larvalwww.39555.com
www.39555.com 时间:2021-03-18 阅读:()
Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46OpenAccessRESEARCHARTICLE2010Yinetal;licenseeBioMedCentralLtd.
ThisisanOpenAccessarticledistributedunderthetermsoftheCreativeCommonsAt-tributionLicense(http://creativecommons.
org/licenses/by/2.
0),whichpermitsunrestricteduse,distribution,andreproductioninanymedium,providedtheoriginalworkisproperlycited.
ResearcharticleAlternativetrans-splicingofCaenorhabditiseleganssma-9/schnurrigeneratesashorttranscriptthatprovidestissue-specificfunctioninBMPsignalingJianghuaYin,LingYuandCathySavage-Dunn*AbstractBackground:TranscriptioncofactorsrelatedtoDrosophilaSchnurrifacilitatethetranscriptionalprogramsregulatedbyBMPsignalinginC.
elegans,Drosophila,Xenopus,andmouse.
Indifferentsystems,SchnurrihomologshavebeenshowntoactaseitheragonistsorantagonistsofSmadfunction,andaseitherpositiveornegativeregulatorsoftranscription.
HowSchnurriproteinsachievethisdiversityofactivitiesisnotclear.
TheC.
eleganssma-9/schnurrilocusundergoesalternativesplicing,includinganunusualtrans-splicingeventthatcouldgeneratetwonon-overlappingshortertranscripts.
Results:Wedemonstrateherethattheshortertranscriptsareexpressedinvivo.
Furthermore,wefindthatoneoftheshorttranscriptsplaysatissue-specificroleinsma-9function,contributingtothepatterningofmale-specificsensoryrays,butnottotheregulationofbodysize.
Basedonpreviousresults,wesuggestthatthistranscriptencodesaC-terminalSMA-9isoformthatmayprovidetranscriptionalactivationactivity,whilefulllengthisoformsmaymediatetranscriptionalrepressionand/oractivationinacontext-dependentmanner.
Conclusion:Thealternativetrans-splicingofsma-9maycontributetothediversityoffunctionsnecessarytomediatetissue-specificoutputsofBMPsignaling.
BackgroundThetransforminggrowthfactorβ(TGFβ)superfamilycomprisesalargenumberofsecretedpeptidegrowthfac-torsthathavemajorregulatoryeffectsoncellgrowthanddifferentiation[1,2].
MembersofthissuperfamilyincludetheTGFβs,theprototypesofthesuperfamily;thebonemorphogeneticproteins(BMPs);andothermemberssuchasactivin,inhibin,andNodal.
TGFβsuperfamilyligandsbindtoaheteromericreceptorcomplexatthecellsurface.
Thiscomplexcontainstworelatedtransmem-braneserine/threoninekinases,thetypeIandtypeIIreceptors[3,4].
SignalingdownstreamofthereceptorsismediatedbytheSmadproteins,whichshuttlebetweenthecytoplasmandnucleustoregulatetargetgeneexpres-sion[5].
TypeIreceptorsdirectlyactivatereceptor-regu-latedSmads(R-Smads)byphosphorylationatC-terminalSXSsequences[6,7].
PhosphorylationofR-Smadspro-motesheterotrimericcomplexformationwithCo-Smadsandaccumulationinthenucleustoregulategenetran-scription[7-11].
Inmammals,fiveR-Smadsarepresent:two(Smad2,3)thattransduceTGFβ/activin/Nodalsig-nalsandthree(Smad1,5,8)thattransduceBMPsignals.
Strikingly,theTGFβfamilyligandsfaroutnumbertheSmadsavailableforsignaltransduction.
Furthermore,manyoftheseligandsarecapableofelicitingdiversecon-text-dependentresponses.
Thus,Smadcomplexesmustbecapableofmediatingmultiplediverseoutcomes.
ItisthoughtthatSmadcomplexesrelyinpartontranscrip-tioncofactorsforappropriateregulationoftargetgenes.
InthenematodeCaenorhabditiselegans,theBMP-relatedfactorDBL-1regulatesbodysizeandmaletailmorphogenesisviaaconservedreceptor/Smadsignalingpathway[12].
Usingageneticapproachtouncovercom-ponentsofthispathway,wepreviouslyidentifiedsma-9,agenethatisrequiredforthebodysizeandmaletailpat-terningfunctionsofDBL-1[13].
sma-9isalsorequiredforpatterningofthemesodermallineage,inwhichitacts*Correspondence:cathy.
savagedunn@qc.
cuny.
edu1DepartmentofBiology,QueensCollege,andBiochemistryPhDProgram,theGraduateSchoolandUniversityCenter,theCityUniversityofNewYork,Flushing,NY11367,USAFulllistofauthorinformationisavailableattheendofthearticleYinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page2of11antagonisticallytotheDBL-1pathway[14].
sma-9ispre-dictedtoencodemultipleproteinproductshomologoustoDrosophilaSchnurri,alargezincfingertranscriptioncofactorthatfunctionsinDpp/BMPsignaling[15,16].
Analysisofsma-9thereforeprovidestheopportunitytoelucidatetherequirementsfortranscriptioncofactorfunctioninaninvivomodelsystemduringthecourseofdevelopment.
InadditiontoDrosophilaSchnurriandC.
elegansSMA-9/Schnurri,threevertebrateSchnurrihomologshavebeenidentified.
ThesebindtheκB-bind-ingsiteandfunctioninTcelldevelopment[17-19].
Nota-bly,vertebrateSchnurrihomologshavemorerecentlybeendemonstratedtomediatetranscriptionalregulationdownstreamofBMPandTGFβligands,indicatingacon-servedroleforthesefamilymembersinTGFβsignaltransduction[20-22].
Thesma-9openreadingframe(ORF)predictedfromgenomicsequenceencodesaproteinof2170aaconsist-ingofanN-terminalGln-richdomain(encodedbypre-dictedexons1-3)andaC-terminaldomaincontainingsevenZnfingers(inpredictedexons12-22).
ThesequencingofcDNAclones,however,revealedatleasteightdifferentmRNAspecieswithalternativeproteincodingregions[13].
Interestingly,likesma-9,humanShn-1andShn-3genesundergoalternativesplicing[23,24],butthefunctionalconsequencesofthisprocess-inghavenotbeenaddressed.
Thesma-9cDNAcloneswereclassifiedbasedontheirpotentialtocodeforthesevenC-terminalZnfingers:classIencodesallsevenZnfingers(ZF1-7),classIIencodesthefirstpairandthetrip-letofZnfingers(ZF1-5),andclassIIIencodesonlythefirstpair(ZF1-2).
Todeterminewhetherthesedomainshavedifferentfunctionsinvivo,Foehretal.
createdgenomic/cDNAhybridconstructsandtestedtheirabilitytorescuethebodysizeandmesodermalpatterningdefectsofsma-9mutants[14].
Theyfoundthatcon-structsencodingclassIandclassIIC-terminiwerecapa-bleofrescuingbothphenotypes,whiletheclassIIIconstructwasnot,suggestingthatthepresenceoftheZnfingertripletiscriticalforfunctioninbodysizeandmesodermallineageregulation.
Sincesma-9isalargegene,mostoftheexistingcDNAclonescontainincompletecodingsequencesthataremissingthe5'endofthegene.
TwocDNAclones,yk1285a11andyk1237d01,however,arenotablydifferent(Figure1).
AlthoughthesetwocDNAclonesarelessthanhalfthelengthofthepredictedtranscript,bothofthemhaveallofthehallmarksofafull-lengthcDNA:trans-splicedleadersequence,poly(A)tail,andacompleteORF.
InC.
elegans,manytranscriptsareprocessedatthe5'endbytrans-splicing,whichresultsintheadditionofa22-nucleotidespliceleadersequence,SL1orSL2[25].
Themechanismoftrans-splicingissimilartothatofcis-splic-ing(intronremoval),exceptthatthesplicedonorsequencesareprovidedbytheSL1andSL2genes,ratherthanbeingcontainedwithinthecontextoftheindividualgene[25].
AbouthalfofallC.
elegansgenesaresubjectedtoSL1trans-splicingatthe5'end.
AsmallersubsetofC.
elegansgenesaretranscribedinpolycistronicoperons.
Inanoperon,the5'-mostgenegenerallyreceivestheSL1spliceleader.
TheindividualdownstreamgenesinanoperonarethenseparatedviaSL2trans-splicingtothe5'endsofeachofthedownstreamgenes.
SL2trans-splicingisaccompaniedbypolyadenylationtocreatethe3'endoftheneighboringgeneupstream.
yk1285a11containstheSL1spliceleaderandsequencesfrompredictedexons1-7.
yk1237d01containstheSL2spliceleaderandsequencesfrompredictedexons11-25.
Aspreviouslynoted[13],theputativeintercistronicregioncontainsfeaturesreminiscentoftheidentifiedsignalsfortrans-splicing,includingaU-richsequence,butisatypicallylong[26,27].
Thus,thestructuresofthesma-9cDNAclonesyk1285a11andyk1237d01suggestthattheymayrepresenttwotranscriptsprocessedfromasinglelongertranscriptbySL2trans-splicingandpolyadenylationasnormallyoccursinaC.
eleganspolycistronicoperon.
Sinceeachofthesetrans-splicedshortsma-9tran-scriptswasonlyrepresentedbyasinglecDNAclone,wecouldnotbecertainthatthecloneswerenotduetoclon-ingartifactsorararespuriousevent.
Furthermore,ourpreviousanalysisdidnotaddresswhethertheseshorttranscriptsprovideanyfunctionrequiredforDBL-1sig-naltransduction.
Wethereforeaddresshereseveralremainingquestionsaboutthesepredictedtranscripts.
First,canweverifythatthemRNAvariantsrepresentedbyyk1285a11andyk1237d01areexpressedinvivo,ratherthanbeingartifactsproducedduringcDNAlibrarycon-structionIfso,thenhowistheexpressionofthesevari-antsregulatedFinally,dothesevariantsshowfunctionaldifferencesbetweeneachotherand/orrelativetofull-lengthtranscriptsinvivoInthiswork,wewillrefertothesplicevariantsrepresentedbyclonesyk1285a11(5')andyk1237d01(3')asA11andD01,respectively.
ResultsIsolationofcDNAsspanningupstreamanddownstreamregionsThesma-9ORFpredictedfromgenomicsequenceisencodedby25exons([13];Figure1).
Previouslycharac-terizedcDNAclones,however,containedonlyasubsetofthese25exonsand,inparticular,noneoftheexistingcDNAclonesspannedaregionincludingboththeN-ter-minalGln-richdomainandtheC-terminalZnfingerdomains.
Wethereforeusedprimersinexons1and25withthepotentialtogeneratenearlyfull-lengthcDNAinsertsbyRT-PCR.
AfterRT-PCR,thecloneswiththelongestinsertswereselectedforsequencing.
ThemostcompletecDNAcloneisolatedbythisapproachwasYinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page3of11pCS368,inwhichexons4-8andpartofexon9aresplicedout(Figure1).
ThistranscriptformisreminiscentofaformidentifiedbytheORFeomeproject,inwhichexon3becomessplicedtoexon20[28].
Theseresultsthussup-porttheexistenceofsma-9transcriptscapableofencod-ingboththeGln-richandtheZnfingerdomains.
AdditionalsupportderivesfromRT-PCRdatausingexon9primers(seebelow).
Expressionofsma-9varianttranscriptsWenextsoughttoobtainevidencethattheshorttran-scriptsrepresentedbycDNAclonesyk1285a11andyk1237d01(A11andD01)areexpressedinvivo,ratherthancDNAartifacts.
Toaddressthisquestion,weneededanapproachtodistinguishtheseshorttranscriptsfromfull-lengthtranscriptscontainingthesameinternalsequences.
Unfortunately,wewereunabletodetectsma-9mRNAsbyNorthernblot(datanotshown).
Instead,weusedtheuniqueterminioftheshorttranscriptstodesignvariant-specificprimerpairsforRT-PCR.
ForA11weemployedanoligo-dT-containingreverseprimeranchoredbytwonucleotidesfromsma-9exon7withagene-specificinternalforwardprimer;forD01weusedanSL2-containingprimeranchoredbytwonucleotidesfromsma-9exon11withagene-specificinternalreverseprimer.
TodetermineempiricallywhethertheseprimerpairsarespecificforA11andD01,weperformedRT-PCRusingagradientofannealingtemperatures(50°C-55°C)andanalyzedtheproducts(Figure2A).
Inbothcases,theprimerpairsamplifiedasinglebandofthecor-rectsize(330bpforA11and300bpforD01),indicatingthattheprimersspecificallydetectthedesiredtran-scripts.
Sincethehigherannealingtemperaturesresultedinreducedyield,weused50°Cforallsubsequentanaly-ses.
Basedontheseresults,weconcludethattranscriptspolyadenylateddownstreamofexon7andtranscriptscontainingtheSL2spliceleadersequenceupstreamofexon11arerepresentedinthepoolofsma-9mRNAsinvivo.
TheinitialRT-PCRexperimentswereperformednon-quantitativelyonmixed-stageRNApreparations.
Wesubsequentlyusedreal-timequantitativeRT-PCR(qRT-PCR)todeterminetherelativeexpressionlevelsofsma-9varianttranscriptsandwhethertheyaredevelopmentallyorsex-specificallyregulated.
Sinceweexpectfull-lengthtranscripts,butnottheshorttranscripts,tocontainsequencesfromexon9,weusedexon9internalprimerstodeterminetheexpressionlevelsofpotentiallyfull-lengthtranscriptsandcalculatedtheabundanceoftheshorttranscriptsrelativetoexon-9-containingtran-scripts.
Threedevelopmentaltimepointswereassayed:24hr(firstlarvalstage-L1),44hr(approximatelyL3),and96hr(adults).
Wefindthatexon-9-containingtran-scriptlevelsincreaseduringlarvaldevelopmentandarehighestinadulthood(Figure2B).
