骨骼肌变形计20140120
变形计20140120 时间:2021-03-25 阅读:()
www.
rehabi.
com.
cnChineseJournalofRehabilitationMedicine,Jan.
2014,Vol.
29,No.
1摘要目的:通过检测急性跑台运动中骨骼肌氧化应激情况以及Fas/FasL、Bcl-2/Bax等凋亡调控基因表达的动态变化,探究急性跑台运动诱导骨骼肌细胞凋亡的分子机制.
方法:以小鼠急性递增负荷跑台运动为实验模型,连续观察与分别测定安静组(Con)、急性跑台运动45min组(E45)、90min组(E90)、120min组(E120)和150min组(E150)的腓肠肌丙二醛(MDA)、过氧化氢(H2O2)、一氧化氮(NO)含量和超氧化物歧化酶(SOD)活性变化以及Fas/FasL、Bcl-2/Bax基因表达水平.
结果:①骨骼肌MDA和H2O2含量在急性跑台运动第45分钟时显著升高(P0.
05).
与Con组相比,E45组H2O2浓度明显增加,差异具有显著性(P0.
05),与E45组相比,E90、E120、E150组H2O2浓度明显下降(分别P<0.
05;P<0.
05;P<0.
01).
与Con组相比,E45、E150组NO含量明显增加,差异具有非常显著性(P<0.
01),E90、E120组含量也明显增加,差异具有显著性(P<0.
05).
见表1.
2.
2急性跑台运动对骨骼肌Fas/FasL、Bcl-2/Bax基因表达的影响与Con组相比,E90、E120组FasmRNA表达均明显上升,差异具有非常显著性(P<0.
01),与E12022www.
rehabi.
com.
cn2014年,第29卷,第1期组相比,FasmRNA表达明显下降,差异具有显著性(P<0.
05).
与Con组相比,E90、E120组FasLmRNA表达均明显上升,差异具有非常显著性(P<0.
01),与E45组相比,E120组FasLmRNA表达明显上升,差异具有非常显著性(P<0.
01),与E120组相比,E150组FasLmRNA表达明显下降,差异具有显著性(P<0.
05).
与Con组相比,E90、E120组Bcl-2mRNA表达均明显下降,差异具有显著性(P<0.
05),与E45组相比,E90组Bcl-2mRNA表达明显下降,差异具有显著性(P<0.
05).
与Con组相比,E45、E90、E120组BaxmRNA表达均显著性上升(分别P<0.
05;P<0.
01;P<0.
05),与E90组相比,E120、E150组BaxmRNA表达均显著性下降(分别P<0.
05;P<0.
01).
与Con组相比,E45、E90、E120组Bcl-2/Bax比值均显著性下降(分别P<0.
05;P<0.
01;P<0.
05).
见表2.
表1急性跑台运动中骨骼肌MDA、SOD、H2O2、NO动态变化情况(x±s)组别Con组E45组E90组E120组E150组与Con组比较:①P<0.
05;②P<0.
01;与E45组比较:③P<0.
05;④P<0.
01;与E90组比较:⑤P<0.
05MDA(nmol/mgprot)7.
221±0.
34612.
75±2.
387①13.
14±3.
259①10.
98±2.
7587.
182±0.
888③⑤SOD(U/mgprot)45.
39±3.
86640.
40±10.
0829.
67±5.
185①36.
33±8.
29635.
61±10.
02H2O2(mmol/gprot)63.
93±5.
111101.
7±19.
11①65.
40±11.
22③64.
81±11.
91③54.
89±6.
019④NO(mol/gprot)0.
804±0.
0902.
337±0.
237②2.
315±0.
487①2.
313±0.
499①2.
385±0.
510②表2急性跑台运动中骨骼肌Fas/FasL、Bcl-2/Bax基因表达的动态变化(x±s)组别Con组E45组E90组E120组E150组与Con组比较:①P<0.
05;②P<0.
01;与E45组比较:③P<0.
05;④P<0.
01;与E90组比较:⑤P<0.
05;⑥P<0.
01;与E120组比较:⑦P<0.
05Fas0.
853±0.
1161.
435±0.
1242.
362±0.
201②2.
853±0.
226②1.
342±0.
135⑦FasL0.
777±0.
2031.
343±0.
2282.
032±0.
298②2.
471±0.
246②④1.
447±0.
360⑦Bcl-21.
093±0.
2790.
916±0.
2560.
475±0.
133①③0.
526±0.
153①0.
725±0.
209Bax0.
857±0.
1501.
702±0.
215①2.
299±0.
286②1.
588±0.
220①⑤1.
191±0.
220⑥Bcl-2/Bax1.
103±0.
2700.
584±0.
112①0.
447±0.
109②0.
510±0.
124①0.
669±0.
2193讨论骨骼肌细胞中同时存在内源性抗氧化系统(CAT、GSH-Px、SOD等)与氧化系统.
正常的生理情况下,二者之间处于动态平衡状态.
MDA是脂质过氧化反应的重要产物,可衡量脂质过氧化程度.
SOD是氧自由基防御体系中重要的抗氧化酶,可通过歧化方式清除氧自由基,减轻细胞氧化损伤和凋亡.
H2O2是线粒体产生的活性氧,可分解成氧自由基,通过对生物膜多不饱和脂肪酸的过氧化而引起脂质过氧化反应,导致细胞氧化损伤和凋亡.
NO是一种多功能因子(氮自由基),可参与细胞多种生理功能的调节.
