同源cv狗狗
cv狗狗 时间:2021-05-25 阅读:()
犬冠状病毒南京株M基因的同源重组分析3王玉燕陆承平33(南京农业大学农业部动物疫病诊断及免疫重点实验室,江苏南京,210095)RecombinationAnalysisofMProteinofaCanineCoronavirusWildStrainIsolatedFromNanjingCityWANGYu2yan,LUCheng2ping33(1.
KeyLabofAnimalDieaseDiagnosticandImmunologyMinistryofAgriculture,NanjingAgricUniv,Nanjing210095,ChinaAbstract:TwoMgenesequencesofCaninecoronavirus(CCV),fromawildstrainNJ17fromadiarrheadogfaecalsampleinNanjingCityandCCV1271referencestrain,wereclonedandse2quenced.
Theresultsshowed,CCV1271sharedhighsimilarity(98.
9~99.
5%)toCCVV1,V2andaCCVstrainfromgiantpandaisolatedinChina2003.
ItsuggestedthattheaboveCCVstrainsexistedquasispeciesfromasameancestorstrain.
OnMgene,CCVNJ17shared87.
0~91.
9%identitieswithotherCCVstrainsandtypicalstrainsofTransmissiblegastroenteritisvirus(TGEV)ofswine,andFelinecoronaviruses(FCoVs)intheGenBank.
TheCCVNJ17mightbeaseparatedevolutionarystraininChina.
RecombinationmarksequencesofallthelandedstrainsontheMgeneputativerecombinationregionhada"CTTTAG"nucleotides,meetingto"CTT(A/T)(A/T)G"thatalwaysappearedneartotherecombination"hotspot"inInfectiousbron2chitisvirusRNAgenome.
Thefirst50aminoacidresiduesonNterminalofCCVNJ17MproteinwashighersimilartoFCoV7921683thanTGEV,whilethefollowingaminoacidresiduese2quencewasclosertoTGEVthanFCoV7921683.
Itindicatedthatarecombinationeventhadhap2penonMgeneofNJ17.
StructureandfunctionpredictionrevealedalltheCCVstrains'Mproteinhadasamesecretorysignalpeptide,fourtransmembraneregions,aminoacidandcarboxylicacidterminalinsertingintotheenvelope.
Putativetrimmedfunctionalsiteswerereserved,andstron2gerantigenicitieswerefoundonthetwoterminalsofMprotein.
Withreservedfunctionalsites,theaminoacidterminalresidueswerehighlyvariedanditsactionseemedtoberelatedtoitsstructuredomainsbutitssequence.
Keywords:Caninecoronavirus(CCV);Mprotein;Similarityrecombination摘要:对来自腹泻犬粪样的犬冠状病毒(CCV)南京株NJ17株及参考株1271的M基因进行了克隆、测序,并与GenBank中所有已知CCV毒株及同亚群的猪冠状病毒(TGEV)和猫冠状病毒(FCoV)代表株的M基因进行了同源性比较和系统进化分析,同时对M蛋白的结构和功能进行了预测分析.
结果表明,CCV1271与近年在中国分离到的CCV毒株V1、V2及大熊猫源的毒株具有98.
9%~99.
5%的同源性,说明这些毒株可能是来自同一毒株的准种.
NJ17与其他中国分离株及国外分离株的同源性为87.
0%~91.
9%,显示国内可能存在一个相对独立进化的CCV毒株.
序列比较发现,所有CCV毒株在可能的同源重组"热点"区内都有一个CTTTAG序列,与鸡传染性支气管炎病毒同源重组模板交换位点附近的特征序列相似.
CCVNJ17株M蛋白在N端50氨基酸序列与FCoV7921683同源性高,而在后212氨基酸序列与TGEV同源性高,提示该毒株可能在M基因上曾经发生过不同病毒收稿日期:2004211209,修回日期:20052032153基金项目:上海科技兴农重点公关项目(农科攻字(2003)第D21号);"江苏省高等学校研究生创新计划"资助项目.
作者简介:王玉燕(19762),女,山东省籍,博士生,研究方向为兽医病毒学.
E2mail:yuyanss@163.
com33通讯作者.
陆承平(19452),男,上海省籍,教授,兽医微生物学与免疫学.
Tel:025284396517,E2mail:lucp@njau.
edu.
cn1995-2006TsinghuaTongfangOpticalDiscCo.
,Ltd.
Allrightsreserved.
的同源重组.
CCVM蛋白的结构及功能预测表明,所有毒株都具有分泌型信号肽,有4个螺旋跨膜区,N末端和C末端均位于膜内.
M蛋白的两末端具有较强的抗原性,M蛋白上存在多种功能性氨基酸修饰位点且相对保守.
N末端的氨基酸变异很大,但是功能性修饰位点相对保守,提示N末端的功能可能与构象有关.
关键词:犬冠状病毒;M蛋白;同源重组中图分类号:S852.
65文献标识码:A文章编号:100325125(2005)0320262206犬状病毒(Caninecoronavirus,CCV)是引起犬急性胃肠炎的重要病原,CCV与猪传染性胃肠炎病毒(Transmissiblegastroenteritisvirus,TGEV)、猫的冠状病毒(Felinecoronaviruses,FCoV)、人冠状病毒229E(Humancoronavirus229E,HCoV229E)等同属于冠状病毒第I群[1].
其中,CCV、FCoV、TGEV的在系统发生树上更为接近,组成一个冠状病G121亚群,而猪流行性腹泻病毒(Porcineepidemicdiarrheavirus,PEDV)和HCoV229E组成G122亚群[2].
不同动物来源的冠状病毒之间可发生同源重组[3,4].
Erles[5]等最近报道,证实冠状病毒也是犬传染性呼吸道综合征(caninein2fectiousrespiractrysyndromediseases,CIRD)的重要病原,其S基因与HCoV2OC43和牛冠状病毒(BCoV)同源性最高,可能来源于OC43或BCoV.
在2003年震惊世界的SARSCoV,很可能就是不同宿主来源冠状病毒重组的结果[6].
CCV不断有新的CCV变异株被发现,而且有较广的宿主范围[7~9],对CCV的研究有潜在的公共卫生意义.
CCV是正股单链RNA病毒,基因组长约27~32Kb,其中2/3的基因组编码一个大的聚合酶蛋白(ORF1a、1b),在ORF1b的下游是几个编码蛋白的ORFs,包括S、E、M、N蛋白及一些可能的非结构蛋白[1].
冠状病毒基因组RNA在复制时采用独特的套式转录,基因组的错配率和重组率都很高[3].
研究发现冠状病毒基因组RNA重组现象的发生是普遍且随机的,几乎在每个位点上都可能发生.
但是,重组位点更趋向于集中在一些重组的"热点(hotspot)"区域,而且总有一些特征的核酸序列出现在其附近[10].
例如,对小鼠肝炎冠状病毒(MHV)研究发现,多数重组体病毒RNA的重组位点在前导序列和5核苷酸重复序列"UCUAA"之间[11].
