarthfromspace:Theorganiccompoundsincarbonaceouschondritesarenotonlyextremelyheterogeneousinstructure(Sephton2002)sothatnospecificisomercouldbecomethefocusofbiologicalenergyconservation,buttheyarealsounfermentablesubstrates(Schnheitetal.
2016),meaningthattheonlywaytoharnessenergyfromthemisthroughoxida-tion,whichrequiresanoxidant,anelectronacceptorlikeO2orFe3+orsimilar.
Theneedforanoxidantbringsustothesecondproblem.
Theinvocationofoxidants,suchasFe3+astheoriginalelec-tronacceptorsfortheoxidativesynthesisofthioestersviaareactionsimilartothatcatalyzedbyGAPDH(deDuve1991),hasanoftenoverlookednegativeconsequenceofgeneralsig-nificance.
InthepresenceofstrongoxidantslikeFe3+,carbonequilibriumliesonthesideofCO2ratherthanonthesideofreducedcarboncompounds,thebuildingblocksoflife(Sousaetal.
2013,AmendandMcCollom2009).
Fe3+isastrongoxidant.
Themidpointpotential,E0',oftheFe3+/Fe2+coupleis+772mV,thatoftheO2/H2Ocoupleis+818mV(Thaueretal.
1977).
Attheonsetofbiochemicalenergyharnessing(orattheoriginoflife,asoneprefers),thefirstandforemosthurdleisthegenerationandaccumulationofreducedorganiccompoundscomprisingthesubstanceofcells,nottheircon-versiontoCO2.
Neitherdisproportionations(fermentations)noroxidationofreducedorganiccompoundsofferatenablepathtogetstartedatbiochemicalorigins.
Whennavigatingthewatersofearlyevolution,physiologyisagoodcompass.
Seenasachemicalprocess,theenergymetaboliccomponentofphysiologyentailsfocusingtheflowofa(modestly)specificspectrumofsubstratesthoughincreas-inglynarrowchannelstowardsveryspecificexergonicreac-tionswhereenergyisconserved.
OntheearlyEarth,themainformofcarbonwasCO2(Sleepetal.
2011).
WecansaythatbecausetheearlyEarthwentthroughaphasewheretheplanetwasmoltenrock,whichishotterthan1000°C.
CarbonincontactwithaglobaloceanofmagmaexistsasCO2,notasglucoseoranythingsimilar.
WhentheEarthcooled,CO2remainedasthemaincarbonspecies.
Thatisgoodinaphys-iologicalsensebecauseitprovidesaveryspecificsubstrateforcorephysiologicalreactions.
Thesurprisingaspectformanyobserversisthatunderstrictlyanaerobicconditions,whichexistedontheearlyEarth,biologicalsystemscanactuallyharnessenergyfromCO2,withoutlight,whenusingH2astheelectronsource:Acetogens(SchuchmannandMüller2014)and,undersomeconditions,methanogenssuchasMethanosarcinaacetovoransgrowingonCO(RotherandMetcalf2004)cou-pletheexergonicreductionofCO2withH2tothesynthesisofATP.
ThatexergonicprocessofATPsynthesisentailsthesamesequenceofsubstratelevelphosphorylationinvolvingthioester–acylphosphate–ATPconversionasdeDuvehadinmind.
However,neitherthesubstratelevelphosphorylationsthatoccursinacetogensandmethanogensnortheenzymesinvolvedhaveanythingtodowithglycolysis(Fig.
4b).
EnergyisreleasedinthereductivedirectionbecauseinthereactionofH2withCO2,thechemicalequilibriumliesonthesideofreducedcarboncompounds(ShockandBoyd2015).
TheH2requiredtodrivesuchreactionsinthedirectionofCO2reductionwaspresentontheearlyEarth,inabun-dance,suppliedthroughageochemicalprocesscalledserpentinization,whichhasbeengeneratingH2inhydrother-malsystemsinmillimolesperliteramountssincetherewaswateronEarth(Sleepetal.
2004),andstilldoessotoday(McCollomandSeewald2013).
Acylphosphatesmighthaveevenbeentheancestralenergycurrencyoflife,moreancientthanATP(Martin2012).
Indeed,theirsynthesisfrommethylgroupsandCOinanaerobicautotrophsentailsreactionscata-lyzedbytransitionmetalsonly(Sousaetal.
2013),pointingtoaveryancientrolefortransitionmetalsinbiochemical1830W.
F.
Martin,R.
Cerffevolution,somethingthatbiologistshavealwayssuspected(EckandDayhoff1966).
Insupportofthatview,reconstruc-tionsofearlyphysiologicalevolutionfrominvestigationofancientgenes(Weissetal.
2016)providegenome-basedevi-dencethatthecommonancestorofalllifeconservedenergyfromthetransitionmetal-dependentreductionofCO2withelectronsfromH2.
Thus,fromtheperspectiveofthermodynamics,naturalH2-producinggeochemicalprocessescouldhaveenablereactionsinvolvingthesynthesisofsmallmolecularweightcarboncompoundsfromCO2tobecoupledtotheexergonicsynthesisofhigh-energyphosphatebonds.
