Pinusrespring
respring 时间:2021-04-16 阅读:()
Chaparralshrubrecoveryafterfuelreduction:acomparisonofprescribedreandmasticationtechniquesJenniferB.
PottsEvaMarinoScottL.
StephensReceived:1August2009/Accepted:7March2010/Publishedonline:23March2010TheAuthor(s)2010.
ThisarticleispublishedwithopenaccessatSpringerlink.
comAbstractFuelmanagementtechniquesarecom-monlyusedinshrublandstoreducewildrerisk.
However,moreinformationabouttheecologicaleffectsofthesetreatmentsisneededbymanagersandecologists.
Inanefforttoaddressthisneed,weperformedareplicated(4replicatespertreatment)48-haexperimentinnorthernCaliforniachaparraldominatedbyAdenostomafasciculatumtodeterminetheeffectsoftwofuelreductiontypes(prescribedreandmastication)andthreedifferentseasonsoftreat-ment(fall,winter,andspring)onshrubcover,height,andseedlingdensity.
Exclosures(2.
5m2each)werealsousedtoassessherbivoryeffects.
Bythethirdpost-treatmentyear,prescribedretreatmentshadhighershrubcover(71±2%)thanmastication(43±4%).
Therewasnotreatmenteffectonshrubheight,speciesrichness,orcomposition.
Seedlingdensitywasinitiallyhigherinprescribedretreatments(31±4seed-lingsm-2)thanmastication(3±0seedlingsm-2);however,prescribedretreatmentsexperiencedsubstantialmortality,especiallyspringburning,result-inginlowerdensities3yearsaftertreatments(18±0seedlingsm-2afterfallandwinterrecomparedto2±0seedlingsm-2afterspringre).
A.
fasciculatumremainedthedominantshrubspeciesafterthetreatments,andCeanothuscuneatusrecruit-mentwashigherinfallburning.
Deerherbivoryonlyaffectedshrubheight,especiallyinmasticatedunits,resultinginheightsof55±2cminunexclosedareascomparedto66±4cminsideexclosuresbythethirdpost-treatmentyear.
Overall,ourndingssuggestthatfueltreatmentsplayanimportantroleinshrublandcommunitydynamics,atleastintheshort-term,withimplicationsforre-treatmentfrequency,communitystructure,andwildlifehabitat.
KeywordsPrescribedburningMasticationResproutingFirehazardHerbivoryChamiseAdenostomafasciculatumCeanothuscuneatusIntroductionFuelsreductiontreatmentshavebecomeacommonpracticeforreducingthenegativeimpactsofwildretohumancommunities(GillandStephens2009;Stephensetal.
2009),butunfortunately,managersarelackingspecicdetailsregardingtrade-offsandJ.
B.
PottsE.
MarinoS.
L.
Stephens(&)DivisionofEcosystemScience,DepartmentofEnvironmentalScience,PolicyandManagement,UniversityofCalifornia,137MulfordHall,Berkeley,CA94720-3114,USAe-mail:sstephens@berkeley.
eduE.
MarinoLaboratoriodeIncendiosForestales,CentrodeInvestigacionForestal,InstitutoNacionaldeInvestigacionyTecnologaAgrariayAlimentaria,Ctra.
LaCorunakm7.
5,28040Madrid,Spain123PlantEcol(2010)210:303–315DOI10.
1007/s11258-010-9758-1effectsofdifferingtreatmenttypesandseasonsinshrub-dominatedecosystems.
Toaddressexistingknowledgegaps,weexaminedshrubcommunityresponsetoveofthemostcommonlypracticedfuelreductiontreatmentsinshrublandsecosystems.
Wefocusouranalysisonsproutingshrubspeciesduetotheiruniquelifehistorystrategies(BondandvanWilgen1996)andoverwhelminginuenceonthebroaderchaparralplantcommunity.
Fuelreduction,byitsinherentnatureofbeingadisturbance,willaffecteachplantspeciesdifferentlydependinguponthespecies'abilitytosurviveandregenerate.
Shrublandplantspeciescanbegroupedintothreecategories,orfunctionalgroups,dependingupontheirpost-disturbanceregenerationresponse:obligateseed-ers,obligatesprouters,andfacultativeseeders(Keeley1991;Wells1969).
Therstfunctionalgroup,obligateseeders,primarilyreproducefromalong-livedre-stimulatedsoil-stored(e.
g.
,Ceanothuscuneatus,Arc-tostaphylosmanzanita)orcanopy-stored(e.
g.
Bank-sia)seedbanksandlackanysproutingability.
Duetotheirdependencyonre-relatedseedgerminationcues,veryfewobligateseederseedlingsareexpectedtoemergeaftermechanicaltreatments.
Incontrast,obligatesproutingspecies(thesecondfunctionalgroup)regenerateprimarilybysproutingvigorouslyfromadventitiousbuds(e.
g.
,Arctostaphylosglandul-osa,Quercusberberidifolia).
Thesespeciesareexpectedtoregeneratewellafterbothreandmasti-cationtreatments,butfewerresproutingspeciesmaysproutfollowinghighintensityres,sincehighlevelsofsoilheatingmaydamagemeristematictissues.
Obligatesprouterscanalsoreproducefromseed,butmostspeciesonlyproduceafewseedsthatareintolerantofheatexposure(e.
g.
,Quercusspp.
),andonlyasmallnumberofspecies(e.
g.
,A.
glandulosa)produceseedsthatrequirerecues,suchassmoke,heat,andcharatetostimulategermination(Keeley1987,1991).
Thethirdfunctionalgroup,facultativeseeders,reproducesbothsexuallybyseedandvege-tativelybysprouting(e.
g.
,Adenostomafasciculatum,chamise).
Theseedbankoffacultativeseedersisoftenpolymorphic,containingbothre-stimulatedandre-sensitiveseedstoensuregerminationafterandbetweenreevents(Keeley1991;ZammitandZedler1988),therefore,weexpectthisgrouptohaveastrongrecoveryafterdisturbance.
Seasonaltimingoffueltreatmentsisanimportantmanagementconsiderationduetoitsinuenceonthereproductivesuccessofeachfunctionalgroup.
Forexample,vegetativetissuesvaryintheirsensitivityandregenerationabilityindifferentseasons.
Inawetseasonre,lowreintensitiesmayfailtoreleaseseeddormancyformanyobligateseeders,butvegetativegrowthfromresproutersmaybemoreprolicsincelessbudtissueisdamaged.
Conversely,burningduringactivegrowthcanincreasemortalityduetoincreasedtissuesensitivity(FlorenceandFlorence1988).
Seasonaltimingisalsoanimportantdeterminantofseedsurvival.