而"CTT(A/T)(A/T)G"总出现在在鸡传染性支气管炎病毒(IBV)同源重组模板交换位点附近[12].
对冠状病毒同源重组热点的研究多集中在聚合酶基因、5′和3′端的非翻译区及变异较大的S基因上.
冠状病毒的M基因相对保守,但是也有研究认为在M基因上存在RNA同源重组的"热点",Herrevegh等研究证实,FCoVⅡ型毒株7921683的M基因327~376碱基之间存在FCoVⅠ和CCV同源模板RNA交换重组位点[4].
作者在此前的研究中发现,在中国犬群中有较高的CCV感染率,而且健康犬、狐、貉携带CCV的比率也很高.
通过比较M基因的保守区发现,来自南京腹泻犬粪样的CCVNJ17与其他毒株的序列同源性较低,可能是一个相对独立的CCV毒株.
本研究用PCR方法克隆了CCVNJ17株和参考株1271株的M基因.
将得到的M基因序列与Gen2BanK中所有CCV毒株的M基因和与CCV同亚群TGEV、FCoV的代表毒株做比较分析,在其中寻找其他冠状病毒RNA同源重组研究中发现的具有模板交换"热点"特征的序列,同时对相应的M蛋白的结构及功能特性进行了预测分析.
1材料与方法1.
1细胞及参考毒株CCV21271参考株及A72细胞(犬纤维肉瘤细胞系)均由德国吉森大学Baljer教授惠赠.
后者用含10%新生犊牛血清的MEM营养液培养,CCV1271增殖维持液为含2%新生犊牛血清的MEM.
1.
2样本采集及处理CCV阳性粪样NJ17于2003年10月采自南京某宠物门诊,用pH7.
40.
01mol/L的PBS进行5倍稀释后,5000r/min4℃离心20min,取上清保存于-20℃待检.
将CCV1271接种A72细胞,至80%以上细胞出现病变后冻融3次收获病毒,-20℃保存备用.
1.
3RT2PCR取200μL粪便上清加1mLTrizol(32ZOL)(MDBio公司产品)按说明书进行操作,提取RNA.
使用RevertAidTMFisrtStrandcDNASynthesisKit(MBI),按操作说明进行cDNA合成.
参照文献[7]发表的引物序列合成扩增M基因全长的引物MF(5′2CCTTGTTTGAACTAAACAAAATGAAG23′),MR(5′2TCCCTGAGAGGCCATTTAGA23′),引物由上海皓嘉公司合成.
反应体系如下:模板DNA5μL,10*PCR缓冲液2.
5μL,25mmol/LMgCl22μL,10mMdNTPs1μL,引362王玉燕,等:犬冠状病毒南京株M基因的同源重组分析1995-2006TsinghuaTongfangOpticalDiscCo.
,Ltd.
Allrightsreserved.
物各0.
5μl(30pmol/μL),exTaqDNA聚合酶(5U/μL,TaKaRa公司)0.
5μL,用ddH2O调整终体积至25μL.
反应条件:94℃30s、55℃30s、72℃1min,扩增35个循环,最后72℃延伸5min.
1.
4测序及分析PCR产物用胶回收试剂盒回收(北京鼎国公司),连接T载体,(TaKaRa公司),转化大肠杆菌DH5α,挑取阳性克隆,由上海基康公司进行测序.
测序结果在GenBanK上进行比对,然后用DNAstarMegAlin软件与GenBank上发表的所有CCVM基因及冠状病毒G121亚群的TGEV和FCV的代表毒株进行序列比对和同源性比较(表1),应用ClustalX和TreeView软件绘制系统进化树.
同时,在序列中寻找可能在同源重组"热点"附近可能出现的特征性序列,然后分段进行比较分析.
在http://www.
predictprotein.
org网站进行相关毒株M蛋白的结构及功能预测分析.
用Sig2nalP3.
0Server(http://www.
cbs.
dtu.
dk)进行信号肽的预测,DNAStarProtean进行蛋白的抗原性分析和表面结构域预测.
表1CCV及相关冠状病毒G121亚群代表毒株背景Table1BordereauofCCVsandotherG121typicalcorona2virusesinthisarticleStrainsGenBankAccessionNO.
Length(bp)GenotypecountryIsolatedtimeCCV1271AY7049163789CCVⅡGermen1971CCVNJ17AY7049173792CCVⅡChina2004CCVV2AY390344792CCVⅡChina2003CCVV1AY390343792CCVⅡChina2003PandaCCVAY436635792CCVⅡChina2003CCVInsavc21D113096789CCVⅡU.
K.
1980sCCVBGF10AY342160789CCVⅡU.
K.
2004CCV259/01AF502583792CCVⅠItaly2002FECV7921683AB086904792FCoVⅡU.
S.
A.
1970sFIPVUCD1AB086902789FCoVⅠU.
S.
A.
1970sTGEVAJ270965789/Spain2000Note:3submittedbyauthors.
CCV:CanineCoronavirus,FECV:felineentericcoronavirus,FIPV:felineinfectiousperitonitisvirus,TGEV:TransmissibleGastroenteritisVirus.
2结果2.
1CCVM基因的核酸及氨基酸序列分析RT2PCR反应可扩出835bp的目的片段,通过测序后在GenBanK中进行比对确认扩增无误,并登录GenBanK获得登录号:CCV1271AY704916,CCVNJ17AY704917.
该序列中包含M基因的完整ORF,其中CCVNJ17的M基因全长为792bp,编码全长263个氨基酸的蛋白,CCV1271的M基因全长为789bp,编码蛋白为262个氨基酸.
用DNAStar软件同源性分析表明,两毒株完整M基因之间的同源性为91.
8%;与其他G121亚群的冠状病毒相比较,CCV1271和CCVNJ17都与CCVV2同源性最高,分别为99.
4%和91.
9%;CCV1271与TGEV的同源性(93.
5%)高于FECV7921683(85.
4%)和FIPVUCD1(81.
6%);CCVNJ17M基因与TEGV的同源性也都高于与FECV7921683和FIPVUCD1,但是都低于CCV1271与这些毒株的同源性(图1).
CCV1271与CCVV1、CCVV2和PandaCCV同源性很高,在系统发生进化树上位于同一分枝上.
而CCVNJ17却与所有已知CCVM基因的同源性都不高,在系统进化树上与TGEV位于一个分枝上.
图1CCVM基因与G121亚群冠状病毒代表株的核苷酸同源性比较Fig.
1NucleotideidentityanddivergenceoftheCCVandotherG121typicalCoronavirusesMgenenucleotidesequences序列比对发现,在CCVM基因的前150bp变异明显,将M基因的核苷酸序列转化成相应的氨基酸序列,应用DNAStar软件分别对全长M蛋白、M蛋白前50氨基酸,和后212个氨基酸的进行同源性分析,并绘制系统进化树(图2,3,4).
