基因关于TEL基因与白血病
关于TEL基因与白血病
目录
1. 1 TEL基因的结构特征
1. 2 TEL基因的功能
1. 3 TEL基因的表达产物
2. 1 TEL基因与急性淋巴细胞白血病
2. 2 TEL基因与急性粒细胞白血病
2. 3 TEL基因与慢性白血病
(5; 12) (q31;p13)染色体t(5; 12)易位见于嗜酸细胞增多型慢性粒单核细胞白血病CMML
3 问题与展望
正文
关键字基因
TEL基因又称ETV6基因是在急、慢性白血病骨髓异常增生综合征MD S和恶性淋巴瘤中均受累的所谓“杂合”基因多种染色体易位均可导致TEL基因重排而不同的TEL基因重排可通过不同的机制参与白血病的发生1 。 由于TEL基因的重要性其功能和导致白血病发病的机制受到广泛的关注。
1 TEL基因
1. 1 TEL基因的结构特征
TEL基因定位于12p13全长300 kb 由8个外显子构成新近又在3号外显子上游发现了另一个替代的1号外显子。该基因编码452个氨基酸是转录因子ETS家族的成员之一2 。 ETS家族都有一个ETS结构域该结构域能识别核心基元GGAA/T介导TEL基因和DNA的结合。 ETS家族的一些蛋白如ETS 1、 FLI 1等的异常在鼠和鸡白血病的发生中起重要的作用3 。 ETS结构域位于TEL基因的羧基端TEL的氨基端为一保守的HLH结构域又称B结构域或poi nted结构域 HLH结构域主要作为自身寡聚合域起作用也能促进TE LAML 1的同二聚体及其他HLH核蛋白的异二聚体的形成参与基因表达的调控。
1.2 TEL基因的功能
LOPEZ等4研究显示 TEL是一个序列特异的转录抑制因子其抑制活性依赖于由B结构域介导的寡聚化。 TEL的寡聚化结构域缺失其抑制活性就被破坏相反如果用一个无关的寡聚化结构域代替TEL的寡聚化结构域则可增强TEL的抑制活性说明TEL的抑制活性依赖于其自身寡聚化的能力。 TEL的转录是从端粒向着丝粒的方向其蛋白拥有一个70个氨基酸与DNA结合的E TS序列 以单聚体的形式结合至DNA调节细胞的分化和增殖。 TEL基因域对各系骨髓造血的建立是必不可少的并对胚胎造血也有一定的影响。剔除TEL基因的小鼠胚胎 因缺乏血管网络的发生及神经和间质细胞的死亡而死亡3 。TEL-/-小鼠胚胎缺乏建立骨髓三系造血的能力并反映出TEL-/-的干细胞在迁移植入骨髓时的能力缺陷。因此 TEL被认为是从胚肝造血到骨髓造血的开关基因。 TEL对造血的调控主要通过结合至自身或(和)FLI 1上两种方式5 。 FLI 1是ETS家族成员能反式激活巨核细胞的启动子而TEL能抑制这一效应但这种效应需要全长的TEL分子包括DNA结合结构域和HLH结合结构域。 由此推测一个突变或重组的TEL可能导致造血调控功能的紊乱而出现恶性表型。 TEL的表达不仅限于造血系统在人和鼠的所有成熟组织中均可检测到TEL的表达。
1.3 TEL基因的表达产物
TEL基因的表达产物为451个氨基酸组成的蛋白多肽属转录因子ETS癌基因家族成员 由羧基端的ETS DNA结合结构域和氨基端HLH结构域组成。 ETS结构域负责与DNA结合。 HLH结构域主要作为自身寡聚
合域起作用此外也可能介导TEL与其他各种不同转录因子间的蛋白蛋白的相互作用6
2 TEL基因与白血病
TEL基因是许多染色体异常累及的靶基因到目前报道的TEL基因相关的染色体易位超过30种7~9 。不同的TEL基因重排可通过不同的机制参与白血病的发生。
2. 1 TEL基因与急性淋巴细胞白血病
急性淋巴细胞白血病ALL在儿童白血病中约占75% 10 。 TEL基因在儿童白血病发病机制中的作用是从发现TELAML 1基因开始引起重视的。