生理水平的NO可减少细胞凋亡,过量则促进细胞凋亡[5].
NO发挥抗凋亡还是促凋亡作用取决于其局部浓度.
通过检测骨骼肌MDA、H2O2、NO含量和SOD活性变化可衡量细胞氧化应激程度.
目前的研究已经充分证实,急性大强度运动可使骨骼肌MDA、H2O2、NO含量显著升高,SOD活性下降.
这是因为,急性运动是一种短时间、大强度的应激刺激,细胞调节机制尚未及时启动,氧化还原平衡被打破,使自由基过度生成与聚集,当其超过抗氧化系统的清除能力时,就会导致骨骼肌氧化损伤[6—7].
本研究结果显示,骨骼肌MDA、H2O2含量在急性跑台运动开始时即稳步上升,在45min时显著增高,这种高水平状态维持一段时间后又开始下降,直到运动结束.
而骨骼肌SOD活性在急性跑台运动90min时开始显著降低,随后又有所升高,直到运动结束.
可知,SOD活性的曲线走向与MDA、H2O2含量的变化态势基本相反.
而骨骼肌NO含量在急性跑台运动不同时相始终处于较高水平状态.
本研究结果与之前的研究结果基本一致.
这说明:急性跑台运动作为一种短时间、大强度的应激刺激,打破了骨骼肌氧化/抗氧化系统之间的动态平衡,使内源性抗氧化物不能有效清除自由基,从而引起脂质过氧化反应.
因此,本研究认为,骨骼肌氧化应激水平在急性跑台运动45min时显著上升,保持一段时间的高水平状态后又开始稳步下降,直到运动结束.
自由基过度生成引起的氧化应激是诱导细胞凋亡的重要信号途径[8].
胞浆中大量聚集的自由基可使氧化应激水平上升,通过激活Fas/FasL系统,直接23www.
rehabi.
com.
cnChineseJournalofRehabilitationMedicine,Jan.
2014,Vol.
29,No.
1启动细胞凋亡的死亡信号转导通路[9].
研究表明[10],H2O2所致的氧化应激可使c-myc、Fas/FasL等凋亡基因的表达明显增强,但经过SOD处理之后,c-myc、Fas/FasL、NF-κB的表达明显降低,凋亡指数也随之降低.
这说明,氧化应激诱导的细胞凋亡与激活NF-κB,促进凋亡相关基因c-myc、Fas/FasL的表达密切有关.
另外,氧化应激还能通过上调Bax基因的表达、抑制Bcl-2基因的表达,使线粒体膜异位,膜完整性受到破坏,细胞色素C(cytochromeC,Cyt-C)大量释放,后者通过与凋亡蛋白酶激活因子(apoptoticproteaseactivatingfactor1,Apaf1)及脱氧三磷酸腺苷结合,激活caspase9并与之协同顺序激活下游caspase3和caspase7,最终导致细胞凋亡[11—13].
Fas/FasL系统是骨骼肌细胞中直接启动细胞凋亡的死亡信号转导系统.
Fas又称Apo-1,属于肿瘤坏死因子受体和神经生长因子受体家族的细胞表面因子,在骨骼肌细胞凋亡中发挥转导凋亡信号的作用.
FasL配体是肿瘤坏死因子受体家族的细胞表面因子.
当骨骼肌细胞表面Fas表达时,可激活其配体FasL,导致Fas死亡域自聚或者交联,并通过激活神经鞘磷脂酶、酪氨酸蛋白激酶和白介素-1β转换酶等多种蛋白酶或内源性核酸酶,引起细胞凋亡.
Sandri等[14]研究表明,一次大强度运动可显著上调C57BL/6小鼠骨骼肌Fas/FasL、Bcl-2等凋亡相关基因的表达,使骨骼肌细胞凋亡.
也有研究表明,大负荷跑台运动(-10°,20m/min)可使腓肠肌SOD活性下降、MDA含量增加,Fas基因表达显著上调,而原花青素可显著降低氧化应激水平,对腓肠肌微损伤及凋亡有一定的缓解与抑制作用[15].
姜振等[16]研究表明,急性力竭运动可使大鼠骨骼肌FasmRNA表达增强、细胞凋亡数量增多,而高压氧疗法可通过增强抗氧化酶系统的活性、降低自由基水平、增加微循环血量、改善机体缺氧状态,从而抑制FasmRNA的表达,减少骨骼肌细胞凋亡,促进骨骼肌疲劳与微损伤的恢复.
本研究结果显示,急性跑台运动在诱导骨骼肌氧化应激的同时,还能使骨骼肌Fas/FasL基因表达水平在90min时显著增加,在120min时达到峰值,随后呈现下降态势.
这与之前的研究结果基本一致.
提示,急性跑台运动时自由基的过度聚集、氧化应激水平的提高可能激活Fas/FasL凋亡信号调控通路,使骨骼肌细胞发生凋亡.
Bcl-2/Bax是骨骼肌细胞凋亡过程中具有重要调控作用的基因.
Bcl-2表达的增强可抑制骨骼肌细胞凋亡,而Bax的过度表达则促进细胞凋亡.
Bcl-2/Bax比率是决定细胞是否发生凋亡以及凋亡程度的重要标志,因此被称为启动细胞凋亡的"分子开关".
高水平的Bcl-2/Bax有助于细胞存活,比率过低则促进细胞凋亡.
细胞凋亡与否最终取决于两者之间的平衡结果.