Manysoft-coatedseeds(facultativeseeders,obligatesprouters)absorbwaterafterrainfallandbecomemorevulnerabletoheat(LeFerandParker2005).
Warmer,drierseasonprescribedres,incontrast,mayproducehighenoughtemperaturestocrackthehardseedcoatsofmostobligateseeders,butthesehighreintensitiesmayalsocausegreatermortalityforsproutingshrubspeciesandheatsensitiveseeds.
Theseasonoftreatmentwillalsostronglyaffectgerminantsurvival.
Aseedthatbreaksdormancyafterafalltreatmentwillhaveseveralmonthstoestablisharootsystembeforethesummerdrought,whereasaspringgermi-nantmaynothaveadequatetimetodeveloproots,andismoresusceptibletodrought-relatedmortality(FlorenceandFlorence1988).
Prescribedresandmasticationcanalsoinuencenon-nativeplantestablishmentandtheassociatedresourcecompeti-tion(PottsandStephens2009).
Shrubgrowthpatternscanbealteredduetodifferentphysiologicalactivityduringtheburntime(Drewaetal.
2002;RadosevichandConard1980;BondandvanWilgen1996).
Lowerresproutingshrubcoverafterspringtreatmentswouldbeexpectedbecausecarbohydrateandwaterreserveshavealreadybeenpartiallyexpended.
Firetreatmentsnotonlyaffectabovegroundvegetationbutalsocanchangesoilproperties.
Whenorganicmatterisconsumedbyre,plantnutrientsarereleasedandbecomehighlyavailableforplantgrowth(DeBanoetal.
1979).
Soilheatingbyrecanalsochangewaterrepellency(DeBano2000)andtherefore,prescribedburningmayhaveadifferentimpactonseedlingrecruitment.
Communitycompositionafterfueltreatmentsmayaffectfuturerepotential.
AdenostomaisconsideredmoreammablethanCeanothus(Biswell1974).
AshiftincommunitycompositionthatfavorsAdenostomamayultimatelycreateahigherriskreenvironment.
304PlantEcol(2010)210:303–315123Herbivorescanhaveastrongselectivepressureonspeciescompositionandabundanceinshrublands(ManierandHobbs2007),particularlyafterdistur-bancewhenregeneratingplantsareeasilyaccessibleandprovidehighlevelsofnutrition.
Obligateseederspeciesthatdependexclusivelyontheirseedbankforregenerationareparticularlyvulnerabletoseedpredators,whileobligatesproutingspeciescanbeheavilyinuencedbyvegetativebrowsers(O'NeilandParker2005).
Duetotheabundanceoflargeherbivoresinourresearchvicinity,includingblack-taileddeer(Odo-coileushemionus),rabbits(LepuscalifornicusandSylvilagus),androdents(Dipodomysagilis,Pero-myscusspp.
,Neotomaspp.
),thereisthepotentialforapronouncedherbivoreimpactontheregeneratingplantcommunity.
Herbivoresmayhavedifferenteffectsonplantrecoveryineachfueltreatment,sincethetypeandseasonoftreatmentcanresultinauniqueinteractionbetweenplantphenologyandbrowsingintensity.
Forexample,springtreatmentswouldtop-killmanysproutingspeciesatthetimeoflowcarbohydrateandwaterreserves,leavingtheplantwithfewresourcestoregenerateinthecoinci-dentperiodofheaviestbrowsingintensity(MorenoandOechel1991).
Herbivorepreferencesforparticularplantspeciesmayalsoinuencethecharacteroftherecoveringshrubcommunity.
Mills(1986)foundthatmamma-lianherbivoressignicantlydecreasedtheabundanceandsurvivorshipofCeanothusseedlingsafterre.
Intheir3-yearstudy,herbivoreselectivityforCeano-thusincreasedovertimeandgaveAdenostomaacompetitiveadvantage.
Adenostoma,however,mayalsosuccumbtodeerbrowsingwithsharplyreducedresproutingvigor(Davis1967).
Althoughmanyimportantstudieshaveexaminedplantrecoveryafterre,amajorityofresearchlackstreatmentreplication(BeyersandWakeman2000;Dunneetal.
1991),managementappropriatespatialscales(MorenoandOechel1991;Tyler1995),oreldsettings(Keeley1987;LeFerandParker2005).
Manyoftheeldstudieshavebeenperformedopportunis-ticallyafterwildresratherthanreplicatedprescribedres(MorenoandOechel1991;Tyler1995;ZammitandZedler1988).
Furthermore,masticationstudiesarerareinthepeer-reviewedliterature,andcomparative,replicatedstudiesofprescribedreandmasticationhaverarelybeenconductedinthechaparralecosystem.
Thegoalofourstudyistoexaminetheecologicalresponsesofthemostcommonfueltreatmentsinchaparralincluding(a)fallprescribedre,(b)winterprescribedre,(c)springprescribedre,(d)fallmastication(slashing),and(e)springmastication.
Inourresearch,weaddressthreequestions:(1)Doesfueltreatmenttypeand/orseasonaffectsproutingshrubcover,(2)Doesfueltreatmenttypeand/orseasonaffectshrubseedlingsurvival,and(3)Doesherbivoryaffectsproutingandseedingspeciesrecoveryaftertreat-ments.
Informationfromthisstudyshouldbeusefultomanagersandecologistsinterestedinshrublandeco-systemsinMediterraneanclimates.
MethodsStudysiteWeconductedourstudyinnorthernCalifornia'sCoastRangechaparral,approximately50kminlandfromthePacicOceanand175kmnorthofSanFrancisco,CA(39°N,123°W).
StudysitesarerepresentativeofCalifornianAdenostomafascicula-tum(chamise)chaparral,withchamisebeingthedominantspecies([65%oftheoverstorycover),andCeanothuscuneatus(buckbrush)andArctostaphylos(manzanita)speciesoccupyingmoreminoroverstorycomponents.
Pre-treatmentvegetationaveraged1–2minheightandnearly100%shrubcover.
Nearbyvegetationtypesincludemixedoakwoodlandonmesic,north-facingslopesandknobconepine(Pinusattenuata)standsonridgetopsandeast-facingslopes.
Chamise-dominatedchaparraloccupieslargeareasinthenortherncoastalrangesofCalifornia.
TheresearchareaexperiencesatypicalMediter-raneanclimatewithhot,drysummersandcool,wetwinters.
Maximummeantemperaturesvarywidelythroughouttheyear,averaging34°Cinsummerand7°Cinwinter.
The30-yearrainfallaverageis100cm,withnearlyallprecipitationoccurringbetweenOctoberandMay.