TELAML1基因与ALL TELAML1 t(12;21)是儿童ALL中最常见的染色体结构异常 由12号染色体上的TEL基因与21号染色体上的AML 1基因融合而成在TELAML 1融合基因形成的同时往往伴随另一个等位基因的缺失易位和缺失使TEL/AML 1致白血病的机制复杂化。 TEL和AML1的融合使AML1编码的Cbra失去与DNA结合能力或不能正常转录而发挥激活靶基因的作用即AML 1从转录激活因子转变成转录抑制因子11 。此外 12p缺失在引起TEL基因缺失的同时还累及其下游的CDKNIB基因 由于该基因能编码周期素依赖激酶抑制蛋白P27具有阻止细胞从G1期进入S期抑制细胞增殖的作用故其受累加速了白血病细胞的过度生长12 。 SHURTLE等13于1995年发现近30%的前B急性淋巴细胞白血病儿童存在TELAML1融合基因并与预后相
关。 TELAML 1融合基因阳性ALL的特点①发病年龄。 ALL出现染色体t(12;21) 即形成TELAML1融合基因约占B系ALL 11%36%在成人中的检出率只有2%4%。 t(12;21)的儿童ALL多发生在110岁 15岁者占76.2% 14 。②免疫表型特征。所有的t(12;21)病人都表达B祖细胞免疫表型 以CD 19、 CD 10阳性最为多见偶见前前B表型未见成熟B细胞及T细胞表型15 。约有24.6%的t(12;21)病儿在20%以上原始细胞上共同表达至少2个髓系抗原CD 13、 CD 37、
CDW65 。 CD9和CD20常为阴性或部分阳性。 TELAML 1阳性的病人CD 10、人类白细胞抗原HLADR的阳性率为90%。 TELAML 1阳性的病例一般为前B ALL该型ALL约占总ALL的80%。③与预后的关系。TELAML 1表达阳性组属高危ALL的占13%而阴性组属高危ALL的占68%说明TELAML 1往往与低危ALL相关预后较好 TELAML 1阳性组地塞米松诱导实验阳性率为88%而阴性组仅为38% 阳性组地塞米松诱导实验效果明显优于阴性组。 由于地塞米松诱导实验体现了化疗的敏感性及其预后 TELAML 1阳性组的预后明显优于阴性病例16 。④微卫星不稳定性。 KIMBLER等17应用基因图谱采用微卫星标志检测81例初诊和缓解期ALL儿童TEL基因位点杂合性缺失LOH 。结果显示 15%的ALL病人显示TEL基因的LOH而经细胞遗传学分析12p缺失者仅为5%。 TEL区域LOH常发生于儿童且前B ALLs的发生率
40%远高于T ALLs 5% 。已存在LOH的ALL病人在明显的缺失区域包含两个候选抑癌基因即TEL和KIP1基因该基因编码cyclin依赖激酶抑制物P27。 TAKEUCHI等18检测38例复发ALL病人的L OH至少一条染色体出现LOH的占68%其中T EL基因位点发生LOH的占25%经序列分析初诊和复发病人在相同位点出现L OH的占
63%表现出相同的克隆改变。另外37%的复发病人表现为明显的LOH位点的改变表明复发时已出现异常克隆改变。两者虽结果不尽相同但都表现出候选基因的LOH表明候选抑癌基因的缺失在儿童ALL的复发中起到了至关重要的作用。
TELJAK2基因与ALL染色体t(9; 12) (p24;p 13)易位TEL基因的336氨基末端融合于JAK2的318羧基末端形成T ELJAK2融合基因主要发生在儿童T细胞白血病。 TELJAK2基因编码TELJAK2蛋白它对白细胞介素3依赖性Ba/F3造血细胞系发挥持续作用使其转化为白细胞介素3非依赖性Ba/F3造血细胞株造成该细胞系的持续增殖19 。这可能因TEL HLH结构域为JAK2的激酶结构域提供了二聚体化界面从而使JA K2不依赖配体而激活引起S T AT家族的DNA结合活性的持续存在造成造血细胞的恶性转化。体外动物模型显示 TELJAK2转基因小鼠及其转基因后代于422周均发展为CD8+T细胞白血病该类型白血病为白血病细胞呈高侵入性的克隆性疾病。