D01transcriptsaccu-mulateinwild-typestrainN2at45%-85%ofthelevelofexon-9-containingtranscriptsatalldevelopmentaltimepointsexamined,whileA11transcriptsaccumulateatapproximately10%ofthelevelofexon-9-containingtran-scripts(Figure2B;Table1).
IfA11andD01areproducedconcurrentlybytrans-splicingoffull-lengthtranscripts,thenthereducedaccumulationofA11maybeduetodif-Figure1Structureofsma-9transcriptvariants.
Thepredictedsma-9intron-exonstructureisshownabove.
Arrowsmarkpositionsofnonsensecodonsinthreesma-9alleles.
pCS386isthemostcompletecDNAvariantisolatedtodate.
ThetwoshortvariantsrepresentedbycDNAclonesyk1285a11(A11)andyk1237d01(D01)areshownbelow.
ARD:acidicresidue-richdomain;NLS:nuclearlocalizationsignal.
pCS368pCS368Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page4of11Figure2Expressionofsma-9transcriptvariants.
A.
DetectionofA11andD01transcriptsusingvariant-specificRT-PCRoveragradientofannealingtemperatures(lefttoright:50°C-55°C).
TheexpectedsizeofthePCRproductis330bpforA11and300bpforD01.
B.
Developmentalprofileofsma-9transcriptexpressionlevels,asdeterminedbyqRT-PCR.
Dataareshownastranscriptabundancerelativetoact-1actingenecontrol,anderrorbarsshowstandarddeviation.
SeeTable1forquantitationoftranscriptabundancerelativetoexon-9-containingcontrol.
ABA11D010501001502002503003502444962444HoursAfterEmbryoCollectionTranscriptLevel(relativetoactincontrol)A11exon9D01N2him-5ferentialstabilityofthetranscripts.
Onepossiblemecha-nismforreducedstabilityoftheA11transcriptistheunusuallocationoftheterminationcodonTAA,inwhichtheterminaladeninesareintroducedduringpolyadenyla-tionandarecontiguouswiththepoly(A)tail.
Totestforsex-specificexpressionofthesetranscripts,him-5(highincidenceofmales)mutantpopulationsthatcontainbothhermaphroditesandmaleswerecomparedtowild-typeN2hermaphroditepopulations.
Inhim-5populations,thefull-lengthandtheD01transcripts,butnotA11,accumulatetoa2-4-foldhigherlevelthaninhermaphro-ditepopulations,suggestingthatthesetranscriptsareexpressedatahigherlevelinmales.
Theshorttranscriptsarenotmale-specific,however,sincetheyarealsoexpressedinhermaphrodites.
Thus,theA11andD01cDNAclonesrepresentdetectablemRNAsthatareexpressedduringlarvalandadultstageswhenDBL-1isactiveinregulatingbodysizeandmaletailpatterning.
Lossoffunctionofsma-9varianttranscriptsPreviousworkhasshownthatsma-9constructslackingtheZnfingertriplet(ZF3-5)areunabletorescuethebodysizeandmesodermalpatterningdefectsofasma-9mutant[14].
Theseexperimentsdidnottest,however,fordifferentialrequirementsofshortvs.
full-lengthtran-scripts.
Totestwhethersma-9varianttranscriptshavespecificfunctionalroles,weperformedRNAitargetingthreedifferentregionsofthesma-9gene.
Inpreviousexperiments,dsRNAwasintroducedbymicroinjection[13,14],butwehavefoundthatRNAibyfeedingresultsinahigherpenetranceofmaletaildefects.
Wehavethere-forerepeatedtheseexperimentsusingthefeedingtech-nique[29],andextendedthembytargetingtheinternalexon9inadditiontothe5'and3'endsofthegene.
TheeffectivenessofeachRNAitreatmentwasmonitoredbytheappearanceofthecharacteristicsmallbodysizephe-notype(datanotshown).
AsacomponentintheDBL-1pathway,sma-9mutantphenotypesincludesmallbodysizeandmaleabnormalphenotypes[13].
Oneaspectofthemaleabnormalphe-notypeinsma-9isthefusionofmaletailsensoryrays8and9,twooftheninebilaterallysymmetricalpairsofmale-specificsensoryorgans.
Inwild-typeanimals,ray8-9fusionsarealsoobserved,butatalowerfrequency.
Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page5of11Treatmentofwild-typeanimalswithRNAitargetingexons1-7causednoincreaseinthefrequencyofray8-9fusions(Table2),althoughthesmallbodysizephenotypewasproduced.
Thistreatmentispredictedtoknockdownexpressionoffull-lengthandA11variantsbutnottheD01variant.
Conversely,RNAitargetingexons21-25,predictedtoknockdowntheD01andfull-lengthvariants,resultedinatwo-foldincreaseinthefrequencyofray8-9fusions(Table2).
TheseresultsareconsistentwithourpreviousfindingsthatRNAiofthe3'endofthegeneresultsinmoreseveremaleabnormalphenotypesthaninactivationofthe5'end[13].
WenextcomparedthemaletailphenotypeproducedbyRNAitargetingthecen-tralexon9,predictedtoinhibitfull-lengthtranscriptsbutnottheshorttranscripts.
ThistreatmentresultedinamaletaildefectoflowerpenetrancethanthatproducedbyRNAitargetingthe3'end(Table2).
SincethemostseveremaletaildefectoccurredwhenD01wastargeted,theseexperimentssuggestthatD01hasaroleinregulat-ingsensoryraypatterning.
Full-lengthsma-9productsarealsopredictedtofunctioninsensoryraypatterning,sincetheinhibitionofacentralexonresultedinapartialmaleabnormalphenotype.
Wesoughttorepeatthisanalysisusinggeneticmutants,sincetheRNAitreatmentsmightcausesomenonspecificeffectsduetotargetingofunsplicedpre-mRNAortoRNAispreading[30,31].
Wetookadvantageofthreesma-9allelescontainingprematureterminationcodonsatdifferentlocationsinthegene(Figure1;[13,14]):qc3inpredictedexon1(withinA11),wk55inpredictedexon9(betweentheshortvariants),andcc604inpredictedexon18(withinD01).
Therefore,inqc3,onlytheD01transcriptiscapableofencodingafunctionalgeneproduct;inwk55,bothA11andD01,butnotthefull-lengthtranscript,arecapableofencodingfunctionalgeneproducts;andincc604,onlytheA11transcriptiscapableofencodingafunctionalgeneproduct.
Sinceadditionaldisruptionsmayoccurduetononsense-medi-ateddecay[32],wedeterminedthetranscriptexpressionprofileforthesestrainsattheL3stage(Figure3;Table1).
Inallthreemutantbackgrounds,thetranscriptlevelsaresomewhatreducedcomparedtothehim-5control.
Thereductionintranscriptlevelsismoststrikinginthesma-9(wk55)background(Figure3).
Thisseverereductionintranscriptlevelsisconsistentwithourobservationsthatthisallelecausesthemostseverebodysizephenotype(Table2;[13]).
Mostimportantlyfortheinterpretationofourexperiments,sma-9(qc3)andsma-9(cc604)animalsexpresstheshorttranscriptsatdetectablelevelsnearthenormalrange(Figure3).
Theusualexpectationforaseriesofnonsensemuta-tionsisthattheearliestprematureterminationcodonwillhavethemostseveremutantphenotypewhilelaterter-minationcodonsmaycauselessseveredefectsifthoseallelesencodepartiallyfunctionalgeneproducts.
Forthesma-9alleles,theoppositeresultisseeninthemaletail.
Thealleleencodingthelatestterminationcodon,cc604,causesthemostseveremaletaildefect,withafrequencyofray8-9fusionsgreaterthananyofthepreviouslychar-acterizedalleles(Table2,[13]).
Sincedifferentinvestiga-torstypicallyvaryintheirquantitationofrayfusionfrequencies,wealsorepeatedmeasurementofthemaleTable1:Relativeexpressionlevelsofsma-9transcriptvariantsStrainDevelopmentalTimepointA11TranscriptLevelD01TranscriptLevelN2L110.
12±1.
0686.
35±1.
17N2L38.
88±2.
5845.
00±0.
87N2adult8.
97±0.
4384.
58±7.
82him-5L18.
23±0.
2887.
01±0.
37him-5L32.
18±0.
0037.
73±0.
11sma-9(qc3);him-5L36.
20±0.
0149.
55±1.
29sma-9(wk55);him-5L31.
81±0.
0988.
50±1.
32sma-9(cc604);him-5L39.
40±0.
0562.
70±1.
26Meanexpressionlevelsaregiveaspercentoftheexpressionlevelofexon-9-containingcontrol,plusorminusstandarddeviation.
Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page6of11taildefectsofqc3andwk55fordirectcomparison.
Con-sistentwithpreviousresults,qc3,theallelecontainingtheearliestterminationcodon,causesamildmaletaildefect,whilewk55,containingaterminationcodoninexon9,resultsinanintermediatefrequencyofrayfusions(Table2).
Thus,theseverityofthesensoryraydefectisinverselycorrelatedwithpredictedD01activity.
Themostseveredefectismanifestedincc604mutants,inwhichtheD01isoformisdisruptedbymutation.
Anintermediatefre-quencyofrayfusionsoccursinwk55mutants,inwhichtheD01transcriptisexpressedatreducedlevelsbutisnotdisruptedbymutation.
Amildmaletaildefectisseeninqc3mutants,inwhichlevelsofD01transcriptsarelowerthaninwildtypebuthigherthaninwk55(Figure3).
Overall,withregardtomaletailpatterning,thesemutationsformanallelicseriesinwhichtheirlevelsofactivityaretheoppositeofthatexpected.
Inbodysize,adifferentresultwasobtainedwiththesethreemutants(Table2).
Thesma-9(wk55)mutantshowsthesmallestbodysize.
Incontrast,bothqc3andcc604producelessseverebodylengthphenotypesthataremutuallyindistinguishable(Table2).
Sinceqc3andcc604produceindistinguishablebodysizedefects,thisanalysisdoesnotsupportaspecificroleforeithertheA11ortheD01transcriptintheregulationofbodysize.
Further-more,themoreseverebodysizephenotypeofwk55mutantscouldbeduetotheloweroveralllevelofsma-9expressioninthesemutants(Figure3).
Theresultsofbodysizeanalysisthussuggestthatfull-lengthisoforms,ratherthanA11orD01,arecriticalforpromotinggrowth.
BasedonourRNAidataandonthedifferentialeffectsofthethreenonsenseallelesonbodysizeandmaletaildevelopment,wehypothesizethattheD01shorttran-scripthasatissue-specificfunctioninsensoryraydevel-opment.
Rescueofsma-9mutantphenotypesbyoverexpressionofD01cDNAConsistentwithourhypothesis,wehavepreviouslydem-onstratedthatoverexpressionoftheD01cDNA,butnotofA11,fromaheatshockpromotercanpartiallyrescuesma-9(wk55)mutantmaletaildefects[33].
Incompari-son,expressionofsma-9fromacosmidgenomicclonecontainingtheentirecodingregionresultedinnearlycompleterescueoftherayfusiondefect[13].
Wenextaskedwhetherexpressionofsma-9shorttranscriptsissufficienttoprovidenormalgenefunctioninqc3andincc604mutants,inwhichtheA11andtheD01transcript,respectively,aredisruptedbyprematureterminationcodons.
Asbefore,transgenicscarryinghs-sma-9con-structsweresubjectedtoheatshockduringtheL3stagewhensensoryrayidentitiesarebeingestablished[34]andmaleswerescoredinadulthood.
Incc604mutants,whichcontainaprematureterminationcodonwithintheD01transcript,expressionofD01resultedinpartialrescueofthemaletaildefect(Figure4A).
Incontrast,overexpres-sionofA11incc604causedaslightreductioninfre-quencyofsensoryrayfusions,butthiseffectwasnotstatisticallysignificant(p=0.
16).
Inqc3mutants,inwhichtheA11transcriptcontainsaprematuretermina-tioncodon,weweresurprisedtofindthatoverexpressionofA11producednochangeinmaletailphenotype,whereasoverexpressionofD01resultedinsignificantres-cueofthemaletaildefect(Figure4A).
Basedontheres-Table2:sma-9lossoffunctionphenotypesinmaletailpatterningandbodysizeRNAi(Exon)orMutantFormsPredictedtobeBlockedFrequencyofRay8-9Fusions1(n)BodyLength2(n)nonenone18%(50)ND1-7full-length,A1120%(50)ND9full-lengthonly29%(100)ND21-25full-length,D0136%(50)NDsma-9(qc3)full-length,A1150%(50)692±85(43)sma-9(wk55)full-lengthonly66%(50)566±34(31)sma-9(cc604)full-length,D0172%(50)716±61(30)1Allmaletailphenotypesarescoredwithhim-5(e1490)inthebackgroundtoincreasethefrequencyofmales.
2Bodylengthisgiveninμm±standarddeviationat96hoursafterembryocollection.
ND:notdetermined.
Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page7of11cueofmaletaildefectsbyD01inbothqc3andcc604mutants,weconcludethatincreasedexpressionofD01maybeabletocompensatepartiallyfordefectivefull-lengthsma-9transcripts.
Finally,welookedforevidenceofA11orD01functioninbodysizeregulation.
Thebodysizephenotypesofqc3andcc604didnotsupportaspecificroleforeitherA11orD01inbodysizeregulation,sincethesetwomutantshaveindistinguishablesizes(Table2).
TodeterminewhetherincreasedexpressionofA11and/orD01canpromotegrowth,weusedourheatshockconstructstodrivetheirexpression.
HeatshockoftransgenicsintheL3stagepro-ducednoevidentchangesinbodysize(datanotshown),soweperformedheatshockearlierindevelopment,inL1animals,andmeasuredbodylengthinadulthood.
Incon-trasttotherescueofmaleabnormalphenotypesbyD01,heatshockofA11andD01transgenicsledeithertoareductioninbodysizeortonochangeinbodysize(Fig-ure4B).
TheseresultssuggestthatA11andD01donotcontributetothegrowthpromotingactivityofsma-9.
Ifanything,increasedexpressionofA11andD01mayinterferewiththeabilityofgeneproductsfromfull-lengthtranscriptstopromoteincreasesinbodysize.