Annualrainfallduringour2001–2005studyaveraged99.
0cmandrangedfrom90.
7to124.
5cm.
Treatmentareasarelocated700–1,000mabovesealevelonsouthandwestaspectswithslopesof25–55%.
Soilsarederivedfromweatheredsandstoneandshaleandareclassiedasshallow,rocky,andmoderatelyacidic.
Fire-returnintervalsfortheseecosystemsbeforeresuppressionPlantEcol(2010)210:303–315305123wereapproximately30–70years(Stephensetal.
2007).
Fireandothermajordisturbanceswereabsentfromthestudyareaforatleast40yearspriortotreatment.
ExperimentaldesignEachofthevefuelreductiontreatmentswasreplicatedfourtimes;inaddition,fourcontrolplotswerealsoinstalledinthestudyarea(totalof24experimentalunits).
Eachexperimentalunitwasapproximately2ha,andunitsweredistributedacrossa\2kmdistance.
Treatmenttype(prescribedreormastication)andseason(fall/winter/spring)wererandomlyassignedtotheexperimentalunits,althoughmasticationwasrestrictedtositeswithslightlymoregradualslopesduetolimitedequipmentmaneuverabilityinsteepterrain.
Treatmentseasonsweredenedas:Fall(3–20November),Winter(8–18January),andSpring(31March–3Aprilforpre-scribedre,and23April–2Juneforspringmastica-tion).
Springmasticationtreatmentswereconductedslightlylaterthanspringretreatmentsbecauseroadswerenotdryenoughforthemasticatortoaccess.
Wedidnotexpectthisdiscrepancyinspringtreatmenttimingtosignicantlyaffectourndings,sincepreviousanalysisshowedthattreatmenttype(pre-scribedreormastication)overshadowedtheeffectoftreatmentseason(PottsandStephens2009).
Weperformedfuelreductiontreatmentsbetween2001and2003attheUniversityofCaliforniaHoplandResearchandExtensionCenterandadjacentUSBureauofLandManagementSouthCowMoun-tainRecreationArea.
Ourinitialgoalwastocompleteallfourreplicatesofeachtreatmentwithinthesameyear,butunfavorableweatherconditionsandequip-mentavailabilityforcedustosplitreplicatesover2years.
Prescribedreswereignitedwithdriptorchestoproduceupslopeheadres(Stephensetal.
2008).
Masticationwasperformedbyatrackbulldozerwithafrontmountedrotatingtootheddrumwhichshred-dedabovegroundbiomassintoapatchylayer(\5cmdeep)ofwoodydebristhatremainedonthesoilsurface.
Particlesizerangedfrom10to40cminlengthby5–10cminwidth.
Sub-surfacesoilandrootsystemswerenotdisturbedbythemasticator.
Treat-mentheterogeneitywasminimalinmasticationtreatments,sincetheequipmentoperatorperformedsystematicpassesthroughthevegetation.
Vegetationmortalitywas90–100%inbothreandmasticationtreatments.
Wedidnotimplementawintermastica-tiontreatmentduetounpredictableroadaccessandpotentialsoildamagecausedbytheheavymachineryinwetconditions.
MonitoringWeinstalled15permanentlinetransectsineachexperimentalunitpriortoreormasticationtomeasureshrubcoverusingthelineinterceptmethod.
Transectswere15minlengthandrandomlyplacedthroughouttheexperimentalunit;coverwasmeasuredbyplacingatapealongthetransectandcontinuouslyrecordingcoverbyspeciesorbareground.
Shrubheight(cm)wasmeasuredat5mintervalsalongeachlinetransect.
Datafromthe15linetransectswerepooledtorepresenttheexperimentalunit.
Attheendpointsof10randomlyselectedtransects,weestablishedpermanent2.
5m2vegetationsamplingquadratstodetermineseedlingabundanceandcom-position.
Aswiththetransectdata,wepooledseedlingdatafromthe10quadratstorepresenttheexperimentalunit.
Wemonitoredshrubandseedlingdatapriortotreatmentandagainduringthesecondandthirdsummersaftertreatment.
Controlplots(4replicates)weremonitoredbeforetreatmentswereinstalledtodetermineabaselinerangeofvariabilityamongtreatmentunits.
Asubsetofcontrolplotswasmoni-toredeachyearandshowednosignicantchangeinheight,cover,orcomposition.
Thisndingissimilarinotherstudiesthatalsoshowverylittlechangeinmaturechaparralstandsoverthestudyperiodintermsofcoverandheight(Hanes1971).
Immediatelyafterthefueltreatment,werandomlyselectedveofthe10seedlingquadratsforherbivoreexclosures.
Weconstructedexclosuresaroundthesequadrats,includinga20cmbufferzone,foratotalexclosedareaof12.
5m2foreachexperimentalunit.
Exclosureswerebuiltwith1.
5mtallwiremeshthatwascomposedof1.
27cmsquares.
Wesupportedthemeshwithmetalt-poststhatweredrivenintothesoil.
Wedidnotaddroofcoveringsbecausewedidnotexpectherbivorestoclimborjumpoverthemeshsides,andwewantedtominimizeshading,moistureaccumulationandotherindirectexclosureeffects.
Exclosureswereinitiallydesignedtoeliminatedeer,rabbit,androdentbrowsing,buterosioncreatedgaps306PlantEcol(2010)210:303–315123beneaththewiremeshthatwerelargeenoughforsmallanimalentry.
Althoughwebelievethattheexclosuresdeterredmostsmallmammals,wecanonlybecondentthattheexclosurecompletelyexcludedblack-taileddeer.
StatisticalanalysisOne-wayanalysisofvariance(ANOVA)wasusedtocheckfordifferencesbetweencontrolandmanipulatedplotsbeforetreatments(n=4).
FactorialANOVAwasusedtoassesstreatmenttypeandherbivoryeffectonshrubheight,cover,andseedlingabundanceaftertreatments.
IntheseANOVAtests,thetreatmentfactorhadvelevels,oneforeachtreatmenttype(FF=refall,FS=respring,FW=rewinter,MF=mas-ticationfall,MS=masticationspring);andtheex-closurefactorhadtwolevels(exclosedandunexclosed).
ThisfactorialANOVAhastheadvantagetobeabletoidentifypotentialstatisticalinteractionsbetweenfactorsaffectingshrubresponse.
Speciesrichnessandcompositionwereonlyassessedintheunexclosedareasbecauseexclosuresizewasnotlargeenoughtoaddressspeciesdiversity.
Wealsoper-formedrepeatedmeasuresANOVAtoaddresstreat-menteffect(betweengroupsfactor)onshrubcoverovertime(withinsubjectfactor)intheunexclosedareas.