Areactionsequenceofthattypecouldhaveresidedattheoriginofphysiologicalenergyconservation.
Ifso,thioesterswouldhaveprecededacylphos-phatesinevolutionandacylphosphateswouldhaveprecededATPastheuniversalenergycurrency.
ThatwouldmakesensebecauseitmeansthatthesimplestformofATPsynthesis,substratelevelphosphorylation,wouldhaveprecededchemi-osmoticATPsynthesis,whichrequiresproteins.
Fromthatwouldfollowthattheearlyhistoryofbiologicalenergycon-servationisrecapitulatedduringeverysinglecatalyticcycleoftheGAPDHreactionintheoxidativedirection,wherebyinearlyenergyconservationthereductivesynthesisofthioestersfromH2andCO2andtheirconversiontoacylphosphateswouldhavebeenexergonic,withacouplingofenergymetab-olismtoautotrophy.
EnvoiEarlyevolutionwillalwaysholdadeepelementoffascination.
Afterall,itisalmostimpossibletoimaginehowanythingascomplicatedaslifecouldgetstartedfromanythingassimpleasCysSHCOHRaCysSCOHRHNAD+NADH+H+CysSCORHPO4=CysSCORHPOOO––OCoASCOCH3HPO4=COPOOO––ONiCCH3CH3+SSONiFeCoASHNiCOCH3SSNiFeCoASHNiCCH3SSONiFeNiSSNiFeCoCOb+electronselectronsCoFig.
4Energyconservationashigh-energyphosphatebonds.
aMechanismoftheGAPDHreactionintheoxidativedirection,redrawnafterSegalandBoyer(1953)andBieseckeretal.
(1977),R=CHOHCH2OPO32.
Theverticalarrowunderscorestheoxidativenatureofthereversiblereactionwhendrawnfromlefttoright.
Forsustainedfluxinthatdirectiontoallowsustainedsynthesisofhigh-energyphosphatebonds,suitableelectronacceptorsmustbeavailableintheenvironment.
Theseacceptorscouldnothavebeengeneratedfermentativelyduringmetabolismoforganicsfromspacebecauseor-ganicsfromspaceareunfermentablesubstrates(Schnheitetal.
2016).
Assumingthe(debateable)existenceofaprimordialsourceofsugars,oxidativeenergyconservationrequirestheexistenceofoxidantsaselec-tronacceptors;oxidantsarehoweverthermodynamicallyunfavorablefortheaccumulationofthebuildingblocksoflife(seetext).
Underphysio-logicalconditions,theacylphosphatebondintherightmostcompound,1,3-bisphosphoglycerate,hasafreeenergyofhydrolysisof52kJ/mol(Thaueretal.
1977).
bReductivesynthesisofacylphosphate(acetylphosphate)fromCOandamethylgroupasitoccursintheacetyl-CoApathwayofacetogensin(Fuchs2011;SchuchmannandMüller2014)andinsomemethanogenswhengrownonCO(RotherandMetcalf2004).
ThereactionsaredrawnfromdatacompiledinSvetlitchnaiaetal.
(2006)ANDinRagsdale(2009)particularlyFigs.
S5andS6,inFuchs(2011),andinSchuchmannandMüller(2014).
Theverticalarrowindicatesthattheexergonicnatureofthereactionsequence,henceitsabilityforsustainedsynthesisofacylphosphate,entailsthecontinuousreductionofCO2andhencerequirestheenvironmentalavailabilityofasuitablereductantsuchasH2,whichwasabundantontheearlyEarth(Sleepetal.
,2011)andisstillabundanttodayinhydrothermalvents(McCollomandSeewald2013).
ElectronsfromtheH2/H+couple,whichhasastandardmidpointpotentialof414kJ/molatpH7(Thaueretal.
1977),areusedbyhydrogenotrophicacetogensandmethanogenstosynthesizeCOandthemethylgroupfromCO2(Fuchs2011,SchuchmannandMüller2014).
Underphysiologicalconditions,theacylphosphatebondintheacetylphosphatehasafreeenergyofhydrolysisof45kJ/mol(Thaueretal.
1977).
Inmicrobialinmetabolism,both1,3-bisphosphoglycerateandacetylphosphatetypicallyphosphorylateADPtoATP,whichhasfreeenergyofhydrolysisof32kJ/mol(Thaueretal.
1977),viasubstratelevelphosphorylation.
Somereadersmightobjecttotheuseofthetermsenergy-richbondorhigh-energybond,butthetermsareveryusefulandthelabilityofthebondsinquestionisfoundedinthecircumstancethatthevalenceelectronsofPandSareinthethirdshell,whichcanacceptfurtherelectronsinthedorbital,andinthecircumstancethatcorrespondingbondshavesubstantialbondlengths,offeringampleopportunityforat-tackbymoleculessuchaswaterthatpossessloneelectronpairs(Wald1964)Physiology,phylogeny,earlyevolution,andGAPDH1831rocks,water,andCO2ontheearlyEarth.