DiscussionWehaveshownherethattheC.
eleganssma-9/schnurrilocusundergoesanoveltrans-splicingprocesstogener-atetwoshortertranscripts,oneofwhichhasatissue-spe-cificfunctioninDBL-1/BMPsignaling.
First,wehavedemonstratedthatanSL2trans-spliced,truncatedcDNAform,yk1237d01(D01),representsanactualmRNAthatisdetectablebyRT-PCR.
Second,wehaveverifiedthatlongertranscriptscontainingboththe5'and3'endsofthegenearealsodetectablebyRT-PCR.
Third,loss-of-functionexperiments(RNAiandprematureterminationmutants)demonstratethattheshorttranscriptD01isnecessarybutnotsufficientformaletailpatterning,sincedepletionofbothD01andfull-lengthtranscriptscausesamoreseveremaletailpatterningdefectthandepletionoffull-lengthtranscriptsalone.
Fourth,gain-of-functionexperiments(heatshock-inducedoverexpression)indi-catethattheshorttranscriptD01canpartiallyrescueFigure3Expressionlevelsofsma-9transcriptsinmutantbackgrounds.
Expressionlevelsofsma-9transcriptsasdeterminedbyqRT-PCR.
AnimalswerecollectedattheL3stage.
Dataareshownastranscriptabundancerelativetoact-1actingenecontrol,anderrorbarsshowstandarddeviation.
SeeTable1forquantitationoftranscriptabundancerelativetoexon-9-containingtranscripts.
(-):transcriptharborsaprematureterminationcodon.
(+):transcriptsequenceisnormal.
050100150200250300350sma-9(qc3);him-5sma-9(wk55);him-5sma-9(cc604);him-5him-5TranscriptLevel(relativetoactincontrol)A11exon9D01(+)(+)(+)(+)(+)(+)(-)(-)(-)(-)(-)Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page8of11maletailbutnotbodysizedefectsofsma-9mutants.
IncontrasttotheresultsonD01,wefindfortheyk1285a11(A11)transcriptlittleornoevidenceofaninvivofunc-tion,aswellaslowabundancethroughoutdevelopment.
Inaddition,classIandIIfull-lengthtranscriptsandtheD01shorttranscript,butnottheA11transcript,encodetheZnfingertripletdomainshowntobecriticalintheregulationofbodysizeandmesodermalpatterning[14].
InDrosophila,thisdomainisalsocriticalinmediatingDpp-responsivetargetgeneregulation[35].
ThegeneproductencodedbytheA11transcriptlacksthisdomainandmaythereforenotbecapableofbeingrecruitedtotargetgenes.
WhythenistheA11transcriptdetectedatallOnepossibilityisthatitisgeneratedasabyproductofthetrans-splicingmechanismthatgeneratesD01.
Sincewehaveshownthatfull-lengthtranscriptsandD01transcriptsareeachnecessarybutnotsufficientformaletailpatterning,wemustconsiderwhatdifferencesinmolecularfunctionarefoundintherespectivegeneprod-ucts.
Inotherorganisms,Schnurrihomologshavebeenshowntoacteitherastranscriptionalactivators[20,21,36]orastranscriptionalrepressors[16,37]mediat-ingBMP-dependenttranscription.
Strikingly,Yaoetal.
[21]haveshownbyswappingDrosophilaandXenopusSchnurrihomologsthattranscriptionalactivationorrepressionactivitiesmaybecontext-dependentratherthananintrinsicpropertyoftheprotein.
InC.
elegans,wehavepreviouslyanalyzedintrinsictranscriptionalactivi-tiesofSMA-9proteindomainsusingaheterologoustran-scriptionassay[33].
Intheseexperiments,theacidicresidue-richdomain(ARD)inexon14exhibitedtran-scriptionalactivatoractivity,whiletheN-terminalregionofSMA-9displayedintrinsictranscriptionalrepressoractivity[33].
Full-lengthsma-9transcriptsshouldencodeproteinproductscontainingbothofthesetranscriptionaldomains,sothattheirtranscriptionalactivitiesmaybecontext-dependentaswasdescribedforDrosophilaandXenopusSchnurri[21].
Ontheotherhand,D01tran-scriptsencodeproductscontainingsolelythetranscrip-tionalactivationdomain(Figure1).
Wethereforehypothesizethatfull-lengthSMA-9isoformsfunctionastranscriptionalrepressorsoractivatorsdependingoncontext,whileD01-encodedisoformsmaybeobligatetranscriptionalactivators.
Thistranscriptionalactivationactivitymaybenecessaryfortherobustactivationofsma-9targetgenesinsensoryraydevelopment,butmaybedispensableinbodysizeregulation.
Consistentwiththishypothesis,aSMA-9Znfingerdomainfusionwithaknowntranscriptionalactivator,VP16,canpartiallyres-cuethemaletaildefectsbutnotthebodysizedefectsofsma-9mutants.
IncontrasttotheSMA-9::VP16fusion,aSMA-9::enRrepressiondomainfusionrescuesbodysize,suggestingthattranscriptionalrepressionismoreimpor-tantthanactivationforSMA-9activityinbodysizeregu-lation[33].
Overall,theinvivoactivityoftheSMA-9::VP16fusionisstrikinglysimilartothatoftheD01iso-form.
Alternativesplicingisregulatedbytrans-actingfactorsthatinfluencethechoiceofsplicesites.
Itwillbeofinter-esttodeterminethetrans-actingfactorsthatregulatealternativetrans-splicingofsma-9.
Inpreliminaryexperi-ments,wetestedwhethersma-9trans-splicingdependsonmec-8,whichencodesanRNArecognitionmotif-con-tainingproteinthatregulatesalternativesplicinginthehypodermis[38,39].
WefoundthatA11,D01,andexon9-containingtranscriptlevelswereallreduced2-to5-foldinmec-8mutantsrelativetoN2(datanotshown).
Ifmec-8weredirectlyinvolvedinprocessingtheshorttran-Figure4Phenotypicrescuebyoverexpressionofsma-9shorttranscripts.
A.
Maletailphenotypesaregivenasfrequencyoffusionofrays8and9permaletailsidescored.
pvalueswerecalculatedusingaStudent'st-test.
*:significantdifferencefromnocDNAcontrol(p<0.
05).
**:highlysignif-icantdifferencefromnocDNAcontrol(p<0.
01).
ThepvaluesforexperimentswithA11cDNAoverexpressionshowednosignificantdifferencefromcontrols(p=0.
59forqc3;p=0.
16forcc604).
Allstrainscontainhim-5(e1490)toincreasetheproportionofmales.
B.
Bodysizephenotypes.
Meanbodylengthat96hoursafterembryocollection(adultstage)isshown.
Errorbarsshowstandarddeviation.
Notethatthesestrainsaretransgenicsexpress-ingtherol-6markerthatinfluencesbodylength,soallcomparisonsaremadebetweenheat-shockedanimalsandnoheatshockcontrolratherthanusinganontransgeniccontrol.
pvalueswerecalculatedusingaStudent'st-test.
**:highlysignificantdifferencefromnoheatshockcontrol(p<0.
01).
Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page9of11scriptsfromalongerprecursor,weexpectedtofindreciprocalchangesinaccumulationofthefull-lengthtranscriptsrelativetoshorttranscripts.
OurresultsarethereforenotsufficientlycleartodrawaconclusionastowhetherMEC-8isdirectlyinvolvedinsma-9trans-splic-ing,andadditionalinvestigationswillbenecessarytoidentifythetrans-actingfactorsthatmediatethisreac-tion.
Avarietyofgeneticmechanismsexistforgeneratingdiversefunctionsfromasinglegeneticlocus,includingthosethataffecttranscriptstructuresuchasalternativesplicing,alternativetranscriptionalstartsites,andalter-nativepolyadenylation.
Oneprominentexampleofalter-nativesplicingistheregulationofsexdeterminationinDrosophilabySex-lethal[40].
Thissystemalsousesdevelopmentallyregulatedalternativepromoters.
InC.
elegans,extensiveuseofalternativepromotersleadingtodifferentproteinisoformshasbeendocumented[41].
Alternativepolyadenylationcanalsoleadtotheproduc-tionoftruncatedproteinvariants,suchasinthedaf-4BMPreceptorgeneinC.
elegans,inwhichalternativepolyadenylationleadstotheproductionofasecretednegativeinhibitorofDAF-4signaling[42].
Toourknowl-edge,alternativetrans-splicingaswehavecharacterizeditforsma-9hasnotpreviouslybeenreportedforotherlociinC.
elegans.
Theuniquefeatureofthistrans-splic-ingeventisthechoicebetweenprocessingthetranscriptviacis-ortrans-splicingatthespliceacceptorsiteofpre-dictedexon11.
Inanorganismwithouttrans-splicing,asimilartruncationcouldbeachievedviauseofadown-streamtranscriptionalstartsiteorviaproteolyticcleav-age.
Forexample,intheHedgehogsignalingpathway,full-lengthtranscriptionfactorsoftheCi/Glifamilyaretranscriptionalactivatorsthat,uponproteolyticcleavage,areconvertedtotranscriptionalrepressors[43].
Wespec-ulatethatsincethetrans-splicingmachineryexistsinC.
elegans,itwasavailabletoberecruitedforthisunusualroleinthesma-9locus.
ConclusionsBothC.
eleganssma-9/schnurri[13]andthehumanhomologsShn-1andShn-3[23,24]displayalternativesplicingthatispredictedtoresultinmultipleproteiniso-forms.
Formostoftheseisoforms,however,nospecificinvivofunctionhasbeendescribed.
Wehavenowdemon-stratedafunctionalroleforanalternativelytrans-splicedsma-9shorttranscriptrepresentedbycDNAcloneyk1237d01(D01).
Thistranscriptmediatesatissue-spe-cificrole,sinceitisrequiredforsensoryraydevelopmentbutnotforbodysizeregulation.
Ourresultsprovideinsightintohowasinglegeneticlocuscancontributetodiverseproteinfunctions.
MethodsC.
elegansstrainsandcultureNematodeswereculturedusingstandardmethodsandgrownat20°Cunlessotherwisenoted[44].
Inadditiontostrainsgeneratedinthiswork,thefollowingstrainswereused:N2(wildtype);LGV,him-5(e1490);LGX,sma-9(qc3,wk55,cc604).
RT-PCRRT-PCRwasperformedintwostepsusingSuperScriptIIIFirst-StrandSynthesisSystem(Invitrogen)andRNAextractedfrommixed-stageN2wild-typeanimals.
Tem-peraturegradientPCRwasperformedonanEppendorfgradientthermalcycler(MasterCycler)followingthemanufacturer'sinstructions.
RealtimeRT-PCRwasperformedonaLightCycler2.
0(Roche).
DataanalysissoftwareusedwasLightCyclersoftwareversion4.
0.
AnimalswerecollectedatdesiredtimepointsandtotalRNAextractedbyTrizolasdescribed[13].
SuperScriptTMIIIplatinumtwo-stepqRT-PCRkitwithSYBRGreen(Invitrogen)wasusedtoperformRT-PCR.
Actingeneact-1wasusedasstandardcontrol.
Aprimerinanintronoftheact-1genewasusedtoconfirmtheabsenceofgenomicDNAinRNApreps.
PrimersequencesforallRT-PCRexperimentsareavail-ableuponrequest.
RNAiRNAifeedingmethodwasperformedasdescribed[29]aftercloningsma-9fragmentsintoRNAifeedingvectorpPD129.
36(agiftfromDr.
A.
Fire).
Toscoresensoryraypatterning,onlyadultmalesdisplayingasmallphenotypewerecollectedandscoredtoensurethattheanimalsana-lyzedhadundergoneaknockdownofsma-9activity.
Heat-shockcloneconstructsandgenerationoftransgenicanimalsTwosma-9splicevariantsyk1285a11andyk1237d01(GenBankAccessionNumbers:AY390537andAY390550)wereidentifiedpreviously[13],whichwereSL1-andSL2-trans-splicedcDNAclonesthatappearedtorepresentshorterbutcompletemRNAs.
Heat-shockvectorpPD49.
83wasakindgiftfromDr.
J.
Liu.
yk1285a11andyk1237d01wereclonedintopPD49.
83atNheIandKpnIsites.
Transgenicnematodesweregener-atedbymicroinjectionofconstructs(10ng/ul)intothegonadalsyncytiaofhim-5hermaphrodites,withrol-6(100ng/ul)asamarker[45].
Mutantstrainscarryingtransgenicconstructsweregeneratedbyappropriatecrossesbetweenthemutantstrainsandtransgeniclines.
Heat-shockexperimentsGravidhermaphroditeswerewashedoffplateswithM9bufferandcollectedinanEppendorftube.
Hermaphro-Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page10of11ditesweredestroyedwithsodiumhypochloritesolution,leavingeggsthatwerecollectedandinoculatedontoOP50-seededplates.
Forbodysizemeasurement,24hoursaftereggcollection,wormswerecollectedandtransferredtoasiliconizedEppendorftubecontaining100ulofM9buffer[44],whichwasplacedinacirculatingwaterbathundertheheat-shockconditionsspecified.
Afterheat-shock,wormswererecovered,placedat25°ConOP50-seededplatesandallowedtodeveloptoadult-hood.
ThelengthofindividualwormswasmeasuredusingImage-ProExpresssoftware(Sigma)withNomar-skioptics.
Formaletailrayexamination,wormswerecol-lected72hoursaftereggcollectionandtransferredtoasiliconizedEppendorftubecontaining100ulofM9buf-fer[44],whichwasplacedinacirculatingwaterbathundertheheat-shockconditionsspecified.
Afterheat-shock,wormswererecovered,placedat25°ConOP50-seededplatesandallowedtodeveloptoadulthoodandthenscoredforraypatterning.
Authors'contributionsJYperformedtheheatshockandRT-PCRexperimentsandparticipatedininterpretationofdata.
LYperformedtheRNAiexperiments.
CSDconceivedofthestudy,participatedinitsdesignandcoordination,analyzedmutantpheno-types,anddraftedthemanuscript.