Wepresentdatafromthesecondandthirdpost-treatmentyears,andexcludedatafromtherstpost-treatmentyearbecausespringtreatmentsdidnothavetimetoproducenewgrowthbeforethesummermonitoringperiod.
Weappliedpower,logarithm,andangular(arcsinesquareroot)transformationstotheseedlingdatatomeetANOVAassumptions.
Weusedanon-parametrictest(Kruskal–Wallis)whendatatrans-formationwasnotsufcienttomeetANOVAassump-tions.
TukeyHSDtestswereperformedtoidentifydifferencesbetweentreatmenttypeswhenANOVAresultsdemonstratedsignicanteffects(P\0.
05).
StatisticalanalysesweredonewiththeRstatisticalsoftwarepackage(http://www.
r-project.
org/).
ResultsShrubcoverandheightControlplotshadnosignicantdifferencesinshrubcover(F5,18=1.
55,P=0.
224)orheight(F5,18=0.
39,P=0.
846)comparedtoexperimentalplotsbeforetreatmentswereapplied.
Exclosedareasonlyhadsignicantlytallershrubsthanunexclosedareasonthethirdpost-treatmentyear(F1,30=1.
16,P=0.
290forthesecondyearandF1,30=5.
27,P=0.
028forthethirdyear;Fig.
1).
Treatmenttypehadnosignicanteffectonshrubheight(F4,30=1.
99,P=0.
122andF4,30=1.
45,P=0.
242forthesecondandthirdyear,respectively),andnointeractionbetweenfactorswasobserved.
Ingeneral,retreat-mentstendedtohavetallershrubsthanmasticationtreatmentsoutsidetheexclosuresinthethirdyear.
Regardingshrubcover,therewasnosignicantdifferencebetweenexclosedandunexclosedareasforbothpost-treatmentyears(F1,30=0.
28,P=0.
601forthesecondyearandF1,30=0.
82,P=0.
371forthethirdyear).
However,shrubcoverwassigni-cantlydifferentbytreatmenttype,withP\0.
0001inbothyears(F4,30=9.
90forthesecondyearandF4,30=11.
64forthethirdyear),andnosignicantinteractionbetweentreatmenttypeandexclosure.
Therefore,tobetteraddresscoverdifferencesbetweentreatments,wedecidedtoexcludedatafromtheexclosuresinthefollowinganalyses.
RepeatedmeasuresANOVAintheunexclosedareasconrmedahighlysignicanteffectoftreatmenttypeonshrubcover(F4,45=29.
76,P\0.
0001;Fig.
2).
Ingeneral,retreatmentshadhighercover010203040506070802yr-post3yr-postShrubheight(cm)unexclosedexclosedaaabFig.
1Averageshrubheight(±1SE)forexclosedandunexclosedareasandpost-treatmentyear(secondyearpost-treatmentandthirdyearpost-treatment).
Differentlettersdepictsignicantdifferencesinheightbetweentreatmentswithineachyearforasignicancelevela=0.
05PlantEcol(2010)210:303–315307123valuesthanmasticationtreatments,exceptforthesecondyearwheremechanicalspringandfallrewerenotsignicantlydifferentfromeachother.
Prescribedreplotshadashrubcoverrangingfrom59to72%bythesecondyearand67–76%bythethirdyear,comparedtomasticationplotcoverof29–42%and39–48%,inthesecondandthirdyears,respectively.
Treatmentseasondidnotaffectshrubrecovery.
Thechangeinshrubcoverovertime(withinsubjectfactor)washighlysignicant(F2,45=244.
98,P\0.
0001).
Shrubcoverreductionforbothpre-scribedreandmasticationwassignicant,withcovervaluesremainingsignicantlylowerthanpre-treat-mentvalues3yearsaftertreatmentswereapplied.
However,coverincreasebetweenthesecondandthethirdyearwasnotsignicantwithinanytreatmenttype,althoughMFtendedtohaveaslightlyhigherrecoveryrate(TukeyHSD,P=0.
064).
SpeciesrichnessandcompositionSpeciescompositionandrichnesswereassessedoutsideexclosures.
Priortotreatments,wefound14shrubspeciesalongthecovertransects,withanaverageof6±1speciesperexperimentalunit.
Adenostomafasciculatumwastheoverwhelminglydominantspecies(76%averagecover;Table1),followedbyCeanothuscuneatus(11%),Arctostaph-ylosspecies(6%),Heteromelesarbutifolia(1%),andQuercusspecies(\1%).
Aftertreatments,weidenti-ed16shrubspeciesalongthecovertransects.
Speciesrichnessdidnotdifferbetweentreatments(F4,15=1.
52,P=0.
247forthesecondyearandKruskal–Wallisv2=8.
03,P=0.
090,forthethirdyear)withanaverageof4±1speciesinthesecondyearand5±1speciesinthethirdyear.
Adenostomawastheprimaryspecies3yearsaftertreatments,withsimilarrelativecoverbetweentreatments(F4,15=0.
45,P=0.
774)rangingfrom91to95%,butdifferenttotalcoverbetweentreatments(F4,15=17.
86,P\0.
0001),withretreatmentsshowingsignicanthighervaluesofthisspeciescomparedtomastication.
Theremaining15speciesoccupied\1%covereachbythethirdpost-treatmentyear.
Ceanothuscuneatusaveragedonly0.
9%totalcoverandupto2%relativecoverafter3yearsandneverexceeded2.
5%totalcoveror6.
5%relativecoverinanyexperimentalunit.
ThefallretreatmenthadsignicantlyhigherCeanothuscuneatustotalandrelativecovercomparedtothespringandwinterre0102030405060708090100pre-treat2yr-post3yr-postShrubcover(%)FFFSFWMFMSaabbcccaabbbFig.
2Averageshrubcover(±1SE)foreachtreatment(FFrefall,FSrespring,FWrewinter,MFmasticationfall,MSmasticationspring)andmonitoringtime(pre-treatment,secondyearpost-treatmentandthirdyearpost-treatment).
Differentlettersdepictsignicantdifferencesincoverbetweentreat-mentswithineachyearforasignicancelevela=0.
05Table1Average±SEshrubcover(%)byspeciesforeachtreatmenttype(refall,respring,rewinter,masticationfall,andmasticationspring)before(pre-treatment)and3yearsaftertreatments(3yr-post)innorthernCaliforniachaparralSpeciesYearFireMasticationFallSpringWinterFallSpringAdenostomafasciculatumPre-treatment75.
1±4.
578.
1±3.
375.
1±6.
380.
7±2.
073.
9±4.