2.2 TEL基因与急性粒细胞白血病
TELABL基因与急性粒细胞白血病t(9; 12) (q34;p 13)易位是少见的白血病染色体结构畸变见于急性未分化型髓细胞白血病、非典型慢性粒细胞白血病aCML和ALL。该易位使TEL基因与正常位于9 q34上的ABL基因发生融合形成TELABL嵌合基因20 。 ABL蛋白是一种非受体型酪氨酸蛋白激酶 由TEL的HLH结构域和ABL的酪氨酸蛋白激酶活性域融合而成 TEL的HLH结构域促使ABL激酶寡聚化从而使ABL激酶激活。 TE LABL可使Ba/F3细胞获得I L3非依赖生长特
性并能转化原代小鼠骨髓细胞和Ra t 1纤维母细胞故较TELPDGFRβ有更强的转化能力。 PEETE等9在1例ALL病人发现t 9; 12(p24;p 13)易位形成一种新的融合基因TELJAK2。 TELJAK2的致白血病作用可能与TELPDGFR β一样是TEL的HLH结构域为JAK2激酶结构域提供了二聚化界面从而激活了JAK 2引起的。
MN1TEL基因与急性粒细胞白血病t 12;22 p13;q11易位见于AMLM1、 M4、 M7及MDS。该易位使22 q11上的MN1基因与TEL基因融合形成MN 1TEL融合基因 由于TEL基因的断裂点不同可产生两种融合基因。 TEL基因2号内含子断裂形成的Ⅰ型嵌合蛋白包含完整的TELHLH结构域和ETS结构域 当断裂点位于内含子3时形成的Ⅱ型嵌合蛋白含有断裂的TEL HLH和完整的ETS结构域。 由此表明 TEL HLH结构域在MN1TEL介导的白血病发生中并不重要而由ETS DNA结合结构域发挥主要作用。 MN1 TEL融合基因编码的嵌合蛋白作为异常转录因子使正常由TEL控制的基因出现调节异常而致细胞转化是白血病发生的原因。另一种与MN 1TEL结构类似的融合基因是t(4; 12) (q 11 q 12;p 13)所致的BTLTEL基因见于AMLM0型白血病。
T ELA RG基因与急性粒细胞白血病AMLM3和AMLM4E0病人可发现由染色体t(1; 12) q25;p 13易位形成的TELARG融合基因 ARG基因
即ABL Related Gene又称ABL2基因是与cABL同属Abelson家族的非受体酪氨酸激酶在TK、 SH2和SH3结构域具有高度的同源性。
2.3 TEL基因与慢性白血病
(5; 12) (q31 ;p13)染色体t(5; 12)易位见于嗜酸细胞增多型慢性粒单核细胞白血病CMML和aCML易位使TEL基因与位于5q31的PDGFR β基因融合即形成TELPDGFR β融合基因。 TELPDGFR β 引起白血病转化的机制在于激活PDGFR β激酶依赖的信号转导途径。单纯的TELPDGFR β是否足以引起细胞的恶性转化尚不肯定可能还需要其他的遗传学改变才能引起细胞的充分转化。临床上以各个不同阶段的嗜酸细胞增多和显著的骨髓纤维化为特征最后均发展为急性髓细胞白血病预后差。 BOURGEADE等5证实 M yc的转录激活对TELPDGFR β发挥细胞转化作用是必需的。类似的易位还有t 6; 12中的TELSTL和t(12; 15) (p 13;q25)中的TELTRKC这些融合基因都有一个共同的特点 即TEL保留其HLH结构域并位于融合基因的5′端编码酪氨酸激酶的基因往往位于融合基因的3′端而且激酶活性区仍然完整。