Peoplelookatearlyevolutionfrommanydifferentangles,fromchemicaltracesforearlylifetoreplication,togenetics,toself-organization,andfinallytotheoriginsoffree-livingprokaryotesthatalmost2billionyearsagogaverisetoeukaryotes—ourancestors—andtruebiologicalcomplexityviaendosymbiosis.
Whatmanyob-serversfailtonoteisthatlifeisachemicalreaction,anenergy-releasingchemicalreaction.
Thesubstanceoflife,andweour-selves,areultimatelyjustsideproductsofthatreaction,whosemeaningistokeepthereactiongoingandwhoseprerogativeistopondertheoriginofitall.
AcknowledgementsWorkinthelaboratoryofRCwasfundedbytheDFG,andworkinthelaboratoryofWFMisfundedbytheERC.
WethankKlausV.
Kowallikformanydiscussionsandcriticalcommentsonthetext,andwethankVerenaZimorskiforthehelpinpreparingthemanuscript.
OpenAccessThisarticleisdistributedunderthetermsoftheCreativeCommonsAttribution4.
0InternationalLicense(http://creativecommons.
org/licenses/by/4.
0/),whichpermitsunrestricteduse,distribution,andreproductioninanymedium,providedyougiveappro-priatecredittotheoriginalauthor(s)andthesource,providealinktotheCreativeCommonslicense,andindicateifchangesweremade.
ReferencesAllenJF(2015)Whychloroplastsandmitochondriaretaintheirowngenomesandgeneticsystems:colocationforredoxregulationofgeneexpression.
ProcNatlAcadSciUSA112:10231–10238.
doi:10.
1073/pnas.
1500012112AmendJP,McCollomTM(2009)EnergeticsofbiomoleculesynthesisonearlyEarth.
In:ZaikowskiL,FriedrichJM,RussellSeidelS(eds)ChemicalevolutionII:fromtheoriginsoflifetomodernsociety.
AmericanChemicalSociety,Washington,DC,pp63–94ArchibaldJM(2014)Oneplusoneequalsone:symbiosisandtheevolu-tionofcomplexlife.
OxfordUniversityPress,Oxford,UKBieseckerG,HarrisJI,ThierryJC,WalkerJE,WonacottAJ(1977)SequenceandstructureofD-glyceraldehyde-3-phosphatedehydro-genasefromBacillusstearothermophilus.
Nature266:328–333BrinkmannH,MartinezP,QuigleyF,MartinW,CerffR(1987)Endosymbioticoriginandcodonbiasofthenucleargeneforchlo-roplastglyceraldehyde-3-phosphatedehydrogenasefrommaize.
JMolEvol26:320–328.
doi:10.
1007/BF02101150BrinkmannH,CerffR,SalomonM,SollJ(1989)CloningandsequenceanalysisofcDNAsencodingthecytosolicprecursorsofsubunitsGapAandGapBofchloroplastglyceraldehyde-3-phosphatedehy-drogenasesfrompeaandspinach.
PlantMolBiol13:81–94BuckelW,EggererH(1965)Ontheopticaldeterminationofcitratesyn-thaseandacetyl-coenzymea.
BiochemZ343:29–43CerffR(1974)Inhibitor-dependentreciprocalchangesintheactivitiesofglyceraldehyde3-phosphatedehydrogenasesinSinapisalbacotyle-dons.
ZPflanzenphysiol73:109–118CerffR(1978a)Glyceraldehyde-3-phosphatedehydrogenase(NADP)fromSinapisalbaL.
:reversibleassociationofenzymewithapro-teinfactorascontrolledbypyridinenucleotidesinvitro.
PlantPhysiol61:369–372.
doi:10.
1104/pp.
61.
3.
369CerffR(1978b)Glyceraldehyde-3-phosphatedehydrogenase(NADP)fromSinapisalbaL.
:steady-statekinetics.
Phytochemistry17:2061–2067.
doi:10.
1016/S0031-9422(00)89281-XCerffR(1979)Quaternarystructureofhigherplantglyceraldehyde-3-phosphatedehydrogenases.
EurJBiochem94:243–247.
doi:10.
1111/j.
1432-1033.
1979.
tb12891.
xCerffR(1982a)SeparationandpurificationofNAD-andNADP-linkedglyceraldehyde-3-phosphatedehydrogenasesfromhigherplants.
In:EdelmanM,HallickRB,ChuaN-H(eds)Methodsinchloroplastmolecularbiology.
ElsevierBiomedicalPress,Amsterdam,pp683–694CerffR(1982b)Evolutionarydivergenceofchloroplastandcytosolicglyceraldehyde-3-phosphatedehydrogenasesfromangiosperms.
EurJBiochem126:513–515.
doi:10.
1111/j.
1432-1033.
1982.
tb06810.
xCerffR,ChambersSE(1979)Subunitstructureofhigherplantglyceraldehyde-3-phosphatedehydrogenase(EC1.
2.
1.
12andEC1.
2.
1.
12).
JBiolChem254:6094–6098CerffR,KloppstechK(1982)Structuraldiversityanddifferentiallightcontrolofmessenger-RNAscodingforangiospermglyceraldehyde-3-phosphatedehydrogenases.