43yr-post63.
3±4.
072.
0±1.
365.
0±2.
336.
5±3.
343.
3±5.
6CeanothuscuneatusPre-treatment16.
2±2.
815.
5±3.
58.
4±4.
59.
0±2.
39.
9±3.
83yr-post0.
9±0.
30.
0±0.
00.
0±0.
00.
4±0.
20.
5±0.
2Othershrubspp.
Pre-treatment5.
6±2.
34.
4±2.
912.
8±4.
56.
6±2.
010.
1±1.
53yr-post3.
1±1.
54.
3±2.
35.
0±1.
11.
7±0.
93.
8±1.
5308PlantEcol(2010)210:303–315123treatmentsbythethirdpost-treatmentyear(F4,15=8.
25,P\0.
001intotalcoverandF4,15=8.
96,P\0.
001inrelativecover).
SeedlingestablishmentTotalshrubseedlingdensitywassignicantlydiffer-entbetweentreatmentsinboththesecondandthirdpost-treatmentyears(F4,30=29.
28,P\0.
0001forthesecondyearandF4,30=26.
15,P\0.
0001forthethirdyear),butdidnotdifferbetweenexclosedandunexclosedareas(F1,30=0.
03,P=0.
862forthesecondyear,andF1,30=0.
58,P=0.
449,forthethirdyear).
Inthesecondpost-treatmentyear,allretreatmentshadconsiderablyhigherseedlingdensitiesthanthemasticationtreatments,withanaverageof31±4seedlingsm-2(mean±standarderror)intheretreatmentscomparedto3±0seedlingsm-2inthemasticatedareas.
Bythethirdpost-treatmentyear,thespringretreatmenthadexperienceddispropor-tionatelyhighermortalityandexhibitedseedlingdensitiesthatweremuchmoresimilartothemasti-cationtreatmentsthantotheotherretreatments.
Attheendofthethirdpost-treatmentyear,thefallandwinterprescribedretreatmentshadsignicantlyhigherseedlingdensities,with19±3and18±4seedlingsm-2,respectively,comparedto3±1seedlingsm-2inthespringretreatmentand2±0seedlingsm-2inthemasticatedareas.
Adenostomawasthemostcommonshrubseed-linginallexperimentalunits(Fig.
3).
Adenostomaseedlingdensitysignicantlydifferedbetweentreatmentsinbothpost-treatmentyears(F4,15=9.
16,P\0.
001forthesecondyearandF4,15=15.
10,P\0.
0001forthethirdyear).
Inthesecondpost-treatmentyear,Adenostomaseed-lingdensitywasconsiderablyhigherintheretreatmentsthaninmasticationtreatments,althoughaverageseedlingdensityforthespringburnunitswaslargelyboostedbyasingletreatmentreplicatewithhighseedlingnumbers.
Bythethirdpost-treatmentyear,highseedlingmortalityinspringburnunitsreducedAdenostomaseedlingstosimilardensitiesasinmasticationtreatments.
FallandwinterretreatmentsstillhadhigherAdenostomaseedlingabundancecomparedtotheothertreatments.
Ceanothuscuneatuswasthesecondmostabundantplantspeciesinthepost-treatmentunits.
Thisspeciesalsoshowedsignicantlydifferentseedlingdensitiesbetweentreatmentsinboththesecondandthirdpost-treatmentyears(F4,15=7.
59,P=0.
001andF4,15=3.
09,P=0.
048,respectively;Fig.
3).
Inthesecondpost-treatmentyear,prescribedrehadhigherCeanothuscuneatusseedlingdensitythanmastication,butonlyresultsfromMFweresignicantlylower.
AlthoughMStendedtohavehigherseedlingdensitythanMF,therewasnosignicanteffectofseasoninmasticationtreatments.
Bythethirdtreatmentyear,alltreatmentshadsimilarseedlingdensity,exceptFFthathadsignicantlyhigherdensityofCeanothuscuneatusseedlingsthanFS.
DiscussionTreatmenttypeSignicantdifferencesinchaparralrecoverybetweentreatmentswereobserved,inbothtermsofresprout-ingandseedgermination.
Ingeneral,prescribedretreatmentsresultedinhighershrubcoverandseedlingdensitythanmasticationtreatments3yearsaftertreatments.
Highershrubcoverafterburningcom-paredtomechanicalremovalwasalsoobservedinotherstudiesindifferentshrublandecosystemsincludingshrublandsinFlorida(FitzgeraldandTanner1992),QuercuscocciferagarrigueinFrance(Godronetal.
1981),andMediterraneanheathland(Calvoetal.
1998,2002)andCistuslaurifoliusshrublandinSpain(Tarregaetal.
1997).
Conversely,shrubheightandspeciesrichnesswasnotsigni-cantlydifferentbetweentreatmentsinthisstudy.
Thelackofshrubheightdifferencebetweentreatmentsmaysuggestthatwoodyspeciesrecoveryduringtherstyearafteradisturbanceismainlyinvestedinreoccupyingthebaresurroundingspace,intheabsenceofcompetitionforlight(Marcosetal.
2004).
Calvoetal.
(1998)observedthatthemostpronouncedincreaseinheightoccurredoncetheshrubshaveoccupiedpracticallyalltheirinitialspace,achievingsimilarheightvalues9yearsaftertreatments.
Bytheendofourstudy(3yearsaftertreatments)shrubcoverwasapproximately70%inretreatmentsand40%inmasticatedareas,whichmayexplaintheabsenceofdifferentheightresponseamongtreatments.
Recoverytoinitialshrubcovervaluesoccurred4yearsafterburninginCaliforniaPlantEcol(2010)210:303–315309123chaparral(Dunneetal.
1991)ormechanicalandretreatmentsinMediterraneanheathland(Calvoetal.
1998)andgarrigue(Godronetal.
1981).
GratiniandAmadori(1991)reportedarapidresproutingofmaquisduringtherst4–5yearsafterawildreinsouthwesternItaly,withvegetationstructureandheightsimilartothoseofinitialcommunity8yearsafterre.
Shrubgerminationwasstimulatedintheretreatmentunits,resultinginhigherseedlingdensitiescomparedtomasticatedunits.
SimilarresultswerereportedbyBaezaandRoy(2008)inaMediterraneangorseshrublandinsoutheasternSpain.
Althoughsomeauthorsreportedthatincreasedsoilheatingbyinsolationaftershrubremovalwasenoughtostim-ulateseedgerminationinchaparral(ChristensenandMuller1975),weexpectedlowerseedlingdensitiesinthemasticationtreatmentsbecausethelackofrestimuliwoulddecreasethegerminationofspeciesthatrequireheat,char,orsmoketoreleasedormancy(Keeley1991).