Allauthorsreadandapprovedthefinalmanuscript.
AcknowledgementsWethankJunKellyLiuforsharingreagentsandforhelpfuldiscussions.
SomeoftheexperimentsweredonewithequipmentfromtheCoreFacilitiesforImag-ing,CellularandMolecularBiologyatQueensCollege.
SomeC.
elegansmutantstrainswereobtainedfromtheCaenorhabditisGeneticsCenter,whichissup-portedbytheNIHNationalCenterforResearchResources(NCRR).
ThisworkwascarriedoutinpartialfulfillmentoftherequirementsforthePh.
D.
degreefromtheGraduateCenterofCityUniversityofNewYork(J.
Y.
).
ThisresearchwassupportedbyRSG-98-230-04-DDCfromtheAmericanCancerSocietytoC.
S.
-D.
AuthorDetailsDepartmentofBiology,QueensCollege,andBiochemistryPhDProgram,theGraduateSchoolandUniversityCenter,theCityUniversityofNewYork,Flushing,NY11367,USAReferences1.
RobertsAB,SpornMB:Physiologicalactionsandclinicalapplicationsoftransforminggrowthfactor-beta(TGF-beta).
GrowthFactors1993,8(1):1-9.
2.
MassagueJ,ChenYG:ControllingTGF-betasignaling.
GenesDev2000,14(6):627-644.
3.
WranaJL,AttisanoL,CarcamoJ,ZentellaA,DoodyJ,LaihoM,WangXF,MassagueJ:TGFbetasignalsthroughaheteromericproteinkinasereceptorcomplex.
Cell1992,71(6):1003-1014.
4.
LiuF,VenturaF,DoodyJ,MassagueJ:HumantypeIIreceptorforbonemorphogenicproteins(BMPs):extensionofthetwo-kinasereceptormodeltotheBMPs.
MolCellBiol1995,15(7):3479-3486.
5.
InmanGJ,NicolasFJ,HillCS:NucleocytoplasmicshuttlingofSmads2,3,and4permitssensingofTGF-betareceptoractivity.
MolCell2002,10(2):283-294.
6.
Macias-SilvaM,AbdollahS,HoodlessPA,PironeR,AttisanoL,WranaJL:MADR2isasubstrateoftheTGFbetareceptoranditsphosphorylationisrequiredfornuclearaccumulationandsignaling.
Cell1996,87(7):1215-1224.
7.
KretzschmarM,LiuF,HataA,DoodyJ,MassagueJ:TheTGF-betafamilymediatorSmad1isphosphorylateddirectlyandactivatedfunctionallybytheBMPreceptorkinase.
GenesDev1997,11(8):984-995.
8.
WuRY,ZhangY,FengXH,DerynckR:HeteromericandhomomericinteractionscorrelatewithsignalingactivityandfunctionalcooperativityofSmad3andSmad4/DPC4.
MolCellBiol1997,17(5):2521-2528.
9.
ZhangY,FengX,WeR,DerynckR:Receptor-associatedMadhomologuessynergizeaseffectorsoftheTGF-betaresponse.
Nature1996,383(6596):168-172.
10.
InmanGJ,HillCS:Stoichiometryofactivesmad-transcriptionfactorcomplexesonDNA.
JBiolChem2002,277(52):51008-51016.
11.
LagnaG,HataA,Hemmati-BrivanlouA,MassagueJ:PartnershipbetweenDPC4andSMADproteinsinTGF-betasignallingpathways.
Nature1996,383(6603):832-836.
12.
Savage-DunnC:TGF-βsignaling.
WormBook2005,9:1-12.
13.
LiangJ,LintsR,FoehrML,TokarzR,YuL,EmmonsSW,LiuJ,Savage-DunnC:TheCaenorhabditiselegansschnurrihomologsma-9mediatesstage-andcelltype-specificresponsestoDBL-1BMP-relatedsignaling.
Development2003,130(26):6453-6464.
14.
FoehrML,LindyAS,FairbankRC,AminNM,XuM,YanowitzJ,FireAZ,LiuJ:AnantagonisticrolefortheC.
elegansSchnurrihomologSMA-9inmodulatingTGFbetasignalingduringmesodermalpatterning.
Development2006,133(15):2887-2896.
15.
DaiH,HoganC,GopalakrishnanB,Torres-VazquezJ,NguyenM,ParkS,RafteryLA,WarriorR,AroraK:ThezincfingerproteinschnurriactsasaSmadpartnerinmediatingthetranscriptionalresponsetodecapentaplegic.
DevBiol2000,227(2):373-387.
16.
MartyT,MullerB,BaslerK,AffolterM:SchnurrimediatesDpp-dependentrepressionofbrinkertranscription.
NatCellBiol2000,2(10):745-749.
17.
TakagiT,HaradaJ,IshiiS:MurineSchnurri-2isrequiredforpositiveselectionofthymocytes.
NatImmunol2001,2(11):1048-1053.
18.
OukkaM,KimST,LugoG,SunJ,WuLC,GlimcherLH:AmammalianhomologofDrosophilaschnurri,KRC,regulatesTNFreceptor-drivenresponsesandinteractswithTRAF2.
MolCell2002,9(1):121-131.
19.
WuLC:ZAS:C2H2zincfingerproteinsinvolvedingrowthanddevelopment.
GeneExpr2002,10(4):137-152.
20.
JinW,TakagiT,KanesashiSN,KurahashiT,NomuraT,HaradaJ,IshiiS:Schnurri-2controlsBMP-dependentadipogenesisviainteractionwithSmadproteins.
DevCell2006,10(4):461-471.
21.
YaoLC,BlitzIL,PeifferDA,PhinS,WangY,OgataS,ChoKW,AroraK,WarriorR:SchnurritranscriptionfactorsfromDrosophilaandvertebratescanmediateBmpsignalingthroughaphylogeneticallyconservedmechanism.
Development2006,133(20):4025-4034.
22.
ShuklaA,MalikM,CataissonC,HoY,FriesenT,SuhKS,YuspaSH:TGF-betasignallingisregulatedbySchnurri-2-dependentnucleartranslocationofCLIC4andconsequentstabilizationofphospho-Smad2and3.
NatCellBiol2009,11(6):777-784.
23.
MuchardtC,SeelerJS,NirulaA,ShurlandDL,GaynorRB:RegulationofhumanimmunodeficiencyvirusenhancerfunctionbyPRDII-BF1andc-relgeneproducts.
JVirol1992,66(1):244-250.
24.
HicarMD,LiuY,AllenCE,WuLC:StructureofthehumanzincfingerproteinHIVEP3:molecularcloning,expression,exon-intronstructure,andcomparisonwithparalogousgenesHIVEP1andHIVEP2.
Genomics2001,71(1):89-100.
25.
BlumenthalT:Trans-splicingandoperons.
WormBook2005,25:1-9.
26.
HuangT,KuerstenS,DeshpandeAM,SpiethJ,MacMorrisM,BlumenthalT:Intercistronicregionrequiredforpolycistronicpre-mRNAprocessinginCaenorhabditiselegans.
MolCellBiol2001,21(4):1111-1120.
27.
BlumenthalT,EvansD,LinkCD,GuffantiA,LawsonD,Thierry-MiegJ,Thierry-MiegD,ChiuWL,DukeK,KiralyM,etal.
:AglobalanalysisofCaenorhabditiselegansoperons.
Nature2002,417(6891):851-854.
28.
LameschP,MilsteinS,HaoT,RosenbergJ,LiN,SequerraR,BosakS,Doucette-StammL,VandenhauteJ,HillDE,etal.
:C.
elegansORFeomeversion3.
1:increasingthecoverageofORFeomeresourceswithimprovedgenepredictions.
GenomeRes2004,14(10B):2064-2069.
29.
KamathRS,AhringerJ:Genome-wideRNAiscreeninginCaenorhabditiselegans.
Methods2003,30(4):313-321.
30.
SijenT,FleenorJ,SimmerF,ThijssenKL,ParrishS,TimmonsL,PlasterkRH,FireA:OntheroleofRNAamplificationindsRNA-triggeredgenesilencing.
Cell2001,107(4):465-476.
Received:3March2010Accepted:17June2010Published:17June2010Thisarticleisavailablefrom:http://www.
biomedcentral.
com/1471-2199/11/462010Yinetal;licenseeBioMedCentralLtd.
ThisisanOpenAccessarticledistributedunderthetermsoftheCreativeCommonsAttributionLicense(http://creativecommons.
org/licenses/by/2.
0),whichpermitsunrestricteduse,distribution,andreproductioninanymedium,providedtheoriginalworkisproperlycited.
BMCMolecularBiology2010,11:46Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page11of1131.
BosherJM,DufourcqP,SookhareeaS,LabouesseM:RNAinterferencecantargetpre-mRNA:consequencesforgeneexpressioninaCaenorhabditiselegansoperon.
Genetics1999,153(3):1245-1256.
32.
MangoSE:StopmakingnonSense:theC.
eleganssmggenes.
TrendsGenet2001,17(11):646-653.
33.
LiangJ,YuL,YinJ,Savage-DunnC:TranscriptionalrepressorandactivatoractivitiesofSMA-9contributedifferentiallytoBMP-relatedsignalingoutputs.
DevBiol2007,305(2):714-725.
34.
LintsR,EmmonsSW:PatterningofdopaminergicneurotransmitteridentityamongCaenorhabditiselegansraysensoryneuronsbyaTGFbetafamilysignalingpathwayandaHoxgene.
Development1999,126(24):5819-5831.
35.
MullerB,HartmannB,PyrowolakisG,AffolterM,BaslerK:ConversionofanextracellularDpp/BMPmorphogengradientintoaninversetranscriptionalgradient.
Cell2003,113(2):221-233.
36.
Torres-VazquezJ,ParkS,WarriorR,AroraK:ThetranscriptionfactorSchnurriplaysadualroleinmediatingDppsignalingduringembryogenesis.
Development2001,128(9):1657-1670.
37.
PyrowolakisG,HartmannB,MullerB,BaslerK,AffolterM:AsimplemolecularcomplexmediateswidespreadBMP-inducedrepressionduringDrosophiladevelopment.
DevCell2004,7(2):229-240.
38.
SpikeCA,DaviesAG,ShawJE,HermanRK:MEC-8regulatesalternativesplicingofunc-52transcriptsinC.
eleganshypodermalcells.
Development2002,129(21):4999-5008.
39.
LundquistEA,HermanRK,RogalskiTM,MullenGP,MoermanDG,ShawJE:Themec-8geneofC.
elegansencodesaproteinwithtwoRNArecognitionmotifsandregulatesalternativesplicingofunc-52transcripts.
Development1996,122(5):1601-1610.
40.
BlackDL:Mechanismsofalternativepre-messengerRNAsplicing.
AnnuRevBiochem2003,72:291-336.
41.
ChoiJ,NewmanAP:Atwo-promotersystemofgeneexpressioninC.
elegans.
DevBiol2006,296(2):537-544.
42.
GuntherCV,RiddleDL:Alternativepolyadenylationresultsinatruncateddaf-4BMPreceptorthatantagonizesDAF-7-mediateddevelopmentinCaenorhabditiselegans.
JBiolChem2004,279(38):39555-39564.
43.
HuangfuD,AndersonKV:SignalingfromSmotoCi/Gli:conservationanddivergenceofHedgehogpathwaysfromDrosophilatovertebrates.
Development2006,133(1):3-14.
44.
BrennerS:ThegeneticsofCaenorhabditiselegans.
Genetics1974,77(1):71-94.
45.
MelloCC,KramerJM,StinchcombD,AmbrosV:EfficientgenetransferinC.
elegans:extrachromosomalmaintenanceandintegrationoftransformingsequences.
EmboJ1991,10(12):3959-3970.
doi:10.
1186/1471-2199-11-46Citethisarticleas:Yinetal.
,Alternativetrans-splicingofCaenorhabditisele-ganssma-9/schnurrigeneratesashorttranscriptthatprovidestissue-specificfunctioninBMPsignalingBMCMolecularBiology2010,11:46
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46OpenAccessRESEARCHARTICLE2010Yinetal;licenseeBioMedCentralLtd.
ThisisanOpenAccessarticledistributedunderthetermsoftheCreativeCommonsAt-tributionLicense(http://creativecommons.
org/licenses/by/2.
0),whichpermitsunrestricteduse,distribution,andreproductioninanymedium,providedtheoriginalworkisproperlycited.
ResearcharticleAlternativetrans-splicingofCaenorhabditiseleganssma-9/schnurrigeneratesashorttranscriptthatprovidestissue-specificfunctioninBMPsignalingJianghuaYin,LingYuandCathySavage-Dunn*AbstractBackground:TranscriptioncofactorsrelatedtoDrosophilaSchnurrifacilitatethetranscriptionalprogramsregulatedbyBMPsignalinginC.
elegans,Drosophila,Xenopus,andmouse.
Indifferentsystems,SchnurrihomologshavebeenshowntoactaseitheragonistsorantagonistsofSmadfunction,andaseitherpositiveornegativeregulatorsoftranscription.
HowSchnurriproteinsachievethisdiversityofactivitiesisnotclear.
TheC.
eleganssma-9/schnurrilocusundergoesalternativesplicing,includinganunusualtrans-splicingeventthatcouldgeneratetwonon-overlappingshortertranscripts.
Results:Wedemonstrateherethattheshortertranscriptsareexpressedinvivo.
Furthermore,wefindthatoneoftheshorttranscriptsplaysatissue-specificroleinsma-9function,contributingtothepatterningofmale-specificsensoryrays,butnottotheregulationofbodysize.
Basedonpreviousresults,wesuggestthatthistranscriptencodesaC-terminalSMA-9isoformthatmayprovidetranscriptionalactivationactivity,whilefulllengthisoformsmaymediatetranscriptionalrepressionand/oractivationinacontext-dependentmanner.
Conclusion:Thealternativetrans-splicingofsma-9maycontributetothediversityoffunctionsnecessarytomediatetissue-specificoutputsofBMPsignaling.