ProcNatlAcadSciUSA79:7624–7628.
doi:10.
1073/pnas.
79.
24.
7624CerffR,MartinW,BrinkmannH(1994)Originofintrons-earlyorlate.
Nature369:527–528CoriGT,SleinMW,CoriCF(1948)CrystallineD-glyceraldehyde-3-phosphatedehydrogenasefromrabbitmuscle.
JBiolChem173:605–618CottonJA,McInerneyJO(2010)Eukaryoticgenesofarchaebacterialoriginaremoreimportantthanthemorenumerouseubacterialgenes,irrespectiveoffunction.
ProcNatlAcadSciUSA107:17252–17255.
doi:10.
1073/pnas.
1000265107DayhoffMO(1965)Atlasofproteinsequenceandstructure.
NationalBiomedicalResearchFoundation,MarylanddeDuveC(1969)Evolutionoftheperoxisome.
AnnNYAcadSci168:369–381.
doi:10.
1111/j.
1749-6632.
1969.
tb43124.
xdeDuveC(1991)Blueprintforacell:thenatureandoriginoflife.
NeilPattersonPublishers,Burlington,NorthCarolinadeDuveC(2007)Theoriginofeukaryotes:areappraisal.
NatRevGenet8:395–403.
doi:10.
1038/nrg2071DoolittleWF(1998)Youarewhatyoueat:agenetransferratchetcouldaccountforbacterialgenesineukaryoticnucleargenomes.
TrendsGenet14:307–311EckRV,DayhoffMO(1966)Evolutionofthestructureofferredoxinbasedonlivingrelicsofprimitiveaminoacidsequences.
Science152:363–366EsserC,AhmadinejadN,WiegandC,RotteC,SebastianiF,Gelius-DietrichG,HenzeK,KretschmannE,RichlyE,LeisterD,BryantD,SteelMA,LockhartPJ,PennyD,MartinW(2004)Agenomephylogenyformitochondriaamongalpha-proteobacteriaandapre-dominantlyeubacterialancestryofyeastnucleargenes.
MolBiolEvol21:1643–1660.
doi:10.
1093/molbev/msh160FabryS,HenselR(1988)Primarystructureofglyceraldehyde-3-phosphatedehydrogenasededucedfromthenucleotidesequenceofthethermophilicarchaebacteriumMethanothermusfervidus.
Gene64:189–197.
doi:10.
1016/0378-1119(88)90334-4FenchelT,FinlayBJ(1995)Ecologyandevolutioninanoxicworlds.
OxfordUniversityPress,OxfordFitchWM,MargoliashE(1967)Constructionofphylogenetictrees.
Science155:279–284FuchsG(2011)Alternativepathwaysofcarbondioxidefixation:insightsintotheearlyevolutionoflifeAnnuRevMicrobiol65:631–658.
doi:10.
1146/annurev-micro-090110-102801GibbsM(1952)Triosephosphatedehydrogenaseandglucose-6-phosphatedehydrogenaseinthepeaplant.
Nature170:164–165.
doi:10.
1038/170164a0GibbsSP(1978)ThechloroplastsofEuglenamayhaveevolvedfromsymbioticgreenalgae.
CanJBot56:2883–2889GilbertW(1978)WhygenesinpiecesNature271:501.
doi:10.
1038/271501a01832W.
F.
Martin,R.
CerffGilbertW(1986)Originoflife:theRNAworld.
Nature319:618.
doi:10.
1038/319618a0GilbertW(1987)Theexontheoryofgenes.
ColdSpringHarbSympQuantBiol52:901–905GouldSB,WallerRF,McFaddenGI(2008)Plastidevolution.
AnnuRevPlantBiol59:491–517.
doi:10.
1146/annurev.
arplant.
59.
032607.
092915HabenichtA,QuesadaA,CerffR(1997)Sequenceofthenon-phosphorylatingglyceraldehyde-3-phosphatedehydrogenasefromNicotianaplumbaginifoliaandphylogeneticoriginofthegenefam-ily.
Gene198:237–243.
doi:10.
1016/S0378-1119(97)00320-XHaldaneJBS(1929)Theoriginoflife.
RationalistAnnual1929:3–10HenzeK,BadrA,WetternM,CerffR,MartinW(1995)AnucleargeneofeubacterialorigininEuglenagracilisreflectscrypticendosymbi-osesduringprotistevolution.
ProcNatlAcadSciUSA92:9122–9126.
doi:10.
1073/pnas.
92.
20.
9122HerrmannRG,BohnertHJ,KowallikKV,SchmittJM(1975)Size,con-formationandpurityofchloroplastDNAofhigherplants.
BiochimBiophysActa378:305–317.
doi:10.
1016/0005-2787(75)90118-5KersanachR,BrinkmannH,LiaudMF,ZhangDX,MartinW,CerffR(1994)Fiveidenticalintronpositionsinancientduplicatedgenesofeubacterialorigin.
Nature367:387–389.
doi:10.