无论完整M蛋白氨基酸序列比较还是分段比较,CCV1271都与CCVV1、CCVV2和PandaCCV在同一分枝上,且与TGEV的同源性高与FCV.
而CCVNJ17在完整M蛋白氨基酸序列比较中,与所有毒株的同源性都不高,但是在N端前50氨基酸中,与FECV7921683的同源性(77.
9%)高于TGEV(76.
9%),而在后212氨基酸片段中,与TGEV的同源性(94.
1%)高于FECV7921683(85.
5%),同样的结果也出现在系统进化树上.
寻找冠状病毒同源重组特征序列发现,所有CCV毒株在相应FECV7921683M基因同源重组的可能区域(3272376bp)内的序列都相对保守,而且在该区域内有一段序列"CTTTAG",符合在鸡传染性支气管炎病毒(IBV)重组研究中发现的特征序列"CTT(A/T)(A/T)G",后者总出现在IBV同源重462中国病毒学第20卷1995-2006TsinghuaTongfangOpticalDiscCo.
,Ltd.
Allrightsreserved.
图2CCVM蛋白氨基酸序列与G121亚群冠状病毒代表株的系统发生进化树Fig.
2PhylogenetictreeofCCVandotherG121typicalCoronavirusesMgeneaminoacidresiduesequences图3CCVM蛋白前50aa序列与G121亚群冠状病毒代表株比较的系统发生进化树Fig.
3PhylogenetictreeofCCVandotherG121typicalCoronavirusesMgenefirst50aminoacidresiduesequences图4CCVM蛋白后212aa序列与G121亚群冠状病毒代表株比较的系统发生进化树Fig.
4PhylogenetictreeofCCVandotherG121typicalCoronavirusesMgenelast212aminoacidresiduesequences组模板交换位点附近[12].
2.
2CCVM基因蛋白结构与功能预测SignalP预测表明所有毒株均在第15、16或17、18位氨基酸之间有信号肽的剪切位点,是典型的分泌型信号肽.
SYNOPSIS预测表明,所有M蛋白在结构上相似,分为膜内区、跨膜区和膜外区,均有4个螺旋跨膜区(如CCVNJ17的跨膜区分别位于55272、792103、1162136、2032220氨基酸残基间),N末端和C末端均位于膜内,两个膜外区.
DISULFIND预测分析在CCVNJ17、Insavc21、BGF10、259/01、FECV7921638、FIPVUCD1等毒株的第9(或10)位C与第31(或30)位C之间可能会形成两个二硫键,而TGEV则可能会在第9位和38位、14与40、15与20、30与157位间的C上形成二硫键.
抗原性分析表明各毒株的抗原位点分布较相似,且主要分布在跨膜区之外(图5,只给出CCVNJ17).
PROSITEmotifsearch寻找蛋白质的功能位点,结果显示在M蛋白上存在多种氨基酸修饰位点且相对保守,毒株之间略有差异(表2).
3讨论冠状病毒在动物中广泛分布以及易于变异而受到关注.
国内对CCV的研究较少,尤其是M基因的变异如何未见报道.
而M基因作为冠状病毒可能发生同源重组的区域之一,值得重视.
CCV1-71株由Binn等于1971年在发生腹泻的德国军犬中分离到,是世界上最早分离到的CCV毒株[13].
由同源性及进化关系上可看出,虽然分离的年代间隔较远,但是CCV1-71与近年在中国分离到几个CCV毒株V1、V2及大熊猫源的毒株具有98.
9%~99.
5%的同源性,说明这些毒株可能是来自同一毒株的准种.
同时表明虽然冠状病毒的变异频率很高,但在M基因上却相对保守.
本研究从腹泻病犬粪样中分离到的CCVNJ17在M基因上与CCV1271、Insavc21、BGF10、259/01等毒株及在中国分离到的其他毒株的同源性都不高(87.
0%~91.
9%),说明国内可能存在一个相对独立进化的CCV毒株.
有研究证实,在相对保守的CCVM基因上存在RNA同源重组的"热点"[4].
本研究通过序列比较发现,所有CCV毒株在对应FCoVⅡ型毒株79~83可能的同源重组"热点"区内都有一个"CTT2TAG"序列,与在IBV同源重组位点附近出现的特征序列"CTT(A/T)(A/T)G"相符.
该特征序列与冠状病毒RNA复制时的通用先导序列非常相似,可能会模拟RNA聚合酶的结合位点,导致RNA复制时模板链发生跳跃转移[12].
CCVNJ17株M蛋白在N末端前50氨基酸序列与FECV同源性高,而在后面的序列与TGEV同源性高,在两者之间具有"CTTTAG"序列,提示该毒株可能在M基因上发生过不同病毒的同源重组.
562王玉燕,等:犬冠状病毒南京株M基因的同源重组分析1995-2006TsinghuaTongfangOpticalDiscCo.
,Ltd.
Allrightsreserved.
表2CCVs及相关G121亚群代表毒株M蛋白功能性位点预测结果Table2PrositemotifpredictionofMproteinofCCVsandotherG121typicalCoronaviruses.
StrainsProbablyfunctionalmotifsiteASNPKCPCK2MYRISRYLAMIPROKARRGDASPCCVNJ1735、56、252/32、103、25427、122、173、185221///CCV127132、55、2512924、34、105、253121、172、1842205//CCVinsavc2132、55、2512940、25336、121、172、184220///CCVBGF1025、55、213、251/105、25135、121、172、184////CCV259/0133、56、2523035、41、254122、172、185////CCVV233、56、2523025、35、106、254122、173、1852216//PandaCCV33、56、2523025、35、106、254122、173、1852216//TGEV32、55、2512924、34、253121、172、1842205//FCV792168333、56、213、2513035、106、254122、173、185////FCVUCD135、55、251/40、25333、121、172、184162/4422Note:ASN:N2glycosylationsite,PKC:ProteinkinaseCphosphorylationsite,PCK2:CaseinkinaseIIphosphorylationsite,MYRISRYL:N2myristoylationsite,AMI:Amidationsite,PROKAR:Prokaryoticmembranelipoproteinlipidattachmentsite,RGD:Cellattachmentse2quence,ASP:Eukaryoticandviralaspartylproteasesactivesite.
图5CCVNJ17M蛋白的抗原性及表面结构域预测Fig.
5AntigenicandsurfaceprobabilitypredictionofCCVNJ17同源重组"热点"区对应的氨基酸位于跨膜区,也相对保守,但是其前面的氨基酸变异明显,尤其是N末端前50氨基酸(从信号肽到第一个跨膜区间的氨基酸),但是该区的几个功能相关位点却相对保守(表2),提示功能可能与构象有关而与氨基酸残基的一级结构无关.
Liao等认为,虽然冠状病毒基因组重组频繁,但是在环境的选择压力下只有某些具有优势的重组体被保存下来,大多数的重组体会被逐渐的淘汰[11].