当TEL的HLH结构域缺失后激酶就不能被激活。推测TELH LH结构域介导两相邻受体二聚体化使激酶不依赖配体而激活通过信号传导途径作用于相邻的转录因子和靶基因造成细胞的恶性转化可能是这一类白血病共同的发病机制。 t(9; 12) (q34;p13)易位形成TELABL融合基因亦在慢性髓细胞性白血1期王海燕谭齐贤TEL基因与白血病89病C ML病人中被发现。 CML的病程发展总是经历慢性期、加速期和急变期采用PCR技术检测其不同时期多个位点的微卫星的遗传不稳定性(MS I)和LOH可探索CML急变的原因并为临床治疗与病情监测提供帮助。有学者选择11、 18号及12号染色体TEL基因附近的位点检测CML 17例结果显示CML在加速期检出MSI的频率
7/9明显高于慢性期1/8 且2个以上位点的MS I占57. 1%
4/9 并观察到有多个位点的MS I的加速期病人易在较短的时间内
发生急变。说明MS I可能参与CML由慢性期转向急性期的演变。
2.4 TEL基因其他易位与白血病
TEL基因在白血病的发生中起重要的作用但TEL基因易位通过不同的分子机制引起白血病发生机制尚不完全清楚。其他TEL基因易位发生较少如t(5; 12) (q31 ;p13) t(6; 12; 17) (p21;p13;q25) t(7; 12) (p15;p13) t(7; 12) (p12;p13) t(7; 12) (q36;p13)t(12; 13) (p13;q12)易位分别在伴有明显嗜酸性粒细胞增多的C ML、儿童AM L、 MDS、 A LL及非何杰金淋巴瘤病人中检测到。 12p 13的断裂点多发生在TEL基因5′端上游外显子编码的HLH结构域处。
3 问题与展望
经过数年的努力对TEL基因在白血病发病机制中的作用做了大量的研究为阐明白血病的发病机制打开了一扇光明之门然而TEL基因仍给我们留下许多问题。如TEL基因的分子机制是什么如何抑制AML1调控基因的表达能否通过抑制激酶活性而进行白血病治疗这些问题的解决将有助于深入了解TEL相关白血病的发病机制并为临床的治疗提供新的思路。
参考文献
1 LODAKA I STARZA R L BAE M et al . Fluorescencein situ hybridization characterization of new tralocation
- 基因关于TEL基因与白血病相关文档
- 教学设计unit 3 could you please tel lme where the restrooms are-教学设计
- 展览公司以色列Historical Tel Aviv公寓
- 展位国药励展中国医药集团总公司和励展博览集团的成员公司Tel
- 基因关于儿童急性淋巴细胞白血病TEL基因微卫星不稳定性检测的临床意义
- 新加坡雅仕维(01993.HK)订立新加坡地铁TEL特许经营协议
- 淋巴细胞儿童急性淋巴细胞白血病与TEL-AML1融合基因关联分析
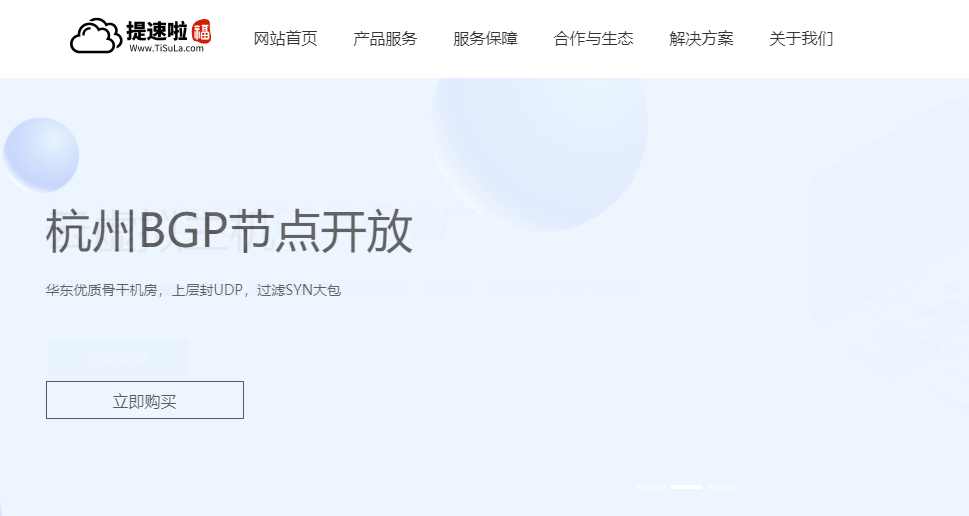
提速啦:美国多IP站群云服务器 8核8G 10M带宽 7IP 88元/月
提速啦(www.tisula.com)是赣州王成璟网络科技有限公司旗下云服务器品牌,目前拥有在籍员工40人左右,社保在籍员工30人+,是正规的国内拥有IDC ICP ISP CDN 云牌照资质商家,2018-2021年连续4年获得CTG机房顶级金牌代理商荣誉 2021年赣州市于都县创业大赛三等奖,2020年于都电子商务示范企业,2021年于都县电子商务融合推广大使。资源优势介绍:Ceranetwo...