跑台训练实验也发现,当骨骼肌细胞发生凋亡时,抗凋亡基因Bcl-2和促凋亡基因Bax的表达均发生了显著变化,且二者之间的变化趋势恰好相反[17].
Podhorska-Okolow等[18]研究表明,持续一夜的自由跑轮运动可使C57BL/6小鼠骨骼肌Bcl-2/Bax比率显著降低,导致细胞凋亡信号的启动,但在运动后的恢复期,Bcl-2/Bax比率反而逐渐升高.
王冬梅等[19]研究发现,一次力竭性游泳运动可使骨骼肌Bcl-2mRNA表达下降8.
6%、BaxmRNA表达增加30.
5%,并认为,力竭性游泳可使骨骼肌抗凋亡能力减弱,促凋亡能力大大增强,并可能启动线粒体凋亡信号传导途径,引起细胞凋亡,造成骨骼肌疲劳或损伤.
柳佳伟等[20]研究表明,耐力训练可显著提高大鼠腓肠肌SOD、GSH-Px活性,降低氧化应激水平,增加Bcl-2mRNA表达,降低BaxmRNA表达;但一次力竭运动却可减弱SOD、GSH-Px活性,升高自由基水平,降低Bcl-2mRNA表达,增加BaxmRNA表达,而一次力竭运动后的补锌可显著提高腓肠肌GSH-Px活性和总抗氧化能力(totalantioxidantcapacity,T-AOC),增加腓肠肌Bcl-2mRNA表达,但对BaxmRNA表达的影响不明显.
陈德权等[21]研究表明,一次高强度的间歇运动可通过增强骨骼肌BaxmRNA表达、降低Bcl-2mRNA表达,升高自由基水平,共同使运动能力下降,而长期间歇训练可能通过增强骨骼肌抗氧化能力,上调抗凋亡基因的表达,使运动能力增强.
本研究结果显示,急性跑台运动在上调骨骼肌氧化应激水平的同时,还能使骨骼肌Bcl-2mRNA表达水平在90—120min时显著降低,随后又呈现上升态势;而BaxmRNA表达却在运动45min时显著增加,在90min时达到峰值,这种高水平态势一直维持到24www.
rehabi.
com.
cn2014年,第29卷,第1期120min后又开始稳步下降;且Bcl-2/Bax比率在运动45min时显著降低,在90min时达最低水平,维持到120min后呈现上升态势.
本研究结果与陈德权等人的研究结果一致.
提示,急性跑台运动时自由基的过度生成与聚集、氧化应激水平的提高可能通过调控骨骼肌Bcl-2/Bax基因的表达,促进细胞凋亡.
最近王雪芹等[22]的研究也同样认为,大强度递增负荷跑台运动可使自由基过度生成,抗氧化系统酶活性降低,导致抗凋亡基因Bcl-2mRNA表达下降,促凋亡基因BaxmRNA表达增加,其寡聚体形成并整合到线粒体上,引起线粒体外膜通透性改变,膜电位下降,游离的Cyt-C释放到胞浆,使线粒体凋亡通路的下游基因发挥凋亡调控作用.
急性跑台运动可使骨骼肌MDA、H2O2、NO含量显著上升、SOD活性下降,致使自由基过度生成与聚集,氧化应激水平提高.
急性跑台运动可显著增强骨骼肌Fas/FasL和Bax基因表达、降低Bcl-2基因表达和Bcl-2/Bax比率,使骨骼肌细胞发生凋亡,这可能与自由基过度聚集,氧化应激水平提高有关.
参考文献[1]ShuklaS,FujitaK,XiaoQ,etal.
Ashearstressrespon-sivegeneproductPP1201protectsagainstFas-mediatedapoptosisbyreducingFasexpressiononthecellsurface[J].
Apoptosis,2011,16(2):162—173.
[2]LimJY,HanTR.
Effectofelectromyostimulationonapopto-sis-relatedfactorsindenervationandreinnervationofratskeletalmuscles[J].
MuscleNerve,2010,42(3):422—430.
[3]ZhuX,SmithMA,HondaK,etal.
VascularoxidativestressinAlzheimerdisease[J].
JNeurolSci,2007,257(1—2):240—246.
[4]WuY,SongW.
RegulationofRCAN1translationanditsroleinoxidativestress-inducedapoptosis[J].
FASEBJ,2013,27(1):208—221.
[5]CorsettiG,PasiniE,AssanelliD,etal.
Acutecaffeinead-ministrationdecreasedNOSandBcl2expressioninratskele-talmuscles[J].
PharmacolRes,2007,55(2):96—103.
[6]张雪,丁树哲.
耐力训练对SD大鼠氧化应激及MAPKERK1/2的影响[J].
体育科学,2010,30(5):49—55.
[7]熊正英,刘军.
Ebselen对大强度耐力训练大鼠肝脏细胞凋亡及自由基代谢的影响[J].
体育科学,2008,28(11):62—67.
[8]Heath-EngelHM,ShoreGC.
Mitochondrialmembranedy-namics,cristaeremodellingandapoptosis[J].
BiochimBio-physActa,2006,1763(5—6):549—560.
[9]NowakM,KoppF,Roelofs-HaarhuisK,etal.
Oralnickeltolerance:Fasligand-expressinginvariantNKTcellspro-motetoleranceinductionbyelicitingapoptoticdeathofanti-gen-carrying,effeteBcells[J].
JImmunol,2006,176(8):4581—4589.