Horsburgh等认为M蛋白上的N末端的变异与逃避宿主免疫系统压力有关[14].
因此对CCVNJ17M蛋白N末端的变异现象值得进一步研究.
M蛋白是冠状病毒组装和出芽所必需的,在病毒粒子的形成及与宿主的相互作用中起重要作用[15~17].
本研究通过预测分析发现,M蛋白的抗原性和功能区主要在两端.
已知M蛋白可诱导宿主产生α2干扰素[16],就TGEV而言,其诱导干扰素的功能与N末端几个糖基化的位点有关.
C末端几个功能性的氨基酸修饰位点在所有冠状病毒株中都存在,且非常保守,可能与诱导中和抗体产生,及补体依赖性的融细胞作用有关[17],其具体功能还需进一步研究.
本研究对CCVM蛋白结构和功能的预测为进一步研究M蛋白在病毒与宿主相互反应中的作用打下了基础.
致谢本研究在上海交通大学兽医生物技术研究所完成,实验中得到该所领导和孙建和博士的大力支持和帮助,在此深表谢意.
参考文献[1]CavanaghD,PrianDA,BVritonP,etal.
Nidovirales:anewordercomprisingcoronaviridaeandarteriviridae[J].
ArchVirol,1997,1452:6292635.
[2]GonzalezJM,PuertasG,CavanaghD,etal.
AcomparativesequenceanalysistorevisethecurrenttaxonomyofthefamilyCoronaviridae[J].
ArchVirol,2003,148(11):220722235.
[3]LaiSMMC,CavanaghD.
Themolevularbiologyofcorona2viruses[J].
AdvVirusRes,1997,48:12100.
[4]HerreveghAA,ISmeenkI,HorzinekMC,etal.
FelinecoronavirustypeIIstrain7921683and7921146originatefromadoublerecombinationbetweenfelinecoronavirustypeIandca2ninecoronavirus[J],JVirol,1998,72:450824514.
[5]ErlesK,ToomeyC,HarrietWB,etal.
Detectionofagroup2coronavirusindogswithcanineinfectiousrespiratorydisease[J].
Virology,2003,310:162223.
662中国病毒学第20卷1995-2006TsinghuaTongfangOpticalDiscCo.
,Ltd.
Allrightsreserved.
[6]StanhopeMJ,BrownJR,Amrine2MadsenH.
Evidencefromtheevolutionaryanylysisofnucleotidesequencesforarecombi2nanthistoryofSARS2CoV[J].
InfectGenetEvol,2004:15219.
[7]PratelliA,MartellaV,PistelloM,etal.
Identificationofcoronavirusesindogsthatsegregateseparatelyfromthecaninecoronavirusgenotype[J].
JVirolMeth,2003,107:2132222.
[8]JoseMS,ScottP,TimothyHM.
MolecularcharacterizationofavirulentcaninecoronavirusBGFstrain[J].
VirusRes,2004,140:27231.
[9]MatthewJH,GavanAH,RobertPM,etal.
IdentificationofcaninecoronavirusstrainsfromfesesbySgenenested2PCRandmolecularcharacterizationofanewAustralliaisolatep[J].
JClinMicrobio1,2001,39:103621041.
[10]BannerLR,LaiMM.
RandomnatureofcoronavirusRNArecombinationintheabsenceofselectionpressure[J].
Virol2ogy,1991,185(1):4412445.
[11]LiaoCL,LaiMM.
RNArecombinationinacoronavirus:recombinationbetweenviralgenomicRNAandtransfectedRNAfragments[J].
JVirol,1992,66:611726124.
[12]WangL,DavidJ,EllenWC.
Evidenceofnaturalrecombi2nationwithinS1geneofinfectiousbronchitisvirus[J].
Vi2rology,1993,192:7102716.
[13]BinnLN,LazarEC,KeenanKP,etal.
Recoveryandchar2acterizationofacoronavirusfrommilitarydogswithdiarrhea[J].
ProcAnnuMeetUSAnimHealthAssoc,1974(78):3592366.
[14]HorsburghBC,BrierleyI,BrownTD.
Analysisofa9.
6kbsequencefromthe3′endofcaninecoronavirusgenomicRNA[J].
JGenVirol,1992,73(Pt11):284922862.
[15]NguyenVP,HogueBG.
Proteininteractionsduringcorona2virusassembly[J].
JVirol,1997,71(12):927829284.
[16]BaudouxP,CarratC,BesnardeauL,etal.
CoronaviruspseudoparticlesfornedwithrecombinantMandEproteinsin2ducealphainterferonsynthesisbyleukocytes[J].
JVirol,1998,72(11):863628643.
[17]deHaanCA,deWitM,KuoL,etal.
Theglycosylationsta2tusofthemurinehepatitiscoronavirusMproteinaffectstheinterferogeniccapacityofthevirusinvitroanditsabilitytoreplicateintheliverbutnotthebrain[J].
Virology,2003,312:3952406.
762王玉燕,等:犬冠状病毒南京株M基因的同源重组分析1995-2006TsinghuaTongfangOpticalDiscCo.
,Ltd.
Allrightsreserved.
KeyLabofAnimalDieaseDiagnosticandImmunologyMinistryofAgriculture,NanjingAgricUniv,Nanjing210095,ChinaAbstract:TwoMgenesequencesofCaninecoronavirus(CCV),fromawildstrainNJ17fromadiarrheadogfaecalsampleinNanjingCityandCCV1271referencestrain,wereclonedandse2quenced.
Theresultsshowed,CCV1271sharedhighsimilarity(98.
9~99.
5%)toCCVV1,V2andaCCVstrainfromgiantpandaisolatedinChina2003.
ItsuggestedthattheaboveCCVstrainsexistedquasispeciesfromasameancestorstrain.
OnMgene,CCVNJ17shared87.
0~91.
9%identitieswithotherCCVstrainsandtypicalstrainsofTransmissiblegastroenteritisvirus(TGEV)ofswine,andFelinecoronaviruses(FCoVs)intheGenBank.
TheCCVNJ17mightbeaseparatedevolutionarystraininChina.
RecombinationmarksequencesofallthelandedstrainsontheMgeneputativerecombinationregionhada"CTTTAG"nucleotides,meetingto"CTT(A/T)(A/T)G"thatalwaysappearedneartotherecombination"hotspot"inInfectiousbron2chitisvirusRNAgenome.
Thefirst50aminoacidresiduesonNterminalofCCVNJ17MproteinwashighersimilartoFCoV7921683thanTGEV,whilethefollowingaminoacidresiduese2quencewasclosertoTGEVthanFCoV7921683.
Itindicatedthatarecombinationeventhadhap2penonMgeneofNJ17.
StructureandfunctionpredictionrevealedalltheCCVstrains'Mproteinhadasamesecretorysignalpeptide,fourtransmembraneregions,aminoacidandcarboxylicacidterminalinsertingintotheenvelope.
Putativetrimmedfunctionalsiteswerereserved,andstron2gerantigenicitieswerefoundonthetwoterminalsofMprotein.