Despitechaparralshrub-seedlingdensitybeinghigherinretreatments2yearsaftertreatments,weobservedhighermortalityratesinburnedplots,especiallyinspringres.
Ourstudy'sseedlingabundancepatternsaresimilartotrendsfoundinotherchaparralresearch(BeyersandWakeman2000;Guo2001;ZammitandZedler1988)whereseedlingnumbersareinitiallyveryhighaftertreatmentandthensuffersignicantmortalityinsubsequentyears.
Drasticmortalityratesintheearlyrecoveryyearscanbecausedbyacombinationoffactors,includingcompetition,herbivory,andtheinsufcientrootdevelopmenttohandlesummerdroughtconditions.
Seedbanklosscanlargelybeattributedtoerosion(Keeley1977)andcoincidenceofbaresoilandheavyrainmayincreaseerosion(Fernandezetal.
2008;GillandGroves1981).
Fernandezetal.
(2008)reportedthatfueltreatmentsaffectedtheinitialrunoff,inl-tration,anderosioninagorseshrublandcommunityinnorthwestSpain,especiallyafterprescribedburn-ingcomparedtomastication.
Initiallowerseedlingdensityinmasticationareasmayexplainthenegligiblemortalityobservedduetoincreasedsoilwaterresourcescausedbyfewerplants(MorenoandOechel1991).
However,Holmgrenetal.
(2000)reportedhigherseedlingsurvivalinburnedplotswhencomparedtomechanicallyclearedplotsinChileanshrubland.
Theseauthorsobservedthatearlyseedlingsurvivalwashigherundertheshadeoflargeshrubs.
Initialgerminationandseedlingsurvivalafterfueltreatmentsiscriticalinchaparralasseedlingrecruitmentisrareuntilthenextdisturbanceoccurs(Keeley1987).
Despitethehighseedlingmortalityobservedinretreatments,ourresultsstilldemonstratedhigherseedlingdensitiesinfallandwinterburningscomparedtospringburningandmastication3yearsafterthetreatments.
Adenostomafasciculatum01020304050FFFSFWMFMSbbbaabBBAAACeanothuscuneatus0123452yr-post3yr-post2yr-post3yr-postDensity(seedlingsm-2)Density(seedlingsm-2)FFFSFWMFMSBAABABABbbbaabFig.
3Averageseedlingdensity(±1SE)forAdenostomafasciculatumandCeanothuscuneatusbytreatmenttype(FFrefall,FSrespring,FWrewinter,MFmasticationfall,MSmasticationspring)andpost-treatmentyear.
Differentlettersdepictsignicantlydifferentseedlingdensitybetweentreat-mentsforthesecondyear(lowercases)andthirdyear(uppercase)forasignicancelevela=0.
05310PlantEcol(2010)210:303–315123Greatershrubrecoveryfollowingremaybeexplainedbythelargenutrientreleasethatresultsfrombiomasscombustionandashdeposition(Chris-tensen1973;ChristensenandMuller1975;DeBanoetal.
1979;RundelandParsons1980).
Masticatedsites,incontrast,haveaslownutrientreleaseastheshreddedbiomassdecaysandthismaycontributetotheslowerregrowth.
Eventhoughregenerationmechanismsofchaparralspeciesareconsideredadaptedtore,theresultsofourandotherstudiesinMediterraneanshrublandssuggestthatthesecom-munitieshaveahighresiliencetodisturbance,eithercausedbyreorbymechanicaltreatments(Calvoetal.
1998;DelBarrioetal.
1999;BondandMidgley2001;PottsandStephens2009).
Furthermore,non-redisturbancesaredemonstratedtoplayanimpor-tantroleintheevolutionofsomechaparralcommu-nities(Ackerly2004).
Thepresenceofre-followingspeciesinmasticatedareas(PottsandStephens2009)maybeattributedtopolymorphicseedbanks(Keeley1991;Sweeney1956;ZammitandZedler1994)and/orsufcientlyhighsolarheatingthatovercomesphysicalseeddormancy(BaskinandBaskin1998;ChristensenandMuller1975).
Calvoetal.
(2002)observedthatdifferencesinshrubcoverafterdiffer-entfueltreatments(includingburningandcutting)duringtherstyearofsuccessioninaMediterraneanheathlandtendedtobeeliminatedafter12years,andmostofthespeciestendedtorecovertotheirinitialcovervalues.
Thenumberofshrubspeciesdidnotdifferbetweenreandmechanicaltreatmentsinourstudy.
AsimilarresultwasfoundbyFitzgeraldandTanner(1992)inchaparralandbyCalvoetal.
(2005)inMediterraneanshrublandecosystems.
Regardingspe-ciescomposition,Adenostomaremainedthedomi-nantspeciesafterdisturbance,showinghighertotalcoverafterrethanmechanicaltreatments,butsimilarrelativecoverbetweentreatments.
Thispatternwasexpectedconsideringpre-treatmentAd-enostomadominanceandthespecies'abilitytobothresproutaftertop-killandgerminatefromasoil-storedseedbank(OdionandDavis2000).
Theversatileadaptationoffacultativeseedersmaybeanadvantageforrecoveringinawiderangeofenviron-mentalconditionsoccurringafterdisturbances(SeligmanandHenkin2000).
Non-dominantspecies,suchasCeanothuscunea-tus,haddifferentresponsesthanthedominantspecies.
Ceanothusisofparticularlystronginteresttolandmanagersduetoitsnutritionalvaluetomanybrowsingwildlifespecies(Biswell1974),includingblack-taileddeer.
Bythethirdyearaftertreatments,Ceanothuscuneatustotalcoverwasstilllowbecausethisspeciesonlyregeneratesbygermination,show-ingslightlyhigherrelativecoverinfallburntreat-mentsthaninthespringandwinterburntreatments.
Nevertheless,theslowrecoveryofCeanothusmaynotbedetrimentaltothisspecies.
Onceseedlingsareestablishedandnitrogenavailabilitydecreasesfol-lowingtheinitialushimmediatelypost-re,nitro-gen-xingspecies,suchasCeanothus,shouldhaveanincreasingcompetitiveadvantageovernon-sym-bioticshrubs(EllisandKummerow1988),especiallyinnitrogen-limitedecosystemslikechaparral(RundelandParsons1980).
InAdenostomadominatedchap-arral,Davis(1967)reportedCeanothuscoverincreased10yearsafterreinsouthernCalifornia.
TreatmentseasonSeasonoftreatmentdidnotsignicantlyaffectshrubresproutinginourstudy.
Resultsontreatmentseasoneffectsarevariedintheliterature.