BackgroundThetransforminggrowthfactorβ(TGFβ)superfamilycomprisesalargenumberofsecretedpeptidegrowthfac-torsthathavemajorregulatoryeffectsoncellgrowthanddifferentiation[1,2].
MembersofthissuperfamilyincludetheTGFβs,theprototypesofthesuperfamily;thebonemorphogeneticproteins(BMPs);andothermemberssuchasactivin,inhibin,andNodal.
TGFβsuperfamilyligandsbindtoaheteromericreceptorcomplexatthecellsurface.
Thiscomplexcontainstworelatedtransmem-braneserine/threoninekinases,thetypeIandtypeIIreceptors[3,4].
SignalingdownstreamofthereceptorsismediatedbytheSmadproteins,whichshuttlebetweenthecytoplasmandnucleustoregulatetargetgeneexpres-sion[5].
TypeIreceptorsdirectlyactivatereceptor-regu-latedSmads(R-Smads)byphosphorylationatC-terminalSXSsequences[6,7].
PhosphorylationofR-Smadspro-motesheterotrimericcomplexformationwithCo-Smadsandaccumulationinthenucleustoregulategenetran-scription[7-11].
Inmammals,fiveR-Smadsarepresent:two(Smad2,3)thattransduceTGFβ/activin/Nodalsig-nalsandthree(Smad1,5,8)thattransduceBMPsignals.
Strikingly,theTGFβfamilyligandsfaroutnumbertheSmadsavailableforsignaltransduction.
Furthermore,manyoftheseligandsarecapableofelicitingdiversecon-text-dependentresponses.
Thus,Smadcomplexesmustbecapableofmediatingmultiplediverseoutcomes.
ItisthoughtthatSmadcomplexesrelyinpartontranscrip-tioncofactorsforappropriateregulationoftargetgenes.
InthenematodeCaenorhabditiselegans,theBMP-relatedfactorDBL-1regulatesbodysizeandmaletailmorphogenesisviaaconservedreceptor/Smadsignalingpathway[12].
Usingageneticapproachtouncovercom-ponentsofthispathway,wepreviouslyidentifiedsma-9,agenethatisrequiredforthebodysizeandmaletailpat-terningfunctionsofDBL-1[13].
sma-9isalsorequiredforpatterningofthemesodermallineage,inwhichitacts*Correspondence:cathy.
savagedunn@qc.
cuny.
edu1DepartmentofBiology,QueensCollege,andBiochemistryPhDProgram,theGraduateSchoolandUniversityCenter,theCityUniversityofNewYork,Flushing,NY11367,USAFulllistofauthorinformationisavailableattheendofthearticleYinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page2of11antagonisticallytotheDBL-1pathway[14].
sma-9ispre-dictedtoencodemultipleproteinproductshomologoustoDrosophilaSchnurri,alargezincfingertranscriptioncofactorthatfunctionsinDpp/BMPsignaling[15,16].
Analysisofsma-9thereforeprovidestheopportunitytoelucidatetherequirementsfortranscriptioncofactorfunctioninaninvivomodelsystemduringthecourseofdevelopment.
InadditiontoDrosophilaSchnurriandC.
elegansSMA-9/Schnurri,threevertebrateSchnurrihomologshavebeenidentified.
ThesebindtheκB-bind-ingsiteandfunctioninTcelldevelopment[17-19].
Nota-bly,vertebrateSchnurrihomologshavemorerecentlybeendemonstratedtomediatetranscriptionalregulationdownstreamofBMPandTGFβligands,indicatingacon-servedroleforthesefamilymembersinTGFβsignaltransduction[20-22].
Thesma-9openreadingframe(ORF)predictedfromgenomicsequenceencodesaproteinof2170aaconsist-ingofanN-terminalGln-richdomain(encodedbypre-dictedexons1-3)andaC-terminaldomaincontainingsevenZnfingers(inpredictedexons12-22).
ThesequencingofcDNAclones,however,revealedatleasteightdifferentmRNAspecieswithalternativeproteincodingregions[13].
Interestingly,likesma-9,humanShn-1andShn-3genesundergoalternativesplicing[23,24],butthefunctionalconsequencesofthisprocess-inghavenotbeenaddressed.
Thesma-9cDNAcloneswereclassifiedbasedontheirpotentialtocodeforthesevenC-terminalZnfingers:classIencodesallsevenZnfingers(ZF1-7),classIIencodesthefirstpairandthetrip-letofZnfingers(ZF1-5),andclassIIIencodesonlythefirstpair(ZF1-2).
Todeterminewhetherthesedomainshavedifferentfunctionsinvivo,Foehretal.
createdgenomic/cDNAhybridconstructsandtestedtheirabilitytorescuethebodysizeandmesodermalpatterningdefectsofsma-9mutants[14].
Theyfoundthatcon-structsencodingclassIandclassIIC-terminiwerecapa-bleofrescuingbothphenotypes,whiletheclassIIIconstructwasnot,suggestingthatthepresenceoftheZnfingertripletiscriticalforfunctioninbodysizeandmesodermallineageregulation.
Sincesma-9isalargegene,mostoftheexistingcDNAclonescontainincompletecodingsequencesthataremissingthe5'endofthegene.
TwocDNAclones,yk1285a11andyk1237d01,however,arenotablydifferent(Figure1).
AlthoughthesetwocDNAclonesarelessthanhalfthelengthofthepredictedtranscript,bothofthemhaveallofthehallmarksofafull-lengthcDNA:trans-splicedleadersequence,poly(A)tail,andacompleteORF.
InC.
elegans,manytranscriptsareprocessedatthe5'endbytrans-splicing,whichresultsintheadditionofa22-nucleotidespliceleadersequence,SL1orSL2[25].
Themechanismoftrans-splicingissimilartothatofcis-splic-ing(intronremoval),exceptthatthesplicedonorsequencesareprovidedbytheSL1andSL2genes,ratherthanbeingcontainedwithinthecontextoftheindividualgene[25].
AbouthalfofallC.
elegansgenesaresubjectedtoSL1trans-splicingatthe5'end.
AsmallersubsetofC.
elegansgenesaretranscribedinpolycistronicoperons.
Inanoperon,the5'-mostgenegenerallyreceivestheSL1spliceleader.
TheindividualdownstreamgenesinanoperonarethenseparatedviaSL2trans-splicingtothe5'endsofeachofthedownstreamgenes.
SL2trans-splicingisaccompaniedbypolyadenylationtocreatethe3'endoftheneighboringgeneupstream.
yk1285a11containstheSL1spliceleaderandsequencesfrompredictedexons1-7.
yk1237d01containstheSL2spliceleaderandsequencesfrompredictedexons11-25.
Aspreviouslynoted[13],theputativeintercistronicregioncontainsfeaturesreminiscentoftheidentifiedsignalsfortrans-splicing,includingaU-richsequence,butisatypicallylong[26,27].
Thus,thestructuresofthesma-9cDNAclonesyk1285a11andyk1237d01suggestthattheymayrepresenttwotranscriptsprocessedfromasinglelongertranscriptbySL2trans-splicingandpolyadenylationasnormallyoccursinaC.
eleganspolycistronicoperon.
Sinceeachofthesetrans-splicedshortsma-9tran-scriptswasonlyrepresentedbyasinglecDNAclone,wecouldnotbecertainthatthecloneswerenotduetoclon-ingartifactsorararespuriousevent.
Furthermore,ourpreviousanalysisdidnotaddresswhethertheseshorttranscriptsprovideanyfunctionrequiredforDBL-1sig-naltransduction.
Wethereforeaddresshereseveralremainingquestionsaboutthesepredictedtranscripts.
First,canweverifythatthemRNAvariantsrepresentedbyyk1285a11andyk1237d01areexpressedinvivo,ratherthanbeingartifactsproducedduringcDNAlibrarycon-structionIfso,thenhowistheexpressionofthesevari-antsregulatedFinally,dothesevariantsshowfunctionaldifferencesbetweeneachotherand/orrelativetofull-lengthtranscriptsinvivoInthiswork,wewillrefertothesplicevariantsrepresentedbyclonesyk1285a11(5')andyk1237d01(3')asA11andD01,respectively.
ResultsIsolationofcDNAsspanningupstreamanddownstreamregionsThesma-9ORFpredictedfromgenomicsequenceisencodedby25exons([13];Figure1).
Previouslycharac-terizedcDNAclones,however,containedonlyasubsetofthese25exonsand,inparticular,noneoftheexistingcDNAclonesspannedaregionincludingboththeN-ter-minalGln-richdomainandtheC-terminalZnfingerdomains.
Wethereforeusedprimersinexons1and25withthepotentialtogeneratenearlyfull-lengthcDNAinsertsbyRT-PCR.
AfterRT-PCR,thecloneswiththelongestinsertswereselectedforsequencing.
ThemostcompletecDNAcloneisolatedbythisapproachwasYinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page3of11pCS368,inwhichexons4-8andpartofexon9aresplicedout(Figure1).
ThistranscriptformisreminiscentofaformidentifiedbytheORFeomeproject,inwhichexon3becomessplicedtoexon20[28].
Theseresultsthussup-porttheexistenceofsma-9transcriptscapableofencod-ingboththeGln-richandtheZnfingerdomains.
AdditionalsupportderivesfromRT-PCRdatausingexon9primers(seebelow).
Expressionofsma-9varianttranscriptsWenextsoughttoobtainevidencethattheshorttran-scriptsrepresentedbycDNAclonesyk1285a11andyk1237d01(A11andD01)areexpressedinvivo,ratherthancDNAartifacts.
Toaddressthisquestion,weneededanapproachtodistinguishtheseshorttranscriptsfromfull-lengthtranscriptscontainingthesameinternalsequences.
Unfortunately,wewereunabletodetectsma-9mRNAsbyNorthernblot(datanotshown).
Instead,weusedtheuniqueterminioftheshorttranscriptstodesignvariant-specificprimerpairsforRT-PCR.
ForA11weemployedanoligo-dT-containingreverseprimeranchoredbytwonucleotidesfromsma-9exon7withagene-specificinternalforwardprimer;forD01weusedanSL2-containingprimeranchoredbytwonucleotidesfromsma-9exon11withagene-specificinternalreverseprimer.
TodetermineempiricallywhethertheseprimerpairsarespecificforA11andD01,weperformedRT-PCRusingagradientofannealingtemperatures(50°C-55°C)andanalyzedtheproducts(Figure2A).
Inbothcases,theprimerpairsamplifiedasinglebandofthecor-rectsize(330bpforA11and300bpforD01),indicatingthattheprimersspecificallydetectthedesiredtran-scripts.
Sincethehigherannealingtemperaturesresultedinreducedyield,weused50°Cforallsubsequentanaly-ses.
Basedontheseresults,weconcludethattranscriptspolyadenylateddownstreamofexon7andtranscriptscontainingtheSL2spliceleadersequenceupstreamofexon11arerepresentedinthepoolofsma-9mRNAsinvivo.
TheinitialRT-PCRexperimentswereperformednon-quantitativelyonmixed-stageRNApreparations.
Wesubsequentlyusedreal-timequantitativeRT-PCR(qRT-PCR)todeterminetherelativeexpressionlevelsofsma-9varianttranscriptsandwhethertheyaredevelopmentallyorsex-specificallyregulated.
Sinceweexpectfull-lengthtranscripts,butnottheshorttranscripts,tocontainsequencesfromexon9,weusedexon9internalprimerstodeterminetheexpressionlevelsofpotentiallyfull-lengthtranscriptsandcalculatedtheabundanceoftheshorttranscriptsrelativetoexon-9-containingtran-scripts.
Threedevelopmentaltimepointswereassayed:24hr(firstlarvalstage-L1),44hr(approximatelyL3),and96hr(adults).
Wefindthatexon-9-containingtran-scriptlevelsincreaseduringlarvaldevelopmentandarehighestinadulthood(Figure2B).
D01transcriptsaccu-mulateinwild-typestrainN2at45%-85%ofthelevelofexon-9-containingtranscriptsatalldevelopmentaltimepointsexamined,whileA11transcriptsaccumulateatapproximately10%ofthelevelofexon-9-containingtran-scripts(Figure2B;Table1).
IfA11andD01areproducedconcurrentlybytrans-splicingoffull-lengthtranscripts,thenthereducedaccumulationofA11maybeduetodif-Figure1Structureofsma-9transcriptvariants.
Thepredictedsma-9intron-exonstructureisshownabove.
Arrowsmarkpositionsofnonsensecodonsinthreesma-9alleles.
pCS386isthemostcompletecDNAvariantisolatedtodate.
ThetwoshortvariantsrepresentedbycDNAclonesyk1285a11(A11)andyk1237d01(D01)areshownbelow.
ARD:acidicresidue-richdomain;NLS:nuclearlocalizationsignal.
pCS368pCS368Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page4of11Figure2Expressionofsma-9transcriptvariants.
A.
DetectionofA11andD01transcriptsusingvariant-specificRT-PCRoveragradientofannealingtemperatures(lefttoright:50°C-55°C).
TheexpectedsizeofthePCRproductis330bpforA11and300bpforD01.
B.
Developmentalprofileofsma-9transcriptexpressionlevels,asdeterminedbyqRT-PCR.
Dataareshownastranscriptabundancerelativetoact-1actingenecontrol,anderrorbarsshowstandarddeviation.
SeeTable1forquantitationoftranscriptabundancerelativetoexon-9-containingcontrol.
ABA11D010501001502002503003502444962444HoursAfterEmbryoCollectionTranscriptLevel(relativetoactincontrol)A11exon9D01N2him-5ferentialstabilityofthetranscripts.
Onepossiblemecha-nismforreducedstabilityoftheA11transcriptistheunusuallocationoftheterminationcodonTAA,inwhichtheterminaladeninesareintroducedduringpolyadenyla-tionandarecontiguouswiththepoly(A)tail.
Totestforsex-specificexpressionofthesetranscripts,him-5(highincidenceofmales)mutantpopulationsthatcontainbothhermaphroditesandmaleswerecomparedtowild-typeN2hermaphroditepopulations.