Withreservedfunctionalsites,theaminoacidterminalresidueswerehighlyvariedanditsactionseemedtoberelatedtoitsstructuredomainsbutitssequence.
Keywords:Caninecoronavirus(CCV);Mprotein;Similarityrecombination摘要:对来自腹泻犬粪样的犬冠状病毒(CCV)南京株NJ17株及参考株1271的M基因进行了克隆、测序,并与GenBank中所有已知CCV毒株及同亚群的猪冠状病毒(TGEV)和猫冠状病毒(FCoV)代表株的M基因进行了同源性比较和系统进化分析,同时对M蛋白的结构和功能进行了预测分析.
结果表明,CCV1271与近年在中国分离到的CCV毒株V1、V2及大熊猫源的毒株具有98.
9%~99.
5%的同源性,说明这些毒株可能是来自同一毒株的准种.
NJ17与其他中国分离株及国外分离株的同源性为87.
0%~91.
9%,显示国内可能存在一个相对独立进化的CCV毒株.
序列比较发现,所有CCV毒株在可能的同源重组"热点"区内都有一个CTTTAG序列,与鸡传染性支气管炎病毒同源重组模板交换位点附近的特征序列相似.
CCVNJ17株M蛋白在N端50氨基酸序列与FCoV7921683同源性高,而在后212氨基酸序列与TGEV同源性高,提示该毒株可能在M基因上曾经发生过不同病毒收稿日期:2004211209,修回日期:20052032153基金项目:上海科技兴农重点公关项目(农科攻字(2003)第D21号);"江苏省高等学校研究生创新计划"资助项目.
作者简介:王玉燕(19762),女,山东省籍,博士生,研究方向为兽医病毒学.
E2mail:yuyanss@163.
com33通讯作者.
陆承平(19452),男,上海省籍,教授,兽医微生物学与免疫学.
Tel:025284396517,E2mail:lucp@njau.
edu.
cn1995-2006TsinghuaTongfangOpticalDiscCo.
,Ltd.
Allrightsreserved.
的同源重组.
CCVM蛋白的结构及功能预测表明,所有毒株都具有分泌型信号肽,有4个螺旋跨膜区,N末端和C末端均位于膜内.
M蛋白的两末端具有较强的抗原性,M蛋白上存在多种功能性氨基酸修饰位点且相对保守.
N末端的氨基酸变异很大,但是功能性修饰位点相对保守,提示N末端的功能可能与构象有关.
关键词:犬冠状病毒;M蛋白;同源重组中图分类号:S852.
65文献标识码:A文章编号:100325125(2005)0320262206犬状病毒(Caninecoronavirus,CCV)是引起犬急性胃肠炎的重要病原,CCV与猪传染性胃肠炎病毒(Transmissiblegastroenteritisvirus,TGEV)、猫的冠状病毒(Felinecoronaviruses,FCoV)、人冠状病毒229E(Humancoronavirus229E,HCoV229E)等同属于冠状病毒第I群[1].
其中,CCV、FCoV、TGEV的在系统发生树上更为接近,组成一个冠状病G121亚群,而猪流行性腹泻病毒(Porcineepidemicdiarrheavirus,PEDV)和HCoV229E组成G122亚群[2].
不同动物来源的冠状病毒之间可发生同源重组[3,4].
Erles[5]等最近报道,证实冠状病毒也是犬传染性呼吸道综合征(caninein2fectiousrespiractrysyndromediseases,CIRD)的重要病原,其S基因与HCoV2OC43和牛冠状病毒(BCoV)同源性最高,可能来源于OC43或BCoV.
在2003年震惊世界的SARSCoV,很可能就是不同宿主来源冠状病毒重组的结果[6].
CCV不断有新的CCV变异株被发现,而且有较广的宿主范围[7~9],对CCV的研究有潜在的公共卫生意义.
CCV是正股单链RNA病毒,基因组长约27~32Kb,其中2/3的基因组编码一个大的聚合酶蛋白(ORF1a、1b),在ORF1b的下游是几个编码蛋白的ORFs,包括S、E、M、N蛋白及一些可能的非结构蛋白[1].
冠状病毒基因组RNA在复制时采用独特的套式转录,基因组的错配率和重组率都很高[3].
研究发现冠状病毒基因组RNA重组现象的发生是普遍且随机的,几乎在每个位点上都可能发生.
但是,重组位点更趋向于集中在一些重组的"热点(hotspot)"区域,而且总有一些特征的核酸序列出现在其附近[10].
例如,对小鼠肝炎冠状病毒(MHV)研究发现,多数重组体病毒RNA的重组位点在前导序列和5核苷酸重复序列"UCUAA"之间[11].
而"CTT(A/T)(A/T)G"总出现在在鸡传染性支气管炎病毒(IBV)同源重组模板交换位点附近[12].
对冠状病毒同源重组热点的研究多集中在聚合酶基因、5′和3′端的非翻译区及变异较大的S基因上.
冠状病毒的M基因相对保守,但是也有研究认为在M基因上存在RNA同源重组的"热点",Herrevegh等研究证实,FCoVⅡ型毒株7921683的M基因327~376碱基之间存在FCoVⅠ和CCV同源模板RNA交换重组位点[4].
作者在此前的研究中发现,在中国犬群中有较高的CCV感染率,而且健康犬、狐、貉携带CCV的比率也很高.
通过比较M基因的保守区发现,来自南京腹泻犬粪样的CCVNJ17与其他毒株的序列同源性较低,可能是一个相对独立的CCV毒株.
本研究用PCR方法克隆了CCVNJ17株和参考株1271株的M基因.
将得到的M基因序列与Gen2BanK中所有CCV毒株的M基因和与CCV同亚群TGEV、FCoV的代表毒株做比较分析,在其中寻找其他冠状病毒RNA同源重组研究中发现的具有模板交换"热点"特征的序列,同时对相应的M蛋白的结构及功能特性进行了预测分析.
1材料与方法1.
1细胞及参考毒株CCV21271参考株及A72细胞(犬纤维肉瘤细胞系)均由德国吉森大学Baljer教授惠赠.
后者用含10%新生犊牛血清的MEM营养液培养,CCV1271增殖维持液为含2%新生犊牛血清的MEM.
1.
2样本采集及处理CCV阳性粪样NJ17于2003年10月采自南京某宠物门诊,用pH7.
40.
01mol/L的PBS进行5倍稀释后,5000r/min4℃离心20min,取上清保存于-20℃待检.
将CCV1271接种A72细胞,至80%以上细胞出现病变后冻融3次收获病毒,-20℃保存备用.
1.
3RT2PCR取200μL粪便上清加1mLTrizol(32ZOL)(MDBio公司产品)按说明书进行操作,提取RNA.
使用RevertAidTMFisrtStrandcDNASynthesisKit(MBI),按操作说明进行cDNA合成.