FitzgeraldandTanner(1992)alsofoundnoeffectsofseasononeitherreormechanicaltreatmentsinshrublandsinFlorida.
Theseauthorsarguedthatsoilmoistureaffectspost-treatmentrecoveryandexplainedtheabsenceofdifferenceintheirstudytorelativelylowmoisturelevelsinbothseasonswhenthetreatmentswereapplied(winterandsummer).
Dunneetal.
(1991)alsoreportednonegativeeffectoffallburnsonchaparralrecoveryandGodronetal.
(1981)didnotobserveanydifferenceinvegetationcoverbetweenfallandspringburntreatmentsinFrenchgarrigue.
RadosevichandConard(1980)reportednoeffectonpost-reAdenostomashootgrowthbetweenburningseasonsinnorthernCaliforniachaparral.
Theseauthorsarguedthatpost-regrowthisnotdependentonthewaterstatusoftheshrubbeforetheburningtime,butonsubsequentsuitableenviron-mentalconditions.
Conversely,MorenoandOechel(1991)reportedthatincreasingreintensityincreasedplantmortality,reducedthenumberofresproutsperplant,anddelayedthetimeofresproutinginchaparral.
SimilarresultswerefoundbyLloretandLopez-Soria(1993)intheMediterraneanshrubEricamultiorawhenPlantEcol(2010)210:303–315311123applyinghigherintensityretoindividualplants.
FlorenceandFlorence(1988)alsoreportedincreasedAdenostomamortalityathighreintensitywhenburnswereconductedduringthegrowingorower-ingseason.
However,Canadelletal.
(1991)didnotobservedifferencesinthebiomassofresproutsofindividualshrubsburnedatdifferentreintensitiesinaMediterraneanshrubland.
OurndingsconictwithsomepreviousresearchthatfoundlowershrubsproutingafterspringprescribedreinsouthernCaliforniachaparral(BeyersandWakeman2000).
Althoughseasonoftreatmentdidnothaveastronginuenceonshrubreprouting,seasonoftreatmentdidhaveasignicantinuenceonseedlingdensity.
Fallandwinterretreatmentshadhigherseedlingdensitiesthananyothertreatment.
WeexpectedhighseedlingdensitiesinfallretreatmentsbecausethistreatmentbestapproximatesCaliforniachaparralwildreregimesbeforeresuppression(Stephensetal.
2007).
However,weweresurprisedtoobservesubstantialseedlingrecruitmentafterthewinterretreatments,sincemoistsoilconditionsatthetimeofburninghavebeenshowntohinderseedlingsuccessforsomespecies(LeFerandParker2005).
Labora-torygerminationresearchbyLeferandParker(2005)hasdemonstratedthatAdenostomaseedsaresensitivetoheatundermoistsoilconditionsduetotheamountofwaterabsorbedintotheseedandthelackofahardseedcoat.
Adenostomaseedlingdensitiesdidnotvarybetweenthethreeburningseasons2yearsaftertreatments,andremainednearlyidenticalinthefallandwinterretreatmentsbythethirdyearaftersubsequentmortality.
Masticationtreatmentsalsoshowedsimilarseedlingdensitiesforthisspeciesbetweenfallandspring.
Weexpectedthespringtreatmentstohavethelowestseedlingdensities,sincegerminatingplantshaveaninsufcienttimeandresourcestodevelopdrought-survivingrootsystemspriortothehot,dryMediterraneansummer(MooneyandParsons1973;Bondetal.
1984;SaruwatariandDavis1989).
Ourseedlingmortalitydataconrmthishypothesis.
ThehighestseedlingmortalitywasobservedinspringburningforbothAdenostomaandCeanothusseed-lingsindistinctively(85and87%,respectively).
Fallburning,whererebehaviorandeffectsareexpectedtobemoresimilartowildreinchaparralcommunities,resultedinmorethan50%seedlingmortalityforAdenostomacomparedto\23%forCeanothus.
Winterburningresultedinlowerseed-lingmortalityinAdenostoma(33%)comparedtoCeanothus(72%).
Although,theabilityofaseedtodevelopadequaterootspriortosummerdroughtmaybethestrongestinuenceonseedlingmortality,functionalgroupcharacteristicsmayalsobeanimportantfactorindeterminingseedlingmortalityandsurvival.
Consis-tentwithourndings,otherstudieshavereportedhighermortalityinfacultativeseederspeciescom-paredtoobligateseederspeciesafterwildre(FrazerandDavis1988;ThomasandDavis1989)andfallburning(OdionandDavis2000)insouthernCalifor-niachaparral,suggestingthatthelackofresproutingabilityamongobligateseedersisoffsetbyanenhancedabilitytoestablishseedlingsafterre.
Anotherchaparralstudyreportedthatobligateseed-ersseedlingarebetteradaptedtowaterstress,demonstratinghighersurvivalratesinopengapsandwithoutshadingfromadjacentshrubcanopies(Prattetal.
2008).
Regardingmastication,althoughfallandspringtreatmentshadsimilarseedlingdensities,seasonseemedtoaffectseedlingmortalityinaverydifferentmanner,withfallmasticationevenshowingseedlingrecruitmentbythethirdyearforbothAdenostomaandCeanothus.
Ourresultssuggestthat,intheabsenceofre-enhancinggermination,seedlingestablishmentafterfallmasticationtreatmentsoccurredforatleast3years.
HerbivoryThisstudydemonstratedarelativeeffectofherbi-voresonvegetationregrowthatourtreatmentscale.
Shrubheightwashigherintheexclosedareas3yearsaftertreatment;however,coverwasnotsignicantlyaffected.
Althoughtreatmenttypehadnosignicanteffectonshrubheight,theherbivoryeffectwashigherinmasticationcomparedtoretreatments.
Thiscouldbeexplainedbythepresenceofplantskeletonsafterburning,notpresentaftermastication,whichmayinterferewithlargemammalmovementpreventingaccesstobasalsprouts(Davis1967).
Itisimportanttonotethatourrelativelysmalltwo-hectaretreatmentareasmayhaveleadtointensebrowsingpressureduetotheirplacementinanotherwisedensechaparralmatrix.
Largertreatmentareasmayexperiencelessbrowsingpressure.
312PlantEcol(2010)210:303–315123Ourresultssuggestthatlargeherbivoresdonotextensivelybrowseshrubseedlingsintheearlyyearsfollowingdisturbance,sinceseedlingabundancewassimilarinexclosedandunexclosedareas(atleastatthespatialscalesexaminedhere).
PreviousherbivorystudiesinCaliforniachaparralalsofoundnosignif-icantexclosureeffectsonvegetationrecovery(Tyler1995,1996).