Inhim-5populations,thefull-lengthandtheD01transcripts,butnotA11,accumulatetoa2-4-foldhigherlevelthaninhermaphro-ditepopulations,suggestingthatthesetranscriptsareexpressedatahigherlevelinmales.
Theshorttranscriptsarenotmale-specific,however,sincetheyarealsoexpressedinhermaphrodites.
Thus,theA11andD01cDNAclonesrepresentdetectablemRNAsthatareexpressedduringlarvalandadultstageswhenDBL-1isactiveinregulatingbodysizeandmaletailpatterning.
Lossoffunctionofsma-9varianttranscriptsPreviousworkhasshownthatsma-9constructslackingtheZnfingertriplet(ZF3-5)areunabletorescuethebodysizeandmesodermalpatterningdefectsofasma-9mutant[14].
Theseexperimentsdidnottest,however,fordifferentialrequirementsofshortvs.
full-lengthtran-scripts.
Totestwhethersma-9varianttranscriptshavespecificfunctionalroles,weperformedRNAitargetingthreedifferentregionsofthesma-9gene.
Inpreviousexperiments,dsRNAwasintroducedbymicroinjection[13,14],butwehavefoundthatRNAibyfeedingresultsinahigherpenetranceofmaletaildefects.
Wehavethere-forerepeatedtheseexperimentsusingthefeedingtech-nique[29],andextendedthembytargetingtheinternalexon9inadditiontothe5'and3'endsofthegene.
TheeffectivenessofeachRNAitreatmentwasmonitoredbytheappearanceofthecharacteristicsmallbodysizephe-notype(datanotshown).
AsacomponentintheDBL-1pathway,sma-9mutantphenotypesincludesmallbodysizeandmaleabnormalphenotypes[13].
Oneaspectofthemaleabnormalphe-notypeinsma-9isthefusionofmaletailsensoryrays8and9,twooftheninebilaterallysymmetricalpairsofmale-specificsensoryorgans.
Inwild-typeanimals,ray8-9fusionsarealsoobserved,butatalowerfrequency.
Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page5of11Treatmentofwild-typeanimalswithRNAitargetingexons1-7causednoincreaseinthefrequencyofray8-9fusions(Table2),althoughthesmallbodysizephenotypewasproduced.
Thistreatmentispredictedtoknockdownexpressionoffull-lengthandA11variantsbutnottheD01variant.
Conversely,RNAitargetingexons21-25,predictedtoknockdowntheD01andfull-lengthvariants,resultedinatwo-foldincreaseinthefrequencyofray8-9fusions(Table2).
TheseresultsareconsistentwithourpreviousfindingsthatRNAiofthe3'endofthegeneresultsinmoreseveremaleabnormalphenotypesthaninactivationofthe5'end[13].
WenextcomparedthemaletailphenotypeproducedbyRNAitargetingthecen-tralexon9,predictedtoinhibitfull-lengthtranscriptsbutnottheshorttranscripts.
ThistreatmentresultedinamaletaildefectoflowerpenetrancethanthatproducedbyRNAitargetingthe3'end(Table2).
SincethemostseveremaletaildefectoccurredwhenD01wastargeted,theseexperimentssuggestthatD01hasaroleinregulat-ingsensoryraypatterning.
Full-lengthsma-9productsarealsopredictedtofunctioninsensoryraypatterning,sincetheinhibitionofacentralexonresultedinapartialmaleabnormalphenotype.
Wesoughttorepeatthisanalysisusinggeneticmutants,sincetheRNAitreatmentsmightcausesomenonspecificeffectsduetotargetingofunsplicedpre-mRNAortoRNAispreading[30,31].
Wetookadvantageofthreesma-9allelescontainingprematureterminationcodonsatdifferentlocationsinthegene(Figure1;[13,14]):qc3inpredictedexon1(withinA11),wk55inpredictedexon9(betweentheshortvariants),andcc604inpredictedexon18(withinD01).
Therefore,inqc3,onlytheD01transcriptiscapableofencodingafunctionalgeneproduct;inwk55,bothA11andD01,butnotthefull-lengthtranscript,arecapableofencodingfunctionalgeneproducts;andincc604,onlytheA11transcriptiscapableofencodingafunctionalgeneproduct.
Sinceadditionaldisruptionsmayoccurduetononsense-medi-ateddecay[32],wedeterminedthetranscriptexpressionprofileforthesestrainsattheL3stage(Figure3;Table1).
Inallthreemutantbackgrounds,thetranscriptlevelsaresomewhatreducedcomparedtothehim-5control.
Thereductionintranscriptlevelsismoststrikinginthesma-9(wk55)background(Figure3).
Thisseverereductionintranscriptlevelsisconsistentwithourobservationsthatthisallelecausesthemostseverebodysizephenotype(Table2;[13]).
Mostimportantlyfortheinterpretationofourexperiments,sma-9(qc3)andsma-9(cc604)animalsexpresstheshorttranscriptsatdetectablelevelsnearthenormalrange(Figure3).
Theusualexpectationforaseriesofnonsensemuta-tionsisthattheearliestprematureterminationcodonwillhavethemostseveremutantphenotypewhilelaterter-minationcodonsmaycauselessseveredefectsifthoseallelesencodepartiallyfunctionalgeneproducts.
Forthesma-9alleles,theoppositeresultisseeninthemaletail.
Thealleleencodingthelatestterminationcodon,cc604,causesthemostseveremaletaildefect,withafrequencyofray8-9fusionsgreaterthananyofthepreviouslychar-acterizedalleles(Table2,[13]).
Sincedifferentinvestiga-torstypicallyvaryintheirquantitationofrayfusionfrequencies,wealsorepeatedmeasurementofthemaleTable1:Relativeexpressionlevelsofsma-9transcriptvariantsStrainDevelopmentalTimepointA11TranscriptLevelD01TranscriptLevelN2L110.
12±1.
0686.
35±1.
17N2L38.
88±2.
5845.
00±0.
87N2adult8.
97±0.
4384.
58±7.
82him-5L18.
23±0.
2887.
01±0.
37him-5L32.
18±0.
0037.
73±0.
11sma-9(qc3);him-5L36.
20±0.
0149.
55±1.
29sma-9(wk55);him-5L31.
81±0.
0988.
50±1.
32sma-9(cc604);him-5L39.
40±0.
0562.
70±1.
26Meanexpressionlevelsaregiveaspercentoftheexpressionlevelofexon-9-containingcontrol,plusorminusstandarddeviation.
Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page6of11taildefectsofqc3andwk55fordirectcomparison.
Con-sistentwithpreviousresults,qc3,theallelecontainingtheearliestterminationcodon,causesamildmaletaildefect,whilewk55,containingaterminationcodoninexon9,resultsinanintermediatefrequencyofrayfusions(Table2).
Thus,theseverityofthesensoryraydefectisinverselycorrelatedwithpredictedD01activity.
Themostseveredefectismanifestedincc604mutants,inwhichtheD01isoformisdisruptedbymutation.
Anintermediatefre-quencyofrayfusionsoccursinwk55mutants,inwhichtheD01transcriptisexpressedatreducedlevelsbutisnotdisruptedbymutation.
Amildmaletaildefectisseeninqc3mutants,inwhichlevelsofD01transcriptsarelowerthaninwildtypebuthigherthaninwk55(Figure3).
Overall,withregardtomaletailpatterning,thesemutationsformanallelicseriesinwhichtheirlevelsofactivityaretheoppositeofthatexpected.
Inbodysize,adifferentresultwasobtainedwiththesethreemutants(Table2).
Thesma-9(wk55)mutantshowsthesmallestbodysize.
Incontrast,bothqc3andcc604producelessseverebodylengthphenotypesthataremutuallyindistinguishable(Table2).
Sinceqc3andcc604produceindistinguishablebodysizedefects,thisanalysisdoesnotsupportaspecificroleforeithertheA11ortheD01transcriptintheregulationofbodysize.
Further-more,themoreseverebodysizephenotypeofwk55mutantscouldbeduetotheloweroveralllevelofsma-9expressioninthesemutants(Figure3).
Theresultsofbodysizeanalysisthussuggestthatfull-lengthisoforms,ratherthanA11orD01,arecriticalforpromotinggrowth.
BasedonourRNAidataandonthedifferentialeffectsofthethreenonsenseallelesonbodysizeandmaletaildevelopment,wehypothesizethattheD01shorttran-scripthasatissue-specificfunctioninsensoryraydevel-opment.
Rescueofsma-9mutantphenotypesbyoverexpressionofD01cDNAConsistentwithourhypothesis,wehavepreviouslydem-onstratedthatoverexpressionoftheD01cDNA,butnotofA11,fromaheatshockpromotercanpartiallyrescuesma-9(wk55)mutantmaletaildefects[33].
Incompari-son,expressionofsma-9fromacosmidgenomicclonecontainingtheentirecodingregionresultedinnearlycompleterescueoftherayfusiondefect[13].
Wenextaskedwhetherexpressionofsma-9shorttranscriptsissufficienttoprovidenormalgenefunctioninqc3andincc604mutants,inwhichtheA11andtheD01transcript,respectively,aredisruptedbyprematureterminationcodons.
Asbefore,transgenicscarryinghs-sma-9con-structsweresubjectedtoheatshockduringtheL3stagewhensensoryrayidentitiesarebeingestablished[34]andmaleswerescoredinadulthood.
Incc604mutants,whichcontainaprematureterminationcodonwithintheD01transcript,expressionofD01resultedinpartialrescueofthemaletaildefect(Figure4A).
Incontrast,overexpres-sionofA11incc604causedaslightreductioninfre-quencyofsensoryrayfusions,butthiseffectwasnotstatisticallysignificant(p=0.
16).
Inqc3mutants,inwhichtheA11transcriptcontainsaprematuretermina-tioncodon,weweresurprisedtofindthatoverexpressionofA11producednochangeinmaletailphenotype,whereasoverexpressionofD01resultedinsignificantres-cueofthemaletaildefect(Figure4A).
Basedontheres-Table2:sma-9lossoffunctionphenotypesinmaletailpatterningandbodysizeRNAi(Exon)orMutantFormsPredictedtobeBlockedFrequencyofRay8-9Fusions1(n)BodyLength2(n)nonenone18%(50)ND1-7full-length,A1120%(50)ND9full-lengthonly29%(100)ND21-25full-length,D0136%(50)NDsma-9(qc3)full-length,A1150%(50)692±85(43)sma-9(wk55)full-lengthonly66%(50)566±34(31)sma-9(cc604)full-length,D0172%(50)716±61(30)1Allmaletailphenotypesarescoredwithhim-5(e1490)inthebackgroundtoincreasethefrequencyofmales.
2Bodylengthisgiveninμm±standarddeviationat96hoursafterembryocollection.
ND:notdetermined.
Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page7of11cueofmaletaildefectsbyD01inbothqc3andcc604mutants,weconcludethatincreasedexpressionofD01maybeabletocompensatepartiallyfordefectivefull-lengthsma-9transcripts.
Finally,welookedforevidenceofA11orD01functioninbodysizeregulation.
Thebodysizephenotypesofqc3andcc604didnotsupportaspecificroleforeitherA11orD01inbodysizeregulation,sincethesetwomutantshaveindistinguishablesizes(Table2).
TodeterminewhetherincreasedexpressionofA11and/orD01canpromotegrowth,weusedourheatshockconstructstodrivetheirexpression.
HeatshockoftransgenicsintheL3stagepro-ducednoevidentchangesinbodysize(datanotshown),soweperformedheatshockearlierindevelopment,inL1animals,andmeasuredbodylengthinadulthood.
Incon-trasttotherescueofmaleabnormalphenotypesbyD01,heatshockofA11andD01transgenicsledeithertoareductioninbodysizeortonochangeinbodysize(Fig-ure4B).
TheseresultssuggestthatA11andD01donotcontributetothegrowthpromotingactivityofsma-9.
Ifanything,increasedexpressionofA11andD01mayinterferewiththeabilityofgeneproductsfromfull-lengthtranscriptstopromoteincreasesinbodysize.
DiscussionWehaveshownherethattheC.
eleganssma-9/schnurrilocusundergoesanoveltrans-splicingprocesstogener-atetwoshortertranscripts,oneofwhichhasatissue-spe-cificfunctioninDBL-1/BMPsignaling.
First,wehavedemonstratedthatanSL2trans-spliced,truncatedcDNAform,yk1237d01(D01),representsanactualmRNAthatisdetectablebyRT-PCR.
Second,wehaveverifiedthatlongertranscriptscontainingboththe5'and3'endsofthegenearealsodetectablebyRT-PCR.
Third,loss-of-functionexperiments(RNAiandprematureterminationmutants)demonstratethattheshorttranscriptD01isnecessarybutnotsufficientformaletailpatterning,sincedepletionofbothD01andfull-lengthtranscriptscausesamoreseveremaletailpatterningdefectthandepletionoffull-lengthtranscriptsalone.
Fourth,gain-of-functionexperiments(heatshock-inducedoverexpression)indi-catethattheshorttranscriptD01canpartiallyrescueFigure3Expressionlevelsofsma-9transcriptsinmutantbackgrounds.
Expressionlevelsofsma-9transcriptsasdeterminedbyqRT-PCR.
AnimalswerecollectedattheL3stage.
Dataareshownastranscriptabundancerelativetoact-1actingenecontrol,anderrorbarsshowstandarddeviation.
SeeTable1forquantitationoftranscriptabundancerelativetoexon-9-containingtranscripts.
(-):transcriptharborsaprematureterminationcodon.
(+):transcriptsequenceisnormal.
050100150200250300350sma-9(qc3);him-5sma-9(wk55);him-5sma-9(cc604);him-5him-5TranscriptLevel(relativetoactincontrol)A11exon9D01(+)(+)(+)(+)(+)(+)(-)(-)(-)(-)(-)Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page8of11maletailbutnotbodysizedefectsofsma-9mutants.
IncontrasttotheresultsonD01,wefindfortheyk1285a11(A11)transcriptlittleornoevidenceofaninvivofunc-tion,aswellaslowabundancethroughoutdevelopment.