参照文献[7]发表的引物序列合成扩增M基因全长的引物MF(5′2CCTTGTTTGAACTAAACAAAATGAAG23′),MR(5′2TCCCTGAGAGGCCATTTAGA23′),引物由上海皓嘉公司合成.
反应体系如下:模板DNA5μL,10*PCR缓冲液2.
5μL,25mmol/LMgCl22μL,10mMdNTPs1μL,引362王玉燕,等:犬冠状病毒南京株M基因的同源重组分析1995-2006TsinghuaTongfangOpticalDiscCo.
,Ltd.
Allrightsreserved.
物各0.
5μl(30pmol/μL),exTaqDNA聚合酶(5U/μL,TaKaRa公司)0.
5μL,用ddH2O调整终体积至25μL.
反应条件:94℃30s、55℃30s、72℃1min,扩增35个循环,最后72℃延伸5min.
1.
4测序及分析PCR产物用胶回收试剂盒回收(北京鼎国公司),连接T载体,(TaKaRa公司),转化大肠杆菌DH5α,挑取阳性克隆,由上海基康公司进行测序.
测序结果在GenBanK上进行比对,然后用DNAstarMegAlin软件与GenBank上发表的所有CCVM基因及冠状病毒G121亚群的TGEV和FCV的代表毒株进行序列比对和同源性比较(表1),应用ClustalX和TreeView软件绘制系统进化树.
同时,在序列中寻找可能在同源重组"热点"附近可能出现的特征性序列,然后分段进行比较分析.
在http://www.
predictprotein.
org网站进行相关毒株M蛋白的结构及功能预测分析.
用Sig2nalP3.
0Server(http://www.
cbs.
dtu.
dk)进行信号肽的预测,DNAStarProtean进行蛋白的抗原性分析和表面结构域预测.
表1CCV及相关冠状病毒G121亚群代表毒株背景Table1BordereauofCCVsandotherG121typicalcorona2virusesinthisarticleStrainsGenBankAccessionNO.
Length(bp)GenotypecountryIsolatedtimeCCV1271AY7049163789CCVⅡGermen1971CCVNJ17AY7049173792CCVⅡChina2004CCVV2AY390344792CCVⅡChina2003CCVV1AY390343792CCVⅡChina2003PandaCCVAY436635792CCVⅡChina2003CCVInsavc21D113096789CCVⅡU.
K.
1980sCCVBGF10AY342160789CCVⅡU.
K.
2004CCV259/01AF502583792CCVⅠItaly2002FECV7921683AB086904792FCoVⅡU.
S.
A.
1970sFIPVUCD1AB086902789FCoVⅠU.
S.
A.
1970sTGEVAJ270965789/Spain2000Note:3submittedbyauthors.
CCV:CanineCoronavirus,FECV:felineentericcoronavirus,FIPV:felineinfectiousperitonitisvirus,TGEV:TransmissibleGastroenteritisVirus.
2结果2.
1CCVM基因的核酸及氨基酸序列分析RT2PCR反应可扩出835bp的目的片段,通过测序后在GenBanK中进行比对确认扩增无误,并登录GenBanK获得登录号:CCV1271AY704916,CCVNJ17AY704917.
该序列中包含M基因的完整ORF,其中CCVNJ17的M基因全长为792bp,编码全长263个氨基酸的蛋白,CCV1271的M基因全长为789bp,编码蛋白为262个氨基酸.
用DNAStar软件同源性分析表明,两毒株完整M基因之间的同源性为91.
8%;与其他G121亚群的冠状病毒相比较,CCV1271和CCVNJ17都与CCVV2同源性最高,分别为99.
4%和91.
9%;CCV1271与TGEV的同源性(93.
5%)高于FECV7921683(85.
4%)和FIPVUCD1(81.
6%);CCVNJ17M基因与TEGV的同源性也都高于与FECV7921683和FIPVUCD1,但是都低于CCV1271与这些毒株的同源性(图1).
CCV1271与CCVV1、CCVV2和PandaCCV同源性很高,在系统发生进化树上位于同一分枝上.
而CCVNJ17却与所有已知CCVM基因的同源性都不高,在系统进化树上与TGEV位于一个分枝上.
图1CCVM基因与G121亚群冠状病毒代表株的核苷酸同源性比较Fig.
1NucleotideidentityanddivergenceoftheCCVandotherG121typicalCoronavirusesMgenenucleotidesequences序列比对发现,在CCVM基因的前150bp变异明显,将M基因的核苷酸序列转化成相应的氨基酸序列,应用DNAStar软件分别对全长M蛋白、M蛋白前50氨基酸,和后212个氨基酸的进行同源性分析,并绘制系统进化树(图2,3,4).
无论完整M蛋白氨基酸序列比较还是分段比较,CCV1271都与CCVV1、CCVV2和PandaCCV在同一分枝上,且与TGEV的同源性高与FCV.
而CCVNJ17在完整M蛋白氨基酸序列比较中,与所有毒株的同源性都不高,但是在N端前50氨基酸中,与FECV7921683的同源性(77.
9%)高于TGEV(76.
9%),而在后212氨基酸片段中,与TGEV的同源性(94.
1%)高于FECV7921683(85.
5%),同样的结果也出现在系统进化树上.
寻找冠状病毒同源重组特征序列发现,所有CCV毒株在相应FECV7921683M基因同源重组的可能区域(3272376bp)内的序列都相对保守,而且在该区域内有一段序列"CTTTAG",符合在鸡传染性支气管炎病毒(IBV)重组研究中发现的特征序列"CTT(A/T)(A/T)G",后者总出现在IBV同源重462中国病毒学第20卷1995-2006TsinghuaTongfangOpticalDiscCo.
,Ltd.
Allrightsreserved.
图2CCVM蛋白氨基酸序列与G121亚群冠状病毒代表株的系统发生进化树Fig.
2PhylogenetictreeofCCVandotherG121typicalCoronavirusesMgeneaminoacidresiduesequences图3CCVM蛋白前50aa序列与G121亚群冠状病毒代表株比较的系统发生进化树Fig.
3PhylogenetictreeofCCVandotherG121typicalCoronavirusesMgenefirst50aminoacidresiduesequences图4CCVM蛋白后212aa序列与G121亚群冠状病毒代表株比较的系统发生进化树Fig.
4PhylogenetictreeofCCVandotherG121typicalCoronavirusesMgenelast212aminoacidresiduesequences组模板交换位点附近[12].
2.
2CCVM基因蛋白结构与功能预测SignalP预测表明所有毒株均在第15、16或17、18位氨基酸之间有信号肽的剪切位点,是典型的分泌型信号肽.
SYNOPSIS预测表明,所有M蛋白在结构上相似,分为膜内区、跨膜区和膜外区,均有4个螺旋跨膜区(如CCVNJ17的跨膜区分别位于55272、792103、1162136、2032220氨基酸残基间),N末端和C末端均位于膜内,两个膜外区.