1038/367387a0KooninEV(2015)Originofeukaryotesfromwithinarchaea,archaealeukaryome,andburstsofgenegain:eukaryogenesisjustmadeeas-ierPhilTransRSocLondBBiolSci370:20140333.
doi:10.
1098/rstb.
2014.
0333KowallikKV,HaberkornG(1971)TheDNA-structuresofthechloro-plastofProrocentrummicans(Dinophyceae).
ArchMicrobiol80:252–261KuC,Nelson-SathiS,RoettgerM,SousaFL,LockhartPJ,BryantD,Hazkani-CovoE,McInerneyJO,LandanG,MartinWF(2015)Endosymbioticoriginanddifferentiallossofeukaryoticgenes.
Nature524:427–432.
doi:10.
1038/nature14963LeeRE(1972)Originofplastidsandthephylogenyofalgae.
Nature237:44–46LeeRE(1977)Evolutionofalgalflagellateswithchloroplastendoplas-micreticulumfromtheciliates.
SAfrJSci73:179–182LiaudMF,ZhangDX,CerffR(1990)Differentialintronlossandendo-symbiotictransferofchloroplastglyceraldehyde-3-phosphatedehy-drogenasegenestothenucleus.
ProcNatlAcadSciUSA87:8918–8922LiaudMF,ValentinC,MartinW,BougetFY,KloaregB,CerffR(1994)Theevolutionaryoriginofredalgaeasdeducedfromthenucleargenesencodingcytosolicandchloroplastglyceraldehyde-3-phosphatedehydrogenasesfromChondruscrispus.
JMolEvol38:319–327.
doi:10.
1007/BF00163149LiaudMF,BrandtU,ScherzingerM,CerffR(1997)Evolutionaryoriginofcryptomonadmicroalgae:twonovelchloroplast/cytosol-specificGAPDHgenesaspotentialmarkersofancestralendosymbiontandhostcellcomponents.
JMolEvol44:S28–S37.
doi:10.
1007/PL00000050LiaudMF,LichtléC,AptK,MartinW,CerffR(2000)Compartment-specificisoformsofTPIandGAPDHareimportedintodiatommitochondriaasafusionprotein:evidenceinfavorofamitochon-drialoriginoftheeukaryoticglycolyticpathway.
MolBiolEvol17:213–223LindmarkDG,MüllerM(1973)Hydrogenosome,acytoplasmicorgan-elleoftheanaerobicflagellateTritrichomonasfoetus,anditsroleinpyruvatemetabolism.
JBiolChem248:7724–7728LipmannF(1941)Metabolicgenerationandutilizationofphosphatebondenergy.
AdvEnzymol1:99–162LongM,DeSouzaS,RosenbergC,GilbertW(1996)Exonshufflingandtheoriginofthemitochondrialtargetingfunctioninplantcyto-chromec1precusor.
ProcNatlAcadSciUSA93:7727–7731MadiganMT,MartinkoJM,BenderKS,BuckleyDH,StahlDA(2012)Brockbiologyofmicroorganisms,13thedn.
BenjaminCummings,BostonMargulisL,ChapmanM,GuerreroR,HallJ(2006)Thelasteukaryoticcommonancestor(LECA):acquisitionofcytoskeletalmotilityfromaerotolerantspirochetesintheProterozoicEon.
ProcNatlAcadSciUSA103:13080–13085.
doi:10.
1073/pnas.
0604985103MartinW(1985)Klonierung,IdentifizierungundSequenzierungvoncDNAsfürNAD+-undNADP+-spezifischeGAPDHsausSinapisalba.
DiplomarbeitamFachbereichBiologiederTechnischenUniversittHanoverunterderAnleitungvonR.
CerffMartinWF(1996)IssomethingwrongwiththetreeoflifeBioEssays18:523–527.
doi:10.
1002/bies.
950180702MartinW(2010)Evolutionaryoriginsofmetaboliccompartmentaliza-tionineukaryotes.
PhilosTransRSocLondB365:847–855.
doi:10.
1098/rstb.
2009.
0252MartinWF(2012)Hydrogen,metals,bifurcatingelectrons,andprotongradients:theearlyevolutionofbiologicalenergyconservation.
FEBSLett9:485–493MartinW,CerffR(1986)Prokaryoticfeaturesofanucleus-encodedenzyme:cDNAsequencesforchloroplastandcytosolicglyceraldehyde-3-phosphatedehydrogenasesfrommustard(Sinapisalba).
EurJBiochem159:323–331.
doi:10.
1111/j.
1432-1033.
1986.
tb09871.
xMartinW,MüllerM(1998)Thehydrogenhypothesisforthefirsteukary-ote.
Nature392:37–41.
doi:10.
1038/32096MartinW,BrinkmannH,SavonaC,CerffR(1993)Evidenceforachi-mericnatureofnucleargenomes:eubacterialoriginofeukaryoticglyceraldehyde-3-phosphatedehydrogenasegenes.
ProcNatlAcadSciUSA90:8692–8696.
doi:10.
1073/pnas.
90.
18.
8692MartinW,GargS,ZimorskiV(2015)Endosymbiotictheoriesforeu-karyoteorigin.