[10]MaheshwariA,MisroMM,AggarwalA,etal.
PathwaysinvolvedintesticulargermcellapoptosisinducedbyH2O2invitro[J].
FEBSJ,2009,276(3):870—881.
[11]Delivoria-PapadopoulosM,GornM,AshrafQM,etal.
ATPandcytochromec-dependentactivationofcaspase-9duringhypoxiainthecerebralcortexofnewbornpiglets[J].
NeurosciLett,2007,429(2—3):115—119.
[12]RoyAM,BaligaMS,ElmetsCA,etal.
Grapeseedproan-thocyanidinsinduceapoptosisthroughp53,Bax,andcas-pase3pathways[J].
Neoplasia,2005,7(1):24—36.
[13]TeijidoO,DejeanL.
UpregulationofBcl2inhibitsapopto-sis-drivenBAXinsertionbutfavorsBAXrelocalizationinmitochondria[J].
FEBSLett,2010,584(15):3305—3310.
[14]SandriM,Podhorska-OkolowM,GeromelV,etal.
Exer-ciseinducesmyonuclearubiquitinationandapoptosisindys-trophin-deficientmuscleofmice[J].
JNeuropatholExpNeu-rol,1997,56(1):45—57.
[15]闵柱,刘文杰,耿潇,等.
原花青素干预大负荷运动大鼠腓肠肌相关凋亡蛋白的表达[J].
中国组织工程研究,2013,17(2):309—314.
[16]姜振,张林,刘鹏程.
高压氧对急性力竭运动大鼠骨骼肌Fas-mRNA表达与细胞凋亡的影响[J].
安徽农业科学,2009,37(22):10520—10521.
[17]SongW,KwakHB,LawlerJM.
Exercisetrainingattenu-atesage-inducedchangesinapoptoticsignalinginratskele-talmuscle[J].
AntioxidRedoxSignal,2006,8(3—4):517—528.
[18]Podhorska-OkolowM,SandriM,ZampieriS,etal.
Apopto-sisofmyofibresandsatellitecells:exercise-induceddam-ageinskeletalmuscleofthemouse[J].
NeuropatholApplNeurobiol,1998,24(6):518—531.
[19]王冬梅,董军,漆正堂,等.
运动对骨骼肌线粒体通透性转换孔及细胞凋亡的影响[J].
西安体育学院学报,2011,28(4):471—475.
[20]柳佳伟,丁树哲,胡志刚.
补锌对耐力训练、力竭运动大鼠腓肠肌超氧化物岐化酶、谷胱甘肽过氧化物酶、总抗氧化能力以及Bax、Bcl-2mRNA的影响[J].
中国运动医学杂志,2011,30(8):740—744.
[21]陈德权,邓树勋,彭峰林.
间歇运动对大鼠骨骼肌凋亡基因Bax、Bcl-2及相关因素的影响[J].
北京体育大学学报,2008,31(7):922—925.
[22]王雪芹,郝选明.
6周递增负荷运动对大鼠小肠集合淋巴结细胞线粒体凋亡途径的影响[J].
体育学刊,2013,20(2):141—144.
25
rehabi.
com.
cnChineseJournalofRehabilitationMedicine,Jan.
2014,Vol.
29,No.
1摘要目的:通过检测急性跑台运动中骨骼肌氧化应激情况以及Fas/FasL、Bcl-2/Bax等凋亡调控基因表达的动态变化,探究急性跑台运动诱导骨骼肌细胞凋亡的分子机制.
方法:以小鼠急性递增负荷跑台运动为实验模型,连续观察与分别测定安静组(Con)、急性跑台运动45min组(E45)、90min组(E90)、120min组(E120)和150min组(E150)的腓肠肌丙二醛(MDA)、过氧化氢(H2O2)、一氧化氮(NO)含量和超氧化物歧化酶(SOD)活性变化以及Fas/FasL、Bcl-2/Bax基因表达水平.
结果:①骨骼肌MDA和H2O2含量在急性跑台运动第45分钟时显著升高(P0.
05).
与Con组相比,E45组H2O2浓度明显增加,差异具有显著性(P0.
05),与E45组相比,E90、E120、E150组H2O2浓度明显下降(分别P<0.
05;P<0.
05;P<0.
01).
与Con组相比,E45、E150组NO含量明显增加,差异具有非常显著性(P<0.
01),E90、E120组含量也明显增加,差异具有显著性(P<0.
05).
见表1.
2.
2急性跑台运动对骨骼肌Fas/FasL、Bcl-2/Bax基因表达的影响与Con组相比,E90、E120组FasmRNA表达均明显上升,差异具有非常显著性(P<0.
01),与E12022www.
rehabi.
com.
cn2014年,第29卷,第1期组相比,FasmRNA表达明显下降,差异具有显著性(P<0.
05).
与Con组相比,E90、E120组FasLmRNA表达均明显上升,差异具有非常显著性(P<0.
01),与E45组相比,E120组FasLmRNA表达明显上升,差异具有非常显著性(P<0.
01),与E120组相比,E150组FasLmRNA表达明显下降,差异具有显著性(P<0.
05).
与Con组相比,E90、E120组Bcl-2mRNA表达均明显下降,差异具有显著性(P<0.
05),与E45组相比,E90组Bcl-2mRNA表达明显下降,差异具有显著性(P<0.
05).
与Con组相比,E45、E90、E120组BaxmRNA表达均显著性上升(分别P<0.
05;P<0.
01;P<0.