妮妮云(119元/季)日本CN2 2核2G 30M 119元/季
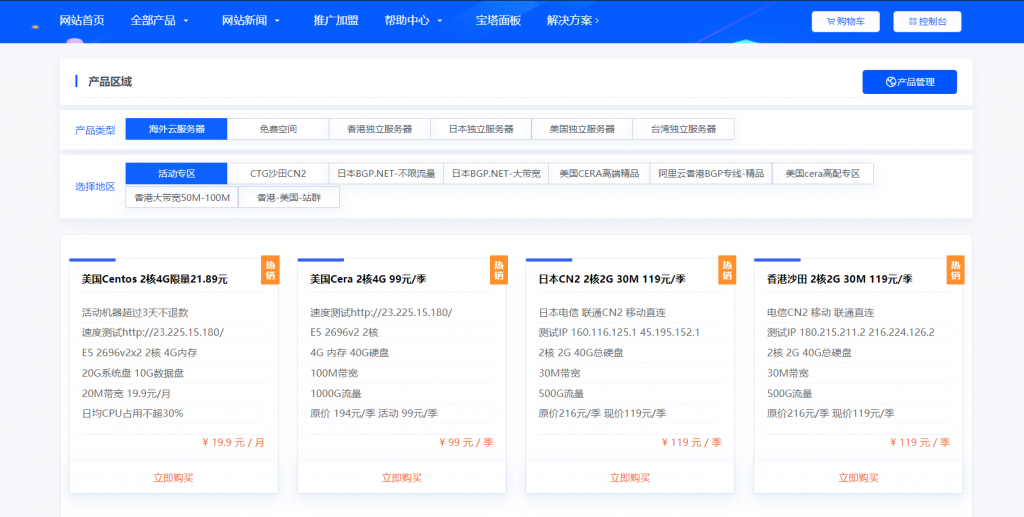
妮妮云的知名度应该也不用多介绍了,妮妮云旗下的云产品提供商,相比起他家其他的产品,云产品还是非常良心的,经常出了一些优惠活动,前段时间的八折活动推出了很多优质产品,近期商家秒杀活动又上线了,秒杀产品比较全面,除了ECS和轻量云,还有一些免费空间、增值代购、云数据库等,如果你是刚入行安稳做站的朋友,可以先入手一个119/元季付的ECS来起步,非常稳定。官网地址:www.niniyun.com活动专区...
星梦云-100G高防4H4G21M月付仅99元,成都/雅安/德阳
商家介绍:星梦云怎么样,星梦云好不好,资质齐全,IDC/ISP均有,从星梦云这边租的服务器均可以备案,属于一手资源,高防机柜、大带宽、高防IP业务,一手整C IP段,四川电信,星梦云专注四川高防服务器,成都服务器,雅安服务器,。活动优惠促销:1、成都电信夏日激情大宽带活动机(封锁UDP,不可解封):机房CPU内存硬盘带宽IP防护流量原价活动价开通方式成都电信优化线路2vCPU2G40G+60G21...
-
明星论坛www.51.com是一个关于什么的网站?找不到光驱电脑找不到光驱怎么办阿?缓冲区溢出教程哪里可以下载黑客教程,详细网址,吴晓波频道买粉《吴晓波频道》《罗辑思维》《专栏精粹》怎么评价?1433端口1433端口怎么打开安卓应用平台手机系统应用在哪唱吧电脑版官方下载唱吧有电脑版吗godaddyGodaddy域名怎么接受xp系统停止服务XP系统停止服务后怎么办?电子商务网站模板电子商务网站模板哪个好?电子商务网站模板免费建站怎么样?