PhilosTransRSocLondB370:20140330.
doi:10.
1098/rstb.
2014.
0330McCollomTM,SeewaldJS(2013)Serpentinites,hydrogen,andlife.
Elements9:129–134.
doi:10.
2113/gselements.
9.
2.
129McInerneyJO,O'ConnellM,PisaniD(2014)ThehybridnatureoftheEukaryotaandaconsilientviewoflifeonEarth.
NatRevMicrobiol12:449–455.
doi:10.
1038/nrmicro3271MelandriBA,PupilloP,Baccarini-MelandriA(1970)D-glyceraldehyde-3-phosphatedehydrogenaseinphotosyntheticcells.
I.
Thereversiblelight-inducedactivationinvivoofNADP-dependentenzymeanditsrelationshiptoNAD-dependentactivities.
BiochimBiophysActa220:178–189.
doi:10.
1016/0005-2744(70)90004-5MereschkowskyC(1905)berNaturundUrsprungderChromatophorenimPflanzenreiche.
BiolCentralbl25:593–604[Englishtranslationin:MartinW,KowallikK(1999)AnnotatedEnglishtranslationofMereschkowsky's1905paper.
EurJPhycol34:287–295]Meyer-GauenG,SchnarrenbergerC,CerffR,MartinW(1994)Molecularcharacterizationofanovel,nuclear-encoded,NAD+-de-pendentglyceraldehyde-3-phosphatedehydrogenaseinplastidsofthegymnospermPinussylvestrisL.
PlantMolBiol26:1155–1166.
doi:10.
1007/BF00040696MeyerhofO(1951)Phosphorusmetabolism.
AmSci39:682–687MichelsPAM,OpperdoesFR(1991)Theevolutionaryoriginofglycosomes.
ParasitolToday7:105–109.
doi:10.
1016/0169-4758(91)90167-MMüllerM(1988)Energymetabolismofprotozoawithoutmitochondria.
AnnuRevMicrobiol42:465–488.
doi:10.
1146/annurev.
mi.
42.
100188.
002341MüllerM,MentelM,vanHellemondJJ,HenzeK,WoehleC,GouldSB,YuRY,vanderGiezenM,TielensAGM,MartinWF(2012)Biochemistryandevolutionofanaerobicenergymetabolismineu-karyotes.
MicrobiolMolBiolRev76:444–495.
doi:10.
1128/MMBR.
05024-11Physiology,phylogeny,earlyevolution,andGAPDH1833NeedhamDM,PillaiRK(1937)Thecouplingofoxido-reductionsanddismutationswithesterificationofphosphateinmuscle.
BiochemJ31:1837–1851PetersenJ,BrinkmannH,CerffR(2003)Origin,evolution,andmetabol-icroleofanovelglycolyticGAPDHenzymerecruitedbylandplantplastids.
JMolEvol57:16–26.
doi:10.
1007/s00239-002-2441-yPetersenJ,TeichR,BeckerB,CerffR,BrinkmannH(2006a)TheGapA/Bgeneduplicationmarkstheoriginofstreptophyta(Charophytesandlandplants).
MolBiolEvol23:1109–1118.
doi:10.
1093/molbev/msj123PetersenJ,TeichR,BrinkmannH,CerffR(2006b)ABgreen^phosphoribulokinaseincomplexalgaewithredplastids:evidenceforasinglesecondaryendosymbiosisleadingtohaptophytes,cryptophytes,heterokonts,anddinoflagellates.
JMolEvol62:143–157.
doi:10.
1007/s00239-004-0305-3PisaniD,CottonJA,McInerneyJO(2007)Supertreesdisentanglethechimericaloriginofeukaryoticgenomes.
MolBiolEvol24:1752–1760.
doi:10.
1093/molbev/msm095QuigleyF,MartinW,CerffR(1988)Intronconservationacrosstheprokaryote-eukaryoteboundry:structureofthenucleargeneforchloroplastglyceraldehyde-3-phosphatedehydrogenasefrommaize.
ProcNatlAcadSciUSA85:2672–2676RagsdaleSW(2009)Nickel-basedenzymesystems.
JBiolChem284:18571–18575RotherM,MetcalfWW(2004)AnaerobicgrowthofMethanosarcinaacetivoransC2Aoncarbonmonoxide:anunusualwayoflifeforamethanogenicarchaeon.
ProcNatlAcadSciUSA101:16929–16934.
doi:10.
1073/pnas.
0407486101SaganL(1967)Ontheoriginofmitosingcells.
JTheoretBiol14:225–274.
doi:10.
1016/0022-5193(67)90079-3SchnheitP,BuckelW,MartinWF(2016)Ontheoriginofheterotrophy.
TrendsMicrobiol24:12–25.
doi:10.
1016/j.
tim.
2015.
10.
003SchuchmannK,MüllerV(2014)Autotrophyatthethermodynamiclimitoflife:amodelforenergyconservationinacetogenicbacteria.
NatRevMicrobiol12:809–821.
doi:10.