05),与E90组相比,E120、E150组BaxmRNA表达均显著性下降(分别P<0.
05;P<0.
01).
与Con组相比,E45、E90、E120组Bcl-2/Bax比值均显著性下降(分别P<0.
05;P<0.
01;P<0.
05).
见表2.
表1急性跑台运动中骨骼肌MDA、SOD、H2O2、NO动态变化情况(x±s)组别Con组E45组E90组E120组E150组与Con组比较:①P<0.
05;②P<0.
01;与E45组比较:③P<0.
05;④P<0.
01;与E90组比较:⑤P<0.
05MDA(nmol/mgprot)7.
221±0.
34612.
75±2.
387①13.
14±3.
259①10.
98±2.
7587.
182±0.
888③⑤SOD(U/mgprot)45.
39±3.
86640.
40±10.
0829.
67±5.
185①36.
33±8.
29635.
61±10.
02H2O2(mmol/gprot)63.
93±5.
111101.
7±19.
11①65.
40±11.
22③64.
81±11.
91③54.
89±6.
019④NO(mol/gprot)0.
804±0.
0902.
337±0.
237②2.
315±0.
487①2.
313±0.
499①2.
385±0.
510②表2急性跑台运动中骨骼肌Fas/FasL、Bcl-2/Bax基因表达的动态变化(x±s)组别Con组E45组E90组E120组E150组与Con组比较:①P<0.
05;②P<0.
01;与E45组比较:③P<0.
05;④P<0.
01;与E90组比较:⑤P<0.
05;⑥P<0.
01;与E120组比较:⑦P<0.
05Fas0.
853±0.
1161.
435±0.
1242.
362±0.
201②2.
853±0.
226②1.
342±0.
135⑦FasL0.
777±0.
2031.
343±0.
2282.
032±0.
298②2.
471±0.
246②④1.
447±0.
360⑦Bcl-21.
093±0.
2790.
916±0.
2560.
475±0.
133①③0.
526±0.
153①0.
725±0.
209Bax0.
857±0.
1501.
702±0.
215①2.
299±0.
286②1.
588±0.
220①⑤1.
191±0.
220⑥Bcl-2/Bax1.
103±0.
2700.
584±0.
112①0.
447±0.
109②0.
510±0.
124①0.
669±0.
2193讨论骨骼肌细胞中同时存在内源性抗氧化系统(CAT、GSH-Px、SOD等)与氧化系统.
正常的生理情况下,二者之间处于动态平衡状态.
MDA是脂质过氧化反应的重要产物,可衡量脂质过氧化程度.
SOD是氧自由基防御体系中重要的抗氧化酶,可通过歧化方式清除氧自由基,减轻细胞氧化损伤和凋亡.
H2O2是线粒体产生的活性氧,可分解成氧自由基,通过对生物膜多不饱和脂肪酸的过氧化而引起脂质过氧化反应,导致细胞氧化损伤和凋亡.
NO是一种多功能因子(氮自由基),可参与细胞多种生理功能的调节.
生理水平的NO可减少细胞凋亡,过量则促进细胞凋亡[5].
NO发挥抗凋亡还是促凋亡作用取决于其局部浓度.
通过检测骨骼肌MDA、H2O2、NO含量和SOD活性变化可衡量细胞氧化应激程度.
目前的研究已经充分证实,急性大强度运动可使骨骼肌MDA、H2O2、NO含量显著升高,SOD活性下降.
这是因为,急性运动是一种短时间、大强度的应激刺激,细胞调节机制尚未及时启动,氧化还原平衡被打破,使自由基过度生成与聚集,当其超过抗氧化系统的清除能力时,就会导致骨骼肌氧化损伤[6—7].
本研究结果显示,骨骼肌MDA、H2O2含量在急性跑台运动开始时即稳步上升,在45min时显著增高,这种高水平状态维持一段时间后又开始下降,直到运动结束.
而骨骼肌SOD活性在急性跑台运动90min时开始显著降低,随后又有所升高,直到运动结束.
可知,SOD活性的曲线走向与MDA、H2O2含量的变化态势基本相反.
而骨骼肌NO含量在急性跑台运动不同时相始终处于较高水平状态.
本研究结果与之前的研究结果基本一致.
这说明:急性跑台运动作为一种短时间、大强度的应激刺激,打破了骨骼肌氧化/抗氧化系统之间的动态平衡,使内源性抗氧化物不能有效清除自由基,从而引起脂质过氧化反应.
因此,本研究认为,骨骼肌氧化应激水平在急性跑台运动45min时显著上升,保持一段时间的高水平状态后又开始稳步下降,直到运动结束.
自由基过度生成引起的氧化应激是诱导细胞凋亡的重要信号途径[8].
胞浆中大量聚集的自由基可使氧化应激水平上升,通过激活Fas/FasL系统,直接23www.
rehabi.
com.
cnChineseJournalofRehabilitationMedicine,Jan.
2014,Vol.
29,No.
1启动细胞凋亡的死亡信号转导通路[9].
研究表明[10],H2O2所致的氧化应激可使c-myc、Fas/FasL等凋亡基因的表达明显增强,但经过SOD处理之后,c-myc、Fas/FasL、NF-κB的表达明显降低,凋亡指数也随之降低.
这说明,氧化应激诱导的细胞凋亡与激活NF-κB,促进凋亡相关基因c-myc、Fas/FasL的表达密切有关.