DISULFIND预测分析在CCVNJ17、Insavc21、BGF10、259/01、FECV7921638、FIPVUCD1等毒株的第9(或10)位C与第31(或30)位C之间可能会形成两个二硫键,而TGEV则可能会在第9位和38位、14与40、15与20、30与157位间的C上形成二硫键.
抗原性分析表明各毒株的抗原位点分布较相似,且主要分布在跨膜区之外(图5,只给出CCVNJ17).
PROSITEmotifsearch寻找蛋白质的功能位点,结果显示在M蛋白上存在多种氨基酸修饰位点且相对保守,毒株之间略有差异(表2).
3讨论冠状病毒在动物中广泛分布以及易于变异而受到关注.
国内对CCV的研究较少,尤其是M基因的变异如何未见报道.
而M基因作为冠状病毒可能发生同源重组的区域之一,值得重视.
CCV1-71株由Binn等于1971年在发生腹泻的德国军犬中分离到,是世界上最早分离到的CCV毒株[13].
由同源性及进化关系上可看出,虽然分离的年代间隔较远,但是CCV1-71与近年在中国分离到几个CCV毒株V1、V2及大熊猫源的毒株具有98.
9%~99.
5%的同源性,说明这些毒株可能是来自同一毒株的准种.
同时表明虽然冠状病毒的变异频率很高,但在M基因上却相对保守.
本研究从腹泻病犬粪样中分离到的CCVNJ17在M基因上与CCV1271、Insavc21、BGF10、259/01等毒株及在中国分离到的其他毒株的同源性都不高(87.
0%~91.
9%),说明国内可能存在一个相对独立进化的CCV毒株.
有研究证实,在相对保守的CCVM基因上存在RNA同源重组的"热点"[4].
本研究通过序列比较发现,所有CCV毒株在对应FCoVⅡ型毒株79~83可能的同源重组"热点"区内都有一个"CTT2TAG"序列,与在IBV同源重组位点附近出现的特征序列"CTT(A/T)(A/T)G"相符.
该特征序列与冠状病毒RNA复制时的通用先导序列非常相似,可能会模拟RNA聚合酶的结合位点,导致RNA复制时模板链发生跳跃转移[12].
CCVNJ17株M蛋白在N末端前50氨基酸序列与FECV同源性高,而在后面的序列与TGEV同源性高,在两者之间具有"CTTTAG"序列,提示该毒株可能在M基因上发生过不同病毒的同源重组.
562王玉燕,等:犬冠状病毒南京株M基因的同源重组分析1995-2006TsinghuaTongfangOpticalDiscCo.
,Ltd.
Allrightsreserved.
表2CCVs及相关G121亚群代表毒株M蛋白功能性位点预测结果Table2PrositemotifpredictionofMproteinofCCVsandotherG121typicalCoronaviruses.
StrainsProbablyfunctionalmotifsiteASNPKCPCK2MYRISRYLAMIPROKARRGDASPCCVNJ1735、56、252/32、103、25427、122、173、185221///CCV127132、55、2512924、34、105、253121、172、1842205//CCVinsavc2132、55、2512940、25336、121、172、184220///CCVBGF1025、55、213、251/105、25135、121、172、184////CCV259/0133、56、2523035、41、254122、172、185////CCVV233、56、2523025、35、106、254122、173、1852216//PandaCCV33、56、2523025、35、106、254122、173、1852216//TGEV32、55、2512924、34、253121、172、1842205//FCV792168333、56、213、2513035、106、254122、173、185////FCVUCD135、55、251/40、25333、121、172、184162/4422Note:ASN:N2glycosylationsite,PKC:ProteinkinaseCphosphorylationsite,PCK2:CaseinkinaseIIphosphorylationsite,MYRISRYL:N2myristoylationsite,AMI:Amidationsite,PROKAR:Prokaryoticmembranelipoproteinlipidattachmentsite,RGD:Cellattachmentse2quence,ASP:Eukaryoticandviralaspartylproteasesactivesite.
图5CCVNJ17M蛋白的抗原性及表面结构域预测Fig.
5AntigenicandsurfaceprobabilitypredictionofCCVNJ17同源重组"热点"区对应的氨基酸位于跨膜区,也相对保守,但是其前面的氨基酸变异明显,尤其是N末端前50氨基酸(从信号肽到第一个跨膜区间的氨基酸),但是该区的几个功能相关位点却相对保守(表2),提示功能可能与构象有关而与氨基酸残基的一级结构无关.
Liao等认为,虽然冠状病毒基因组重组频繁,但是在环境的选择压力下只有某些具有优势的重组体被保存下来,大多数的重组体会被逐渐的淘汰[11].
Horsburgh等认为M蛋白上的N末端的变异与逃避宿主免疫系统压力有关[14].
因此对CCVNJ17M蛋白N末端的变异现象值得进一步研究.
M蛋白是冠状病毒组装和出芽所必需的,在病毒粒子的形成及与宿主的相互作用中起重要作用[15~17].
本研究通过预测分析发现,M蛋白的抗原性和功能区主要在两端.
已知M蛋白可诱导宿主产生α2干扰素[16],就TGEV而言,其诱导干扰素的功能与N末端几个糖基化的位点有关.
C末端几个功能性的氨基酸修饰位点在所有冠状病毒株中都存在,且非常保守,可能与诱导中和抗体产生,及补体依赖性的融细胞作用有关[17],其具体功能还需进一步研究.
本研究对CCVM蛋白结构和功能的预测为进一步研究M蛋白在病毒与宿主相互反应中的作用打下了基础.
致谢本研究在上海交通大学兽医生物技术研究所完成,实验中得到该所领导和孙建和博士的大力支持和帮助,在此深表谢意.
参考文献[1]CavanaghD,PrianDA,BVritonP,etal.
Nidovirales:anewordercomprisingcoronaviridaeandarteriviridae[J].
ArchVirol,1997,1452:6292635.
[2]GonzalezJM,PuertasG,CavanaghD,etal.
AcomparativesequenceanalysistorevisethecurrenttaxonomyofthefamilyCoronaviridae[J].
ArchVirol,2003,148(11):220722235.
[3]LaiSMMC,CavanaghD.
Themolevularbiologyofcorona2viruses[J].
AdvVirusRes,1997,48:12100.
[4]HerreveghAA,ISmeenkI,HorzinekMC,etal.
FelinecoronavirustypeIIstrain7921683and7921146originatefromadoublerecombinationbetweenfelinecoronavirustypeIandca2ninecoronavirus[J],JVirol,1998,72:450824514.
[5]ErlesK,ToomeyC,HarrietWB,etal.
Detectionofagroup2coronavirusindogswithcanineinfectiousrespiratorydisease[J].
Virology,2003,310:162223.
662中国病毒学第20卷1995-2006TsinghuaTongfangOpticalDiscCo.
,Ltd.
Allrightsreserved.
[6]StanhopeMJ,BrownJR,Amrine2MadsenH.