1038/nrmicro3365SchwartzRM,DayhoffMO(1978)Originsofprokaryotes,eukaryotes,mitochondria,andchloroplasts.
Science199:395–403.
doi:10.
1126/science.
202030SegalHL,BoyerPD(1953)TheroleofsulfhydrylgroupsintheactivityofD-glyceraldehyde-3-phosphatedehydrogenase.
JBiolChem204:265–281SephtonMA(2002)Organiccompoundsincarbonaceousmeteorites.
NatProdRep19:292–311.
doi:10.
1039/b103775gShockEL,BoydES(2015)Geomicrobiologyandmicrobialgeochemis-try:principlesofgeobiochemistry.
Elements11:389–394.
doi:10.
2113/gselements.
11.
6.
395SitteP(2007)EndosymbioseinderZell-undLebensevolution.
In:StorchV,WelschU,WinkM(eds)Evolutionsbiologie,2.
Auflage.
Springer,Berlin,Heidelberg,NewYork,pp227–238SleepNH,MeibomA,FridrikssonT,ColemanRG,BirdDK(2004)H2-richfluidsfromserpentinization:geochemicalandbioticimplica-tions.
ProcNatlAcadSciUSA101:12818–12823.
doi:10.
1073/pnas.
0405289101SleepNH,BirdDK,PopeEC(2011)Serpentiniteandthedawnoflife.
PhilosTransRSocLondSerBBiolSci366:2857–2869.
doi:10.
1098/rstb.
2011.
0129SollJ,SchleiffE(2004)Proteinimportintochloroplasts.
NatRevMolCellBiol5:198–208.
doi:10.
1038/nrm1333SousaFL,ThiergartT,LandanG,Nelson-SathiS,PereiraIAC,AllenJF,LaneN,MartinWF(2013)Earlybioenergeticevolution.
PhilosTransRSocLondB368:20130088.
doi:10.
1098/rstb.
2013.
0088StoebeB,MaierUG(2002)One,two,three:nature'stoolboxforbuildingplastids.
Protoplasma219:123–130.
doi:10.
1007/s007090200013StumpfPK(1950)Carbohydratemetabolisminhigherplants.
III.
Breakdownoffructosediphosphatebypeaextracts.
JBiolChem182:261–272SvetlitchnaiaT,SvetlitchnyiV,MeyerO,DobbekH(2006)Structuralinsightsintomethyltransferreactionsofacorrinoidiron-sulfurpro-teininvolvedinacetyl-CoAsynthesis.
ProcNatlAcadSciUSA103:14331–14336ThauerRK,JungermannK,DeckerK(1977)Energyconservationinchemotrophicanaerobicbacteria.
BacteriolRev41:100–180WaldG(1964)Theoriginsoflife.
ProcNatlAcadSciUSA52:595–611WallinIE(1925)Onthenatureofmitochondria.
IX.
Demonstrationofthebacterialnatureofmitochondria.
AmJAnat36:131–139.
doi:10.
1002/aja.
1000360106WarburgO,ChristianW(1939)IsolierungundKrystallisationdesProteinsdesoxydierendenGarungsferments.
BiochemZ303:40–68WedelN,SollJ(1998)EvolutionaryconservedlightregulationofCalvincycleactivitybyNADPH-mediatedreversiblephosphoribulokinase/CP12/glyceraldehyde-3-phosphatedehydro-genasecomplexdissociation.
ProcNatlAcadSciUSA95:9699–9704WedelN,SollJ,PaapB(1997)CP12providesanewmodeoflightregulationofCalvincycleactivityinhigherplants.
ProcNatlAcadSciUSA94:10479–10484WeedenNF(1981)Geneticandbiochemicalimplicationsoftheendo-symbioticoriginofthechloroplast.
JMolEvol17:133–139.
doi:10.
1007/BF01733906WeissMC,SousaFL,MrnjavacN,NeukirchenS,RttgerM,Nelson-SathiS,MartinWF.
Thephysiologyandhabitatofthelastuniversalcommonancestor.
NatureMicrobiology1:16116(2016)WilliamsTA,FosterPG,CoxCJ,EmbleyTM(2013)Anarchaealoriginofeukaryotessupportsonlytwoprimarydomainsoflife.
Nature504:231–236.
doi:10.
1038/nature12779WoeseCR,KandlerO,WheelisML(1990)Towardsanaturalsystemoforganisms:proposalforthedomainsarchaea,bacteria,andEucarya.
ProcNatlAcadSciUSA87:4576–4579.
doi:10.
1073/pnas.
87.
12.
4576Zaremba-NiedzwiedzkaK,CaceresEF,SawJH,BckstrmD,JuzokaiteL,VancaesterE,SeitzKW,AnantharamanK,StarnawskiP,KjeldsenKU,StottMB,NunouraT,BanfieldJF,SchrammA,BakerBJ,SpangA,EttemaTJG(2017)Asgardarchaeailluminatetheoriginofeukaryoticcellularcomplexity.
Nature541:353–358.
doi:10.
1038/nature21031ZimorskiV,KuC,MartinWF,GouldSB(2014)Endosymbiotictheoryfororganelleorigins.