Inaddition,classIandIIfull-lengthtranscriptsandtheD01shorttranscript,butnottheA11transcript,encodetheZnfingertripletdomainshowntobecriticalintheregulationofbodysizeandmesodermalpatterning[14].
InDrosophila,thisdomainisalsocriticalinmediatingDpp-responsivetargetgeneregulation[35].
ThegeneproductencodedbytheA11transcriptlacksthisdomainandmaythereforenotbecapableofbeingrecruitedtotargetgenes.
WhythenistheA11transcriptdetectedatallOnepossibilityisthatitisgeneratedasabyproductofthetrans-splicingmechanismthatgeneratesD01.
Sincewehaveshownthatfull-lengthtranscriptsandD01transcriptsareeachnecessarybutnotsufficientformaletailpatterning,wemustconsiderwhatdifferencesinmolecularfunctionarefoundintherespectivegeneprod-ucts.
Inotherorganisms,Schnurrihomologshavebeenshowntoacteitherastranscriptionalactivators[20,21,36]orastranscriptionalrepressors[16,37]mediat-ingBMP-dependenttranscription.
Strikingly,Yaoetal.
[21]haveshownbyswappingDrosophilaandXenopusSchnurrihomologsthattranscriptionalactivationorrepressionactivitiesmaybecontext-dependentratherthananintrinsicpropertyoftheprotein.
InC.
elegans,wehavepreviouslyanalyzedintrinsictranscriptionalactivi-tiesofSMA-9proteindomainsusingaheterologoustran-scriptionassay[33].
Intheseexperiments,theacidicresidue-richdomain(ARD)inexon14exhibitedtran-scriptionalactivatoractivity,whiletheN-terminalregionofSMA-9displayedintrinsictranscriptionalrepressoractivity[33].
Full-lengthsma-9transcriptsshouldencodeproteinproductscontainingbothofthesetranscriptionaldomains,sothattheirtranscriptionalactivitiesmaybecontext-dependentaswasdescribedforDrosophilaandXenopusSchnurri[21].
Ontheotherhand,D01tran-scriptsencodeproductscontainingsolelythetranscrip-tionalactivationdomain(Figure1).
Wethereforehypothesizethatfull-lengthSMA-9isoformsfunctionastranscriptionalrepressorsoractivatorsdependingoncontext,whileD01-encodedisoformsmaybeobligatetranscriptionalactivators.
Thistranscriptionalactivationactivitymaybenecessaryfortherobustactivationofsma-9targetgenesinsensoryraydevelopment,butmaybedispensableinbodysizeregulation.
Consistentwiththishypothesis,aSMA-9Znfingerdomainfusionwithaknowntranscriptionalactivator,VP16,canpartiallyres-cuethemaletaildefectsbutnotthebodysizedefectsofsma-9mutants.
IncontrasttotheSMA-9::VP16fusion,aSMA-9::enRrepressiondomainfusionrescuesbodysize,suggestingthattranscriptionalrepressionismoreimpor-tantthanactivationforSMA-9activityinbodysizeregu-lation[33].
Overall,theinvivoactivityoftheSMA-9::VP16fusionisstrikinglysimilartothatoftheD01iso-form.
Alternativesplicingisregulatedbytrans-actingfactorsthatinfluencethechoiceofsplicesites.
Itwillbeofinter-esttodeterminethetrans-actingfactorsthatregulatealternativetrans-splicingofsma-9.
Inpreliminaryexperi-ments,wetestedwhethersma-9trans-splicingdependsonmec-8,whichencodesanRNArecognitionmotif-con-tainingproteinthatregulatesalternativesplicinginthehypodermis[38,39].
WefoundthatA11,D01,andexon9-containingtranscriptlevelswereallreduced2-to5-foldinmec-8mutantsrelativetoN2(datanotshown).
Ifmec-8weredirectlyinvolvedinprocessingtheshorttran-Figure4Phenotypicrescuebyoverexpressionofsma-9shorttranscripts.
A.
Maletailphenotypesaregivenasfrequencyoffusionofrays8and9permaletailsidescored.
pvalueswerecalculatedusingaStudent'st-test.
*:significantdifferencefromnocDNAcontrol(p<0.
05).
**:highlysignif-icantdifferencefromnocDNAcontrol(p<0.
01).
ThepvaluesforexperimentswithA11cDNAoverexpressionshowednosignificantdifferencefromcontrols(p=0.
59forqc3;p=0.
16forcc604).
Allstrainscontainhim-5(e1490)toincreasetheproportionofmales.
B.
Bodysizephenotypes.
Meanbodylengthat96hoursafterembryocollection(adultstage)isshown.
Errorbarsshowstandarddeviation.
Notethatthesestrainsaretransgenicsexpress-ingtherol-6markerthatinfluencesbodylength,soallcomparisonsaremadebetweenheat-shockedanimalsandnoheatshockcontrolratherthanusinganontransgeniccontrol.
pvalueswerecalculatedusingaStudent'st-test.
**:highlysignificantdifferencefromnoheatshockcontrol(p<0.
01).
Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page9of11scriptsfromalongerprecursor,weexpectedtofindreciprocalchangesinaccumulationofthefull-lengthtranscriptsrelativetoshorttranscripts.
OurresultsarethereforenotsufficientlycleartodrawaconclusionastowhetherMEC-8isdirectlyinvolvedinsma-9trans-splic-ing,andadditionalinvestigationswillbenecessarytoidentifythetrans-actingfactorsthatmediatethisreac-tion.
Avarietyofgeneticmechanismsexistforgeneratingdiversefunctionsfromasinglegeneticlocus,includingthosethataffecttranscriptstructuresuchasalternativesplicing,alternativetranscriptionalstartsites,andalter-nativepolyadenylation.
Oneprominentexampleofalter-nativesplicingistheregulationofsexdeterminationinDrosophilabySex-lethal[40].
Thissystemalsousesdevelopmentallyregulatedalternativepromoters.
InC.
elegans,extensiveuseofalternativepromotersleadingtodifferentproteinisoformshasbeendocumented[41].
Alternativepolyadenylationcanalsoleadtotheproduc-tionoftruncatedproteinvariants,suchasinthedaf-4BMPreceptorgeneinC.
elegans,inwhichalternativepolyadenylationleadstotheproductionofasecretednegativeinhibitorofDAF-4signaling[42].
Toourknowl-edge,alternativetrans-splicingaswehavecharacterizeditforsma-9hasnotpreviouslybeenreportedforotherlociinC.
elegans.
Theuniquefeatureofthistrans-splic-ingeventisthechoicebetweenprocessingthetranscriptviacis-ortrans-splicingatthespliceacceptorsiteofpre-dictedexon11.
Inanorganismwithouttrans-splicing,asimilartruncationcouldbeachievedviauseofadown-streamtranscriptionalstartsiteorviaproteolyticcleav-age.
Forexample,intheHedgehogsignalingpathway,full-lengthtranscriptionfactorsoftheCi/Glifamilyaretranscriptionalactivatorsthat,uponproteolyticcleavage,areconvertedtotranscriptionalrepressors[43].
Wespec-ulatethatsincethetrans-splicingmachineryexistsinC.
elegans,itwasavailabletoberecruitedforthisunusualroleinthesma-9locus.
ConclusionsBothC.
eleganssma-9/schnurri[13]andthehumanhomologsShn-1andShn-3[23,24]displayalternativesplicingthatispredictedtoresultinmultipleproteiniso-forms.
Formostoftheseisoforms,however,nospecificinvivofunctionhasbeendescribed.
Wehavenowdemon-stratedafunctionalroleforanalternativelytrans-splicedsma-9shorttranscriptrepresentedbycDNAcloneyk1237d01(D01).
Thistranscriptmediatesatissue-spe-cificrole,sinceitisrequiredforsensoryraydevelopmentbutnotforbodysizeregulation.
Ourresultsprovideinsightintohowasinglegeneticlocuscancontributetodiverseproteinfunctions.
MethodsC.
elegansstrainsandcultureNematodeswereculturedusingstandardmethodsandgrownat20°Cunlessotherwisenoted[44].
Inadditiontostrainsgeneratedinthiswork,thefollowingstrainswereused:N2(wildtype);LGV,him-5(e1490);LGX,sma-9(qc3,wk55,cc604).
RT-PCRRT-PCRwasperformedintwostepsusingSuperScriptIIIFirst-StrandSynthesisSystem(Invitrogen)andRNAextractedfrommixed-stageN2wild-typeanimals.
Tem-peraturegradientPCRwasperformedonanEppendorfgradientthermalcycler(MasterCycler)followingthemanufacturer'sinstructions.
RealtimeRT-PCRwasperformedonaLightCycler2.
0(Roche).
DataanalysissoftwareusedwasLightCyclersoftwareversion4.
0.
AnimalswerecollectedatdesiredtimepointsandtotalRNAextractedbyTrizolasdescribed[13].
SuperScriptTMIIIplatinumtwo-stepqRT-PCRkitwithSYBRGreen(Invitrogen)wasusedtoperformRT-PCR.
Actingeneact-1wasusedasstandardcontrol.
Aprimerinanintronoftheact-1genewasusedtoconfirmtheabsenceofgenomicDNAinRNApreps.
PrimersequencesforallRT-PCRexperimentsareavail-ableuponrequest.
RNAiRNAifeedingmethodwasperformedasdescribed[29]aftercloningsma-9fragmentsintoRNAifeedingvectorpPD129.
36(agiftfromDr.
A.
Fire).
Toscoresensoryraypatterning,onlyadultmalesdisplayingasmallphenotypewerecollectedandscoredtoensurethattheanimalsana-lyzedhadundergoneaknockdownofsma-9activity.
Heat-shockcloneconstructsandgenerationoftransgenicanimalsTwosma-9splicevariantsyk1285a11andyk1237d01(GenBankAccessionNumbers:AY390537andAY390550)wereidentifiedpreviously[13],whichwereSL1-andSL2-trans-splicedcDNAclonesthatappearedtorepresentshorterbutcompletemRNAs.
Heat-shockvectorpPD49.
83wasakindgiftfromDr.
J.
Liu.
yk1285a11andyk1237d01wereclonedintopPD49.
83atNheIandKpnIsites.
Transgenicnematodesweregener-atedbymicroinjectionofconstructs(10ng/ul)intothegonadalsyncytiaofhim-5hermaphrodites,withrol-6(100ng/ul)asamarker[45].
Mutantstrainscarryingtransgenicconstructsweregeneratedbyappropriatecrossesbetweenthemutantstrainsandtransgeniclines.
Heat-shockexperimentsGravidhermaphroditeswerewashedoffplateswithM9bufferandcollectedinanEppendorftube.
Hermaphro-Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page10of11ditesweredestroyedwithsodiumhypochloritesolution,leavingeggsthatwerecollectedandinoculatedontoOP50-seededplates.
Forbodysizemeasurement,24hoursaftereggcollection,wormswerecollectedandtransferredtoasiliconizedEppendorftubecontaining100ulofM9buffer[44],whichwasplacedinacirculatingwaterbathundertheheat-shockconditionsspecified.
Afterheat-shock,wormswererecovered,placedat25°ConOP50-seededplatesandallowedtodeveloptoadult-hood.
ThelengthofindividualwormswasmeasuredusingImage-ProExpresssoftware(Sigma)withNomar-skioptics.
Formaletailrayexamination,wormswerecol-lected72hoursaftereggcollectionandtransferredtoasiliconizedEppendorftubecontaining100ulofM9buf-fer[44],whichwasplacedinacirculatingwaterbathundertheheat-shockconditionsspecified.
Afterheat-shock,wormswererecovered,placedat25°ConOP50-seededplatesandallowedtodeveloptoadulthoodandthenscoredforraypatterning.
Authors'contributionsJYperformedtheheatshockandRT-PCRexperimentsandparticipatedininterpretationofdata.
LYperformedtheRNAiexperiments.
CSDconceivedofthestudy,participatedinitsdesignandcoordination,analyzedmutantpheno-types,anddraftedthemanuscript.
Allauthorsreadandapprovedthefinalmanuscript.
AcknowledgementsWethankJunKellyLiuforsharingreagentsandforhelpfuldiscussions.
SomeoftheexperimentsweredonewithequipmentfromtheCoreFacilitiesforImag-ing,CellularandMolecularBiologyatQueensCollege.
SomeC.
elegansmutantstrainswereobtainedfromtheCaenorhabditisGeneticsCenter,whichissup-portedbytheNIHNationalCenterforResearchResources(NCRR).
ThisworkwascarriedoutinpartialfulfillmentoftherequirementsforthePh.
D.
degreefromtheGraduateCenterofCityUniversityofNewYork(J.
Y.
).
ThisresearchwassupportedbyRSG-98-230-04-DDCfromtheAmericanCancerSocietytoC.
S.
-D.
AuthorDetailsDepartmentofBiology,QueensCollege,andBiochemistryPhDProgram,theGraduateSchoolandUniversityCenter,theCityUniversityofNewYork,Flushing,NY11367,USAReferences1.
RobertsAB,SpornMB:Physiologicalactionsandclinicalapplicationsoftransforminggrowthfactor-beta(TGF-beta).
GrowthFactors1993,8(1):1-9.
2.
MassagueJ,ChenYG:ControllingTGF-betasignaling.
GenesDev2000,14(6):627-644.
3.
WranaJL,AttisanoL,CarcamoJ,ZentellaA,DoodyJ,LaihoM,WangXF,MassagueJ:TGFbetasignalsthroughaheteromericproteinkinasereceptorcomplex.
Cell1992,71(6):1003-1014.
4.
LiuF,VenturaF,DoodyJ,MassagueJ:HumantypeIIreceptorforbonemorphogenicproteins(BMPs):extensionofthetwo-kinasereceptormodeltotheBMPs.
MolCellBiol1995,15(7):3479-3486.
5.
InmanGJ,NicolasFJ,HillCS:NucleocytoplasmicshuttlingofSmads2,3,and4permitssensingofTGF-betareceptoractivity.
MolCell2002,10(2):283-294.
6.