Evidencefromtheevolutionaryanylysisofnucleotidesequencesforarecombi2nanthistoryofSARS2CoV[J].
InfectGenetEvol,2004:15219.
[7]PratelliA,MartellaV,PistelloM,etal.
Identificationofcoronavirusesindogsthatsegregateseparatelyfromthecaninecoronavirusgenotype[J].
JVirolMeth,2003,107:2132222.
[8]JoseMS,ScottP,TimothyHM.
MolecularcharacterizationofavirulentcaninecoronavirusBGFstrain[J].
VirusRes,2004,140:27231.
[9]MatthewJH,GavanAH,RobertPM,etal.
IdentificationofcaninecoronavirusstrainsfromfesesbySgenenested2PCRandmolecularcharacterizationofanewAustralliaisolatep[J].
JClinMicrobio1,2001,39:103621041.
[10]BannerLR,LaiMM.
RandomnatureofcoronavirusRNArecombinationintheabsenceofselectionpressure[J].
Virol2ogy,1991,185(1):4412445.
[11]LiaoCL,LaiMM.
RNArecombinationinacoronavirus:recombinationbetweenviralgenomicRNAandtransfectedRNAfragments[J].
JVirol,1992,66:611726124.
[12]WangL,DavidJ,EllenWC.
Evidenceofnaturalrecombi2nationwithinS1geneofinfectiousbronchitisvirus[J].
Vi2rology,1993,192:7102716.
[13]BinnLN,LazarEC,KeenanKP,etal.
Recoveryandchar2acterizationofacoronavirusfrommilitarydogswithdiarrhea[J].
ProcAnnuMeetUSAnimHealthAssoc,1974(78):3592366.
[14]HorsburghBC,BrierleyI,BrownTD.
Analysisofa9.
6kbsequencefromthe3′endofcaninecoronavirusgenomicRNA[J].
JGenVirol,1992,73(Pt11):284922862.
[15]NguyenVP,HogueBG.
Proteininteractionsduringcorona2virusassembly[J].
JVirol,1997,71(12):927829284.
[16]BaudouxP,CarratC,BesnardeauL,etal.
CoronaviruspseudoparticlesfornedwithrecombinantMandEproteinsin2ducealphainterferonsynthesisbyleukocytes[J].
JVirol,1998,72(11):863628643.
[17]deHaanCA,deWitM,KuoL,etal.
Theglycosylationsta2tusofthemurinehepatitiscoronavirusMproteinaffectstheinterferogeniccapacityofthevirusinvitroanditsabilitytoreplicateintheliverbutnotthebrain[J].
Virology,2003,312:3952406.
762王玉燕,等:犬冠状病毒南京株M基因的同源重组分析1995-2006TsinghuaTongfangOpticalDiscCo.
,Ltd.
Allrightsreserved.
RepriseHosting:$27.97/月-L5640,16G内存,1TB硬盘,10TB月流量,西雅图机房
RepriseHosting是成立于2012年的国外主机商,提供独立服务器租用和VPS主机等产品,数据中心在美国西雅图和拉斯维加斯机房。商家提供的独立服务器以较低的价格为主,目前针对西雅图机房部分独立服务器提供的优惠仍然有效,除了价格折扣外,还免费升级内存和带宽,商家支持使用支付宝或者PayPal、信用卡等付款方式。配置一 $27.97/月CPU:Intel Xeon L5640内存:16GB(原...
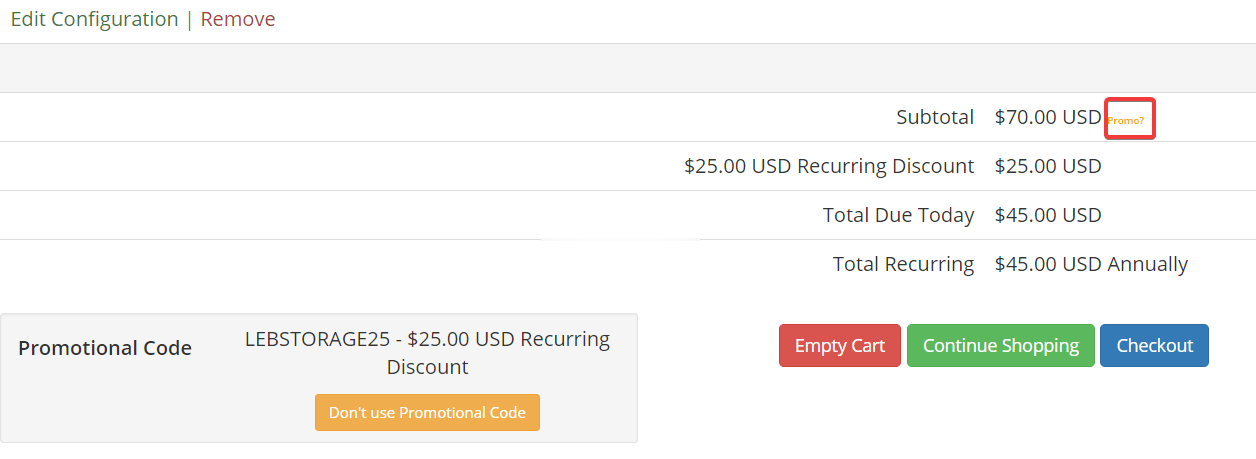
VirMach(8元/月)KVM VPS,北美、欧洲
VirMach,成立于2014年的美国IDC商家,知名的低价便宜VPS销售商,支持支付宝、微信、PayPal等方式付款购买,主打美国、欧洲暑假中心产品,拥有包括洛杉矶、西雅图、圣何塞、凤凰城在内的11个数据中心可以选择,可以自由搭配1Gbps、2Gbps、10Gbps带宽端口,有Voxility DDoS高防IP可以选择(500Gbps以上的防御能力),并且支持在控制面板付费切换机房和更换IP(带...
DiyVM:499元/月香港沙田服务器,L5630*2/16G内存/120G SSD硬盘/5M CN2线路
DiyVM是一家成立于2009年的国人主机商,提供的产品包括VPS主机、独立服务器租用等,产品数据中心包括中国香港、日本大阪和美国洛杉矶等,其中VPS主机基于XEN架构,支持异地备份与自定义镜像,VPS和独立服务器均可提供内网IP功能。商家VPS主机均2GB内存起步,三个地区机房可选,使用优惠码后每月69元起;独立服务器开设在香港沙田电信机房,CN2线路,自动化开通上架,最低499元/月起。下面以...
cv狗狗为你推荐
-
Dimensionfastreport2支付apple支持ipad支持ipad您的iphoneiphone连不上wifi苹果手机无法连接wifi是什么原因ipad上网为什么ipad网速特别慢重庆电信宽带管家重庆电信宽带多少钱一个月迅雷快鸟用迅雷快鸟提示:您所在的网络暂不支持迅雷快鸟迅雷雷鸟迅雷会员每日免费抽奖,抽中迅雷的雷鸟披肩了,要钱吗