CurrOpinMicrobiol22:38–48.
doi:10.
1016/j.
mib.
2014.
09.
008ZwicklP,FabryS,BogedianC,HaasA,HenselR(1990)Glyceraldehyde-3-phosphatedehydrogenasefromthehyperthermo-philicarchaebacteriumPyrococcuswoesei:characterisationoftheenzyme,cloningandsequencingofthegene,andexpressioninEscherichiacoli.
JBacteriol172:4329–43381834W.
F.
Martin,R.
Cerff
- glyk12818.com相关文档
- 4.k12818.com
- ghsunilk12818.com
- epidemiologicalk12818.com
- interferencek12818.com
- obtaink12818.com
- Aberk12818.com
VirMach(8元/月)KVM VPS,北美、欧洲
VirMach,成立于2014年的美国IDC商家,知名的低价便宜VPS销售商,支持支付宝、微信、PayPal等方式付款购买,主打美国、欧洲暑假中心产品,拥有包括洛杉矶、西雅图、圣何塞、凤凰城在内的11个数据中心可以选择,可以自由搭配1Gbps、2Gbps、10Gbps带宽端口,有Voxility DDoS高防IP可以选择(500Gbps以上的防御能力),并且支持在控制面板付费切换机房和更换IP(带...
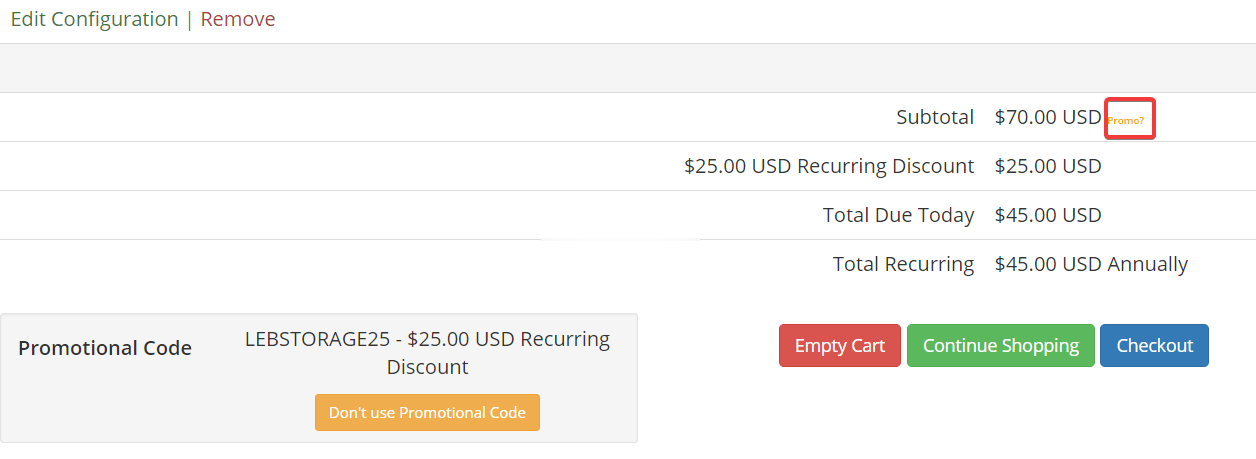
HostKvm($4.25/月)俄罗斯/香港高防VPS
HostKvm又上新了,这次上架了2个线路产品:俄罗斯和香港高防VPS,其中俄罗斯经测试电信CN2线路,而香港高防VPS提供30Gbps攻击防御。HostKvm是一家成立于2013年的国外主机服务商,主要提供基于KVM架构的VPS主机,可选数据中心包括日本、新加坡、韩国、美国、中国香港等多个地区机房,均为国内直连或优化线路,延迟较低,适合建站或者远程办公等。俄罗斯VPSCPU:1core内存:2G...
spinservers($89/月),圣何塞10Gbps带宽服务器,达拉斯10Gbps服务器
spinservers是Majestic Hosting Solutions LLC旗下站点,主要提供国外服务器租用和Hybrid Dedicated等产品的商家,数据中心包括美国达拉斯和圣何塞机房,机器一般10Gbps端口带宽,高配置硬件,支持使用PayPal、信用卡、支付宝或者微信等付款方式。目前,商家针对部分服务器提供优惠码,优惠后达拉斯机房服务器最低每月89美元起,圣何塞机房服务器最低每月...
k12818.com为你推荐
-
嘉兴商标注册怎么查商标注册日期地陷裂口造成地陷都有哪些原因?www.jjwxc.net在哪个网站看小说?百度指数词什么是百度指数bbs2.99nets.com这个"风情东南亚"网站有78kg.cn做网址又用bbs.风情东南亚.cn那么多此一举啊!kb123.netwww.zhmmjyw.net百度收录慢?partnersonline我家Internet Explorer为什么开不起来www.cn12365.orgwww.12365china.net是可靠的网站吗?还是骗子拿出来忽悠人的朴容熙这个人男的女的,哪国人。叫什么。鹤城勿扰齐齐哈尔,又叫鹤城吗?