另外,氧化应激还能通过上调Bax基因的表达、抑制Bcl-2基因的表达,使线粒体膜异位,膜完整性受到破坏,细胞色素C(cytochromeC,Cyt-C)大量释放,后者通过与凋亡蛋白酶激活因子(apoptoticproteaseactivatingfactor1,Apaf1)及脱氧三磷酸腺苷结合,激活caspase9并与之协同顺序激活下游caspase3和caspase7,最终导致细胞凋亡[11—13].
Fas/FasL系统是骨骼肌细胞中直接启动细胞凋亡的死亡信号转导系统.
Fas又称Apo-1,属于肿瘤坏死因子受体和神经生长因子受体家族的细胞表面因子,在骨骼肌细胞凋亡中发挥转导凋亡信号的作用.
FasL配体是肿瘤坏死因子受体家族的细胞表面因子.
当骨骼肌细胞表面Fas表达时,可激活其配体FasL,导致Fas死亡域自聚或者交联,并通过激活神经鞘磷脂酶、酪氨酸蛋白激酶和白介素-1β转换酶等多种蛋白酶或内源性核酸酶,引起细胞凋亡.
Sandri等[14]研究表明,一次大强度运动可显著上调C57BL/6小鼠骨骼肌Fas/FasL、Bcl-2等凋亡相关基因的表达,使骨骼肌细胞凋亡.
也有研究表明,大负荷跑台运动(-10°,20m/min)可使腓肠肌SOD活性下降、MDA含量增加,Fas基因表达显著上调,而原花青素可显著降低氧化应激水平,对腓肠肌微损伤及凋亡有一定的缓解与抑制作用[15].
姜振等[16]研究表明,急性力竭运动可使大鼠骨骼肌FasmRNA表达增强、细胞凋亡数量增多,而高压氧疗法可通过增强抗氧化酶系统的活性、降低自由基水平、增加微循环血量、改善机体缺氧状态,从而抑制FasmRNA的表达,减少骨骼肌细胞凋亡,促进骨骼肌疲劳与微损伤的恢复.
本研究结果显示,急性跑台运动在诱导骨骼肌氧化应激的同时,还能使骨骼肌Fas/FasL基因表达水平在90min时显著增加,在120min时达到峰值,随后呈现下降态势.
这与之前的研究结果基本一致.
提示,急性跑台运动时自由基的过度聚集、氧化应激水平的提高可能激活Fas/FasL凋亡信号调控通路,使骨骼肌细胞发生凋亡.
Bcl-2/Bax是骨骼肌细胞凋亡过程中具有重要调控作用的基因.
Bcl-2表达的增强可抑制骨骼肌细胞凋亡,而Bax的过度表达则促进细胞凋亡.
Bcl-2/Bax比率是决定细胞是否发生凋亡以及凋亡程度的重要标志,因此被称为启动细胞凋亡的"分子开关".
高水平的Bcl-2/Bax有助于细胞存活,比率过低则促进细胞凋亡.
细胞凋亡与否最终取决于两者之间的平衡结果.
跑台训练实验也发现,当骨骼肌细胞发生凋亡时,抗凋亡基因Bcl-2和促凋亡基因Bax的表达均发生了显著变化,且二者之间的变化趋势恰好相反[17].
Podhorska-Okolow等[18]研究表明,持续一夜的自由跑轮运动可使C57BL/6小鼠骨骼肌Bcl-2/Bax比率显著降低,导致细胞凋亡信号的启动,但在运动后的恢复期,Bcl-2/Bax比率反而逐渐升高.
王冬梅等[19]研究发现,一次力竭性游泳运动可使骨骼肌Bcl-2mRNA表达下降8.
6%、BaxmRNA表达增加30.
5%,并认为,力竭性游泳可使骨骼肌抗凋亡能力减弱,促凋亡能力大大增强,并可能启动线粒体凋亡信号传导途径,引起细胞凋亡,造成骨骼肌疲劳或损伤.
柳佳伟等[20]研究表明,耐力训练可显著提高大鼠腓肠肌SOD、GSH-Px活性,降低氧化应激水平,增加Bcl-2mRNA表达,降低BaxmRNA表达;但一次力竭运动却可减弱SOD、GSH-Px活性,升高自由基水平,降低Bcl-2mRNA表达,增加BaxmRNA表达,而一次力竭运动后的补锌可显著提高腓肠肌GSH-Px活性和总抗氧化能力(totalantioxidantcapacity,T-AOC),增加腓肠肌Bcl-2mRNA表达,但对BaxmRNA表达的影响不明显.
陈德权等[21]研究表明,一次高强度的间歇运动可通过增强骨骼肌BaxmRNA表达、降低Bcl-2mRNA表达,升高自由基水平,共同使运动能力下降,而长期间歇训练可能通过增强骨骼肌抗氧化能力,上调抗凋亡基因的表达,使运动能力增强.
本研究结果显示,急性跑台运动在上调骨骼肌氧化应激水平的同时,还能使骨骼肌Bcl-2mRNA表达水平在90—120min时显著降低,随后又呈现上升态势;而BaxmRNA表达却在运动45min时显著增加,在90min时达到峰值,这种高水平态势一直维持到24www.
rehabi.
com.
cn2014年,第29卷,第1期120min后又开始稳步下降;且Bcl-2/Bax比率在运动45min时显著降低,在90min时达最低水平,维持到120min后呈现上升态势.
本研究结果与陈德权等人的研究结果一致.