Macias-SilvaM,AbdollahS,HoodlessPA,PironeR,AttisanoL,WranaJL:MADR2isasubstrateoftheTGFbetareceptoranditsphosphorylationisrequiredfornuclearaccumulationandsignaling.
Cell1996,87(7):1215-1224.
7.
KretzschmarM,LiuF,HataA,DoodyJ,MassagueJ:TheTGF-betafamilymediatorSmad1isphosphorylateddirectlyandactivatedfunctionallybytheBMPreceptorkinase.
GenesDev1997,11(8):984-995.
8.
WuRY,ZhangY,FengXH,DerynckR:HeteromericandhomomericinteractionscorrelatewithsignalingactivityandfunctionalcooperativityofSmad3andSmad4/DPC4.
MolCellBiol1997,17(5):2521-2528.
9.
ZhangY,FengX,WeR,DerynckR:Receptor-associatedMadhomologuessynergizeaseffectorsoftheTGF-betaresponse.
Nature1996,383(6596):168-172.
10.
InmanGJ,HillCS:Stoichiometryofactivesmad-transcriptionfactorcomplexesonDNA.
JBiolChem2002,277(52):51008-51016.
11.
LagnaG,HataA,Hemmati-BrivanlouA,MassagueJ:PartnershipbetweenDPC4andSMADproteinsinTGF-betasignallingpathways.
Nature1996,383(6603):832-836.
12.
Savage-DunnC:TGF-βsignaling.
WormBook2005,9:1-12.
13.
LiangJ,LintsR,FoehrML,TokarzR,YuL,EmmonsSW,LiuJ,Savage-DunnC:TheCaenorhabditiselegansschnurrihomologsma-9mediatesstage-andcelltype-specificresponsestoDBL-1BMP-relatedsignaling.
Development2003,130(26):6453-6464.
14.
FoehrML,LindyAS,FairbankRC,AminNM,XuM,YanowitzJ,FireAZ,LiuJ:AnantagonisticrolefortheC.
elegansSchnurrihomologSMA-9inmodulatingTGFbetasignalingduringmesodermalpatterning.
Development2006,133(15):2887-2896.
15.
DaiH,HoganC,GopalakrishnanB,Torres-VazquezJ,NguyenM,ParkS,RafteryLA,WarriorR,AroraK:ThezincfingerproteinschnurriactsasaSmadpartnerinmediatingthetranscriptionalresponsetodecapentaplegic.
DevBiol2000,227(2):373-387.
16.
MartyT,MullerB,BaslerK,AffolterM:SchnurrimediatesDpp-dependentrepressionofbrinkertranscription.
NatCellBiol2000,2(10):745-749.
17.
TakagiT,HaradaJ,IshiiS:MurineSchnurri-2isrequiredforpositiveselectionofthymocytes.
NatImmunol2001,2(11):1048-1053.
18.
OukkaM,KimST,LugoG,SunJ,WuLC,GlimcherLH:AmammalianhomologofDrosophilaschnurri,KRC,regulatesTNFreceptor-drivenresponsesandinteractswithTRAF2.
MolCell2002,9(1):121-131.
19.
WuLC:ZAS:C2H2zincfingerproteinsinvolvedingrowthanddevelopment.
GeneExpr2002,10(4):137-152.
20.
JinW,TakagiT,KanesashiSN,KurahashiT,NomuraT,HaradaJ,IshiiS:Schnurri-2controlsBMP-dependentadipogenesisviainteractionwithSmadproteins.
DevCell2006,10(4):461-471.
21.
YaoLC,BlitzIL,PeifferDA,PhinS,WangY,OgataS,ChoKW,AroraK,WarriorR:SchnurritranscriptionfactorsfromDrosophilaandvertebratescanmediateBmpsignalingthroughaphylogeneticallyconservedmechanism.
Development2006,133(20):4025-4034.
22.
ShuklaA,MalikM,CataissonC,HoY,FriesenT,SuhKS,YuspaSH:TGF-betasignallingisregulatedbySchnurri-2-dependentnucleartranslocationofCLIC4andconsequentstabilizationofphospho-Smad2and3.
NatCellBiol2009,11(6):777-784.
23.
MuchardtC,SeelerJS,NirulaA,ShurlandDL,GaynorRB:RegulationofhumanimmunodeficiencyvirusenhancerfunctionbyPRDII-BF1andc-relgeneproducts.
JVirol1992,66(1):244-250.
24.
HicarMD,LiuY,AllenCE,WuLC:StructureofthehumanzincfingerproteinHIVEP3:molecularcloning,expression,exon-intronstructure,andcomparisonwithparalogousgenesHIVEP1andHIVEP2.
Genomics2001,71(1):89-100.
25.
BlumenthalT:Trans-splicingandoperons.
WormBook2005,25:1-9.
26.
HuangT,KuerstenS,DeshpandeAM,SpiethJ,MacMorrisM,BlumenthalT:Intercistronicregionrequiredforpolycistronicpre-mRNAprocessinginCaenorhabditiselegans.
MolCellBiol2001,21(4):1111-1120.
27.
BlumenthalT,EvansD,LinkCD,GuffantiA,LawsonD,Thierry-MiegJ,Thierry-MiegD,ChiuWL,DukeK,KiralyM,etal.
:AglobalanalysisofCaenorhabditiselegansoperons.
Nature2002,417(6891):851-854.
28.
LameschP,MilsteinS,HaoT,RosenbergJ,LiN,SequerraR,BosakS,Doucette-StammL,VandenhauteJ,HillDE,etal.
:C.
elegansORFeomeversion3.
1:increasingthecoverageofORFeomeresourceswithimprovedgenepredictions.
GenomeRes2004,14(10B):2064-2069.
29.
KamathRS,AhringerJ:Genome-wideRNAiscreeninginCaenorhabditiselegans.
Methods2003,30(4):313-321.
30.
SijenT,FleenorJ,SimmerF,ThijssenKL,ParrishS,TimmonsL,PlasterkRH,FireA:OntheroleofRNAamplificationindsRNA-triggeredgenesilencing.
Cell2001,107(4):465-476.
Received:3March2010Accepted:17June2010Published:17June2010Thisarticleisavailablefrom:http://www.
biomedcentral.
com/1471-2199/11/462010Yinetal;licenseeBioMedCentralLtd.
ThisisanOpenAccessarticledistributedunderthetermsoftheCreativeCommonsAttributionLicense(http://creativecommons.
org/licenses/by/2.
0),whichpermitsunrestricteduse,distribution,andreproductioninanymedium,providedtheoriginalworkisproperlycited.
BMCMolecularBiology2010,11:46Yinetal.
BMCMolecularBiology2010,11:46http://www.
biomedcentral.
com/1471-2199/11/46Page11of1131.
BosherJM,DufourcqP,SookhareeaS,LabouesseM:RNAinterferencecantargetpre-mRNA:consequencesforgeneexpressioninaCaenorhabditiselegansoperon.
Genetics1999,153(3):1245-1256.
32.
MangoSE:StopmakingnonSense:theC.
eleganssmggenes.
TrendsGenet2001,17(11):646-653.
33.
LiangJ,YuL,YinJ,Savage-DunnC:TranscriptionalrepressorandactivatoractivitiesofSMA-9contributedifferentiallytoBMP-relatedsignalingoutputs.
DevBiol2007,305(2):714-725.
34.
LintsR,EmmonsSW:PatterningofdopaminergicneurotransmitteridentityamongCaenorhabditiselegansraysensoryneuronsbyaTGFbetafamilysignalingpathwayandaHoxgene.
Development1999,126(24):5819-5831.
35.
MullerB,HartmannB,PyrowolakisG,AffolterM,BaslerK:ConversionofanextracellularDpp/BMPmorphogengradientintoaninversetranscriptionalgradient.
Cell2003,113(2):221-233.
36.
Torres-VazquezJ,ParkS,WarriorR,AroraK:ThetranscriptionfactorSchnurriplaysadualroleinmediatingDppsignalingduringembryogenesis.
Development2001,128(9):1657-1670.
37.
PyrowolakisG,HartmannB,MullerB,BaslerK,AffolterM:AsimplemolecularcomplexmediateswidespreadBMP-inducedrepressionduringDrosophiladevelopment.
DevCell2004,7(2):229-240.
38.
SpikeCA,DaviesAG,ShawJE,HermanRK:MEC-8regulatesalternativesplicingofunc-52transcriptsinC.
eleganshypodermalcells.
Development2002,129(21):4999-5008.
39.
LundquistEA,HermanRK,RogalskiTM,MullenGP,MoermanDG,ShawJE:Themec-8geneofC.
elegansencodesaproteinwithtwoRNArecognitionmotifsandregulatesalternativesplicingofunc-52transcripts.
Development1996,122(5):1601-1610.
40.
BlackDL:Mechanismsofalternativepre-messengerRNAsplicing.
AnnuRevBiochem2003,72:291-336.
41.
ChoiJ,NewmanAP:Atwo-promotersystemofgeneexpressioninC.
elegans.
DevBiol2006,296(2):537-544.
42.
GuntherCV,RiddleDL:Alternativepolyadenylationresultsinatruncateddaf-4BMPreceptorthatantagonizesDAF-7-mediateddevelopmentinCaenorhabditiselegans.
JBiolChem2004,279(38):39555-39564.
43.
HuangfuD,AndersonKV:SignalingfromSmotoCi/Gli:conservationanddivergenceofHedgehogpathwaysfromDrosophilatovertebrates.
Development2006,133(1):3-14.
44.
BrennerS:ThegeneticsofCaenorhabditiselegans.
Genetics1974,77(1):71-94.
45.
MelloCC,KramerJM,StinchcombD,AmbrosV:EfficientgenetransferinC.
elegans:extrachromosomalmaintenanceandintegrationoftransformingsequences.
EmboJ1991,10(12):3959-3970.
doi:10.
1186/1471-2199-11-46Citethisarticleas:Yinetal.
,Alternativetrans-splicingofCaenorhabditisele-ganssma-9/schnurrigeneratesashorttranscriptthatprovidestissue-specificfunctioninBMPsignalingBMCMolecularBiology2010,11:46
- larvalwww.39555.com相关文档
- 亮度www.39555.com
- open55
- 登录www
香港ceranetworks(69元/月) 2核2G 50G硬盘 20M 50M 100M 不限流量
香港ceranetworks提速啦是成立于2012年的十分老牌的一个商家这次给大家评测的是 香港ceranetworks 8核16G 100M 这款产品 提速啦老板真的是豪气每次都给高配我测试 不像别的商家每次就给1核1G,废话不多说开始跑脚本。香港ceranetworks 2核2G 50G硬盘20M 69元/月30M 99元/月50M 219元/月100M 519元/月香港ceranetwork...
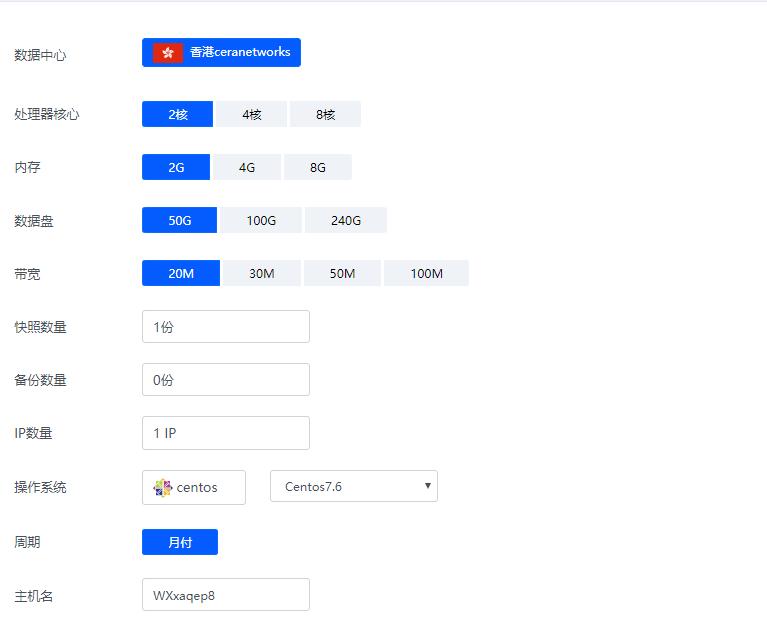
iON Cloud:新加坡cn2 gia vps/1核/2G内存/25G SSD/250G流量/10M带宽,$35/月
iON Cloud怎么样?iON Cloud升级了新加坡CN2 VPS的带宽和流量最低配的原先带宽5M现在升级为10M,流量也从原先的150G升级为250G。注意,流量也仅计算出站方向。iON Cloud是Krypt旗下的云服务器品牌,成立于2019年,是美国老牌机房(1998~)krypt旗下的VPS云服务器品牌,主打国外VPS云服务器业务,均采用KVM架构,整体性能配置较高,云服务器产品质量靠...

PhotonVPS:$4/月,KVM-2GB/30GB/2TB/洛杉矶&达拉斯&芝加哥等
很久没有分享PhotonVPS的消息,最近看到商家VPS主机套餐有一些更新所以分享下。这是一家成立于2008年的国外VPS服务商,Psychz机房旗下的站点,主要提供VPS和独立服务器等,数据中心包括美国洛杉矶、达拉斯、芝加哥、阿什本等。目前,商家针对Cloud VPS提供8折优惠码,优惠后最低2G内存套餐每月4美元起。下面列出几款主机配置信息。CPU:1core内存:2GB硬盘:30GB NVm...
www.39555.com为你推荐
-
敬汉卿姓名被抢注身份证信息被抢注12306账号怎么办原代码什么叫源代码,源代码有什么作用陈嘉垣马德钟狼吻案事件是怎么回事seo优化工具SEO优化神器有什么比较好的?百度关键词工具常见百度关键词挖掘方法分别是什么请列举?百度关键词分析如何正确分析关键词?抓站工具仿站必备软件有哪些工具?最好好用的仿站工具是那个几个?16668.com香港最快开奖现场直播今晚开www.175qq.com最炫的qq分组www.dm8.cc有没有最新的日本动漫网站?