However,theseresultsareincontrasttootherstudies(Mills1983,1986)wheredifferencesinseedlingdensitywerefoundbetweenexclosedandunexclosedareas.
Nevertheless,itshouldbepointedoutthat,inourstudy,theexclosureswereonlyeffectiveagainstlargemammals,i.
e.
,black-taileddeer,andprobablydidnotpreventherbivoryfromsmallermammals,suchasrabbitsormice.
ManagementimplicationsInordertoimproveblack-taileddeerhabitat,pre-scribedrewouldbemostbenecialinfall,sinceitproducessignicantlyhigherCeanothuscuneatusseedlingdensityafterthethirdpost-treatmentyear.
Ingeneral,springretreatmentcouldbedetrimentalduetothelowerseedlingsurvivalratesduetosummerdrought.
Regardingmechanicaltreatments,fallandspringmasticationresultedinsimilarseedlingdensities,althoughfalltreatmentsforbothreandmasticationreducedseedlingmortalityasseedlingshadmoretimetodevelopadequaterootssystemspriortothesummerdrought.
Fromawildlifemanagementstandpoint,retreatmentshavefaster,densershrubregrowth,whichmayencouragewildliferecoverywithinashorttimeafterfueltreatment.
However,regardingwildreprevention,masticationtreatmentsaremoreeffectiveintermsofshrubfuel-loadaccumulationbecausetheyhavedramaticallyslowershrubregrowth,andtherefore,reducepotentialreintensityincaseofawildre.
However,rehazardswouldprobablybehigherinmasticatedareascomparedtothosetreatedwithprescribedreintheshort-termbecausenewlymasticatedareashavenedeadfuelsontheground(Bradleyetal.
2006)andincreasenon-nativegrasscover(PottsandStephens2009).
Inthepast,limitedplantrecoverydatahashinderedfuelsmanagementdecisionsandcausedpotentiallydetrimentalmanagementactions.
Regionaldifferencesinplantresponse,relianceonpost-wildredata(duetoalackofprescribedredata),andthealmostcompleteabsenceofmasticationresearchhavecontributedtotheconfusion.
Despitethiscomplexdecisionenvironment,itisimportantformanagerstomakethemostresponsibledecisionsbasedontheirobjectives.
Thegoalofourresearchwastoprovidemanagersandscientistswithdatatoreducepreviousuncertainty.
Clearly,moreresearchwillbeneededtollintheremaininggapsinknowledgeandtestthendingsofthisstudyinthelong-termandinothergeographicregions.
AcknowledgmentsWethanktheUSJointFireScienceProgram(Project#00-2-02)andtheUSFSRiversideFireLab(DavidWeise)forfundingthisresearch,andourprojectcooperatorsfromtheBureauofLandManagementUkiahFieldOfce(BLM),UniversityofCaliforniaHoplandResearchandExtensionCenter(HREC),andCaliforniaDepartmentofForestryandFireProtectionMendocinoandLakeCountyUnits(CALFIRE).
WeextendspecialthankstoJ.
Dawson(BLM),R.
Keiffer(HREC),T.
Derum(CALFIRE),M.
Tolbert(CALFIRE),andW.
Baxter(CALFIRE).
Wealsoextendourgratitudetonumerouseldassistants,particularlyD.
Fry,whoalsoreviewedthismanuscript.
EvaMarino'sparticipationinthisresearchwaspossiblethroughaPh.
D.
scholarshipfundedbyINIA(InstitutoNacionaldeInvestigacionyTecnologaAgrariayAlimentaria),Spain.
OpenAccessThisarticleisdistributedunderthetermsoftheCreativeCommonsAttributionNoncommercialLicensewhichpermitsanynoncommercialuse,distribution,andreproductioninanymedium,providedtheoriginalauthor(s)andsourcearecredited.
ReferencesAckerlyD(2004)Functionalstrategiesofchaparralshrubsinrelationtoseasonalwaterdecitanddisturbance.
EcolMonogr74:25–44BaezaMJ,RoyJ(2008)Germinationofanobligateseeder(Ulexparviorus)andconsequencesforwildreman-agement.
ForEcolManag256:685–693BaskinCC,BaskinJM(1998)Seeds:ecology,biogeography,andevolutionofdormancyandgermination.
AcademicPress,SanDiegoBeyersJL,WakemanCD(2000)SeasonofburneffectsinSouthernCaliforniachaparral.
In:KeeleyJE,Baer-KeeleyM,FotheringhamCJ(eds)2ndInterfacebetweenEcologyandLandDevelopmentinCalifornia.
U.
S.
GeologicalSurveyOpenFileReport00-62,pp45–55BiswellHH(1974)Effectsofreonchaparral.
In:KozlowskiTT,AhlgrenCE(eds)Fireandecosystems.
AcademicPress,NewYorkBondWJ,MidgleyJJ(2001)Ecologyofsproutinginwoodyplants:thepersistenceniche.
TrendsEcolEvol16:45–51PlantEcol(2010)210:303–315313123BondWJ,vanWilgenBW(1996)FireandPlants.
ChapmanandHall,LondonBondWJ,VlokJ,ViviersM(1984)VariationsinseedlingrecruitmentofCapeProteaceaeafterre.
JEcol72:209–221BradleyT,GibsonJ,BunnW(2006)Fireseverityandintensityduringspringburninginnaturalandmasticatedmixedshrubwoodland.
USDAForestServiceProc.
RMRS-P-41CalvoL,TarregaR,LuisE(1998)Space-timedistributionpatternsofEricaaustralisL.
subsp.
aragonensis(Willk)afterexperimentalburning,cutting,andploughing.
PlantEcol137:1–12CalvoL,TarregaR,DeLuisE(2002)ThedynamicsofMediterraneanshrubsspeciesover12yearsfollowingperturbations.
PlantEcol160:25–42CalvoL,TarregaR,LuisE,ValbuenaL,MarcosE(2005)Recoveryafterexperimentalcuttingandburninginthreeshrubcommunitieswithdifferentdominantspecies.
PlantEcol180:175–185CanadellJ,LloretF,Lopez-SoriaL(1991)ResproutingvigouroftwoMediterraneanshrubspeciesafterexperimentalretreatments.
Vegetatio95:119–126ChristensenNL(1973)FireandthenitrogencycleinCali-forniachaparral.
Science188:66–68ChristensenNL,MullerCH(1975)EffectsofreonfactorscontrollingplantgrowthinAdenostomachaparral.
EcolMonogr45:29–55DavisJ(1967)Someeffectsofdeerbrowsingonchamisesproutsafterre.