提示,急性跑台运动时自由基的过度生成与聚集、氧化应激水平的提高可能通过调控骨骼肌Bcl-2/Bax基因的表达,促进细胞凋亡.
最近王雪芹等[22]的研究也同样认为,大强度递增负荷跑台运动可使自由基过度生成,抗氧化系统酶活性降低,导致抗凋亡基因Bcl-2mRNA表达下降,促凋亡基因BaxmRNA表达增加,其寡聚体形成并整合到线粒体上,引起线粒体外膜通透性改变,膜电位下降,游离的Cyt-C释放到胞浆,使线粒体凋亡通路的下游基因发挥凋亡调控作用.
急性跑台运动可使骨骼肌MDA、H2O2、NO含量显著上升、SOD活性下降,致使自由基过度生成与聚集,氧化应激水平提高.
急性跑台运动可显著增强骨骼肌Fas/FasL和Bax基因表达、降低Bcl-2基因表达和Bcl-2/Bax比率,使骨骼肌细胞发生凋亡,这可能与自由基过度聚集,氧化应激水平提高有关.
参考文献[1]ShuklaS,FujitaK,XiaoQ,etal.
Ashearstressrespon-sivegeneproductPP1201protectsagainstFas-mediatedapoptosisbyreducingFasexpressiononthecellsurface[J].
Apoptosis,2011,16(2):162—173.
[2]LimJY,HanTR.
Effectofelectromyostimulationonapopto-sis-relatedfactorsindenervationandreinnervationofratskeletalmuscles[J].
MuscleNerve,2010,42(3):422—430.
[3]ZhuX,SmithMA,HondaK,etal.
VascularoxidativestressinAlzheimerdisease[J].
JNeurolSci,2007,257(1—2):240—246.
[4]WuY,SongW.
RegulationofRCAN1translationanditsroleinoxidativestress-inducedapoptosis[J].
FASEBJ,2013,27(1):208—221.
[5]CorsettiG,PasiniE,AssanelliD,etal.
Acutecaffeinead-ministrationdecreasedNOSandBcl2expressioninratskele-talmuscles[J].
PharmacolRes,2007,55(2):96—103.
[6]张雪,丁树哲.
耐力训练对SD大鼠氧化应激及MAPKERK1/2的影响[J].
体育科学,2010,30(5):49—55.
[7]熊正英,刘军.
Ebselen对大强度耐力训练大鼠肝脏细胞凋亡及自由基代谢的影响[J].
体育科学,2008,28(11):62—67.
[8]Heath-EngelHM,ShoreGC.
Mitochondrialmembranedy-namics,cristaeremodellingandapoptosis[J].
BiochimBio-physActa,2006,1763(5—6):549—560.
[9]NowakM,KoppF,Roelofs-HaarhuisK,etal.
Oralnickeltolerance:Fasligand-expressinginvariantNKTcellspro-motetoleranceinductionbyelicitingapoptoticdeathofanti-gen-carrying,effeteBcells[J].
JImmunol,2006,176(8):4581—4589.
[10]MaheshwariA,MisroMM,AggarwalA,etal.
PathwaysinvolvedintesticulargermcellapoptosisinducedbyH2O2invitro[J].
FEBSJ,2009,276(3):870—881.
[11]Delivoria-PapadopoulosM,GornM,AshrafQM,etal.
ATPandcytochromec-dependentactivationofcaspase-9duringhypoxiainthecerebralcortexofnewbornpiglets[J].
NeurosciLett,2007,429(2—3):115—119.
[12]RoyAM,BaligaMS,ElmetsCA,etal.
Grapeseedproan-thocyanidinsinduceapoptosisthroughp53,Bax,andcas-pase3pathways[J].
Neoplasia,2005,7(1):24—36.
[13]TeijidoO,DejeanL.
UpregulationofBcl2inhibitsapopto-sis-drivenBAXinsertionbutfavorsBAXrelocalizationinmitochondria[J].
FEBSLett,2010,584(15):3305—3310.
[14]SandriM,Podhorska-OkolowM,GeromelV,etal.
Exer-ciseinducesmyonuclearubiquitinationandapoptosisindys-trophin-deficientmuscleofmice[J].
JNeuropatholExpNeu-rol,1997,56(1):45—57.
[15]闵柱,刘文杰,耿潇,等.
原花青素干预大负荷运动大鼠腓肠肌相关凋亡蛋白的表达[J].
中国组织工程研究,2013,17(2):309—314.
[16]姜振,张林,刘鹏程.
高压氧对急性力竭运动大鼠骨骼肌Fas-mRNA表达与细胞凋亡的影响[J].
安徽农业科学,2009,37(22):10520—10521.
[17]SongW,KwakHB,LawlerJM.
Exercisetrainingattenu-atesage-inducedchangesinapoptoticsignalinginratskele-talmuscle[J].
AntioxidRedoxSignal,2006,8(3—4):517—528.
[18]Podhorska-OkolowM,SandriM,ZampieriS,etal.
Apopto-sisofmyofibresandsatellitecells:exercise-induceddam-ageinskeletalmuscleofthemouse[J].
NeuropatholApplNeurobiol,1998,24(6):518—531.
[19]王冬梅,董军,漆正堂,等.
运动对骨骼肌线粒体通透性转换孔及细胞凋亡的影响[J].
西安体育学院学报,2011,28(4):471—475.
[20]柳佳伟,丁树哲,胡志刚.
补锌对耐力训练、力竭运动大鼠腓肠肌超氧化物岐化酶、谷胱甘肽过氧化物酶、总抗氧化能力以及Bax、Bcl-2mRNA的影响[J].
中国运动医学杂志,2011,30(8):740—744.
[21]陈德权,邓树勋,彭峰林.
间歇运动对大鼠骨骼肌凋亡基因Bax、Bcl-2及相关因素的影响[J].
北京体育大学学报,2008,31(7):922—925.
[22]王雪芹,郝选明.
6周递增负荷运动对大鼠小肠集合淋巴结细胞线粒体凋亡途径的影响[J].
体育学刊,2013,20(2):141—144.
25
- 骨骼肌变形计20140120相关文档
- 膝关节变形计20140120
- suivante变形计20140120
- 股份变形计20140120
- Gregorian变形计20140120
- 中国社会福利基金会2018年度工作报告
- 芒果变形计20140120
如何低价香港服务器购买?有没有便宜的香港服务器推荐?
如何低价香港服务器购买?想要做一个个人博客,想用香港服务器,避免繁琐备案,性能不需要多高,只是记录一些日常而已,也没啥视频之类的东西,想问问各位大佬有没有低价的香港服务器推荐?香港距大陆近,相比美国服务器最大的优势在于延迟低,ping值低,但是带宽紧张,普遍都是1M,一般戏称其为“毛细血管”。同时价格普遍高,优质稳定的一般价格不菲。大厂云梯队阿里云、腾讯云两家都有香港服务器,要注意的是尽量不要选择...

Megalayer优化带宽和VPS主机主机方案策略 15M CN2优化带宽和30M全向带宽
Megalayer 商家主营业务是以独立服务器和站群服务器的,后来也陆续的有新增香港、菲律宾数据中心的VPS主机产品。由于其线路的丰富,还是深受一些用户喜欢的,有CN2优化直连线路,有全向国际线路,以及针对欧美的国际线路。这次有看到商家也有新增美国机房的VPS主机,也有包括15M带宽CN2优化带宽以及30M带宽的全向线路。Megalayer 商家提供的美国机房VPS产品,提供的配置方案也是比较多,...
六一云互联(41元)美国(24元)/香港/湖北/免费CDN/免费VPS
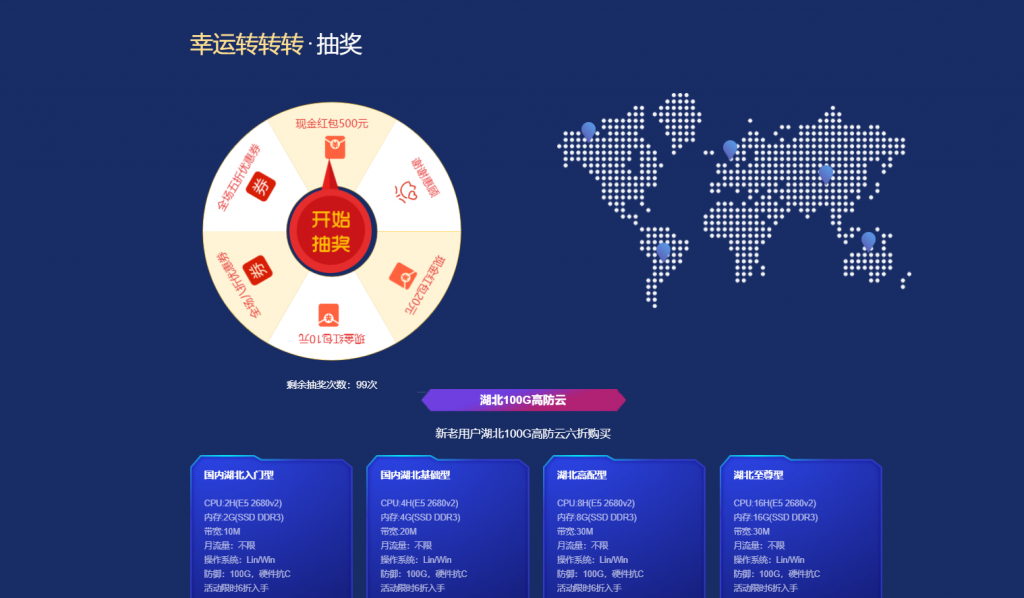
六一云互联六一云互联为西安六一网络科技有限公司的旗下产品。是一个正规持有IDC/ISP/CDN的国内公司,成立于2018年,主要销售海外高防高速大带宽云服务器/CDN,并以高质量.稳定性.售后相应快.支持退款等特点受很多用户的支持!近期公司也推出了很多给力的抽奖和折扣活动如:新用户免费抽奖,最大可获得500元,湖北新购六折续费八折折上折,全场八折等等最新活动:1.湖北100G高防:新购六折续费八折...
变形计20140120为你推荐
-
嘉兴商标注册个人如何申请商标注册刘祚天DJ是什么职业?杰景新特美国杰尼.巴尼特的资料xyq.163.cbg.com梦幻CBG的网站是什么。rawtoolsRAW是什么衣服牌子钟神发战旗TV ID:新年快乐丶未央不见是哪个主播5xoy.comhttp://www.5yau.com (舞与伦比),以前是这个地址,后来更新了,很长时间没玩了,谁知道现在的地址? 谢谢,www.zjs.com.cn中国快递公司排名avtt4.comCOM1/COM3/COM4是什么意思??/bbs2.99nets.com让(bbs www)*****.cn进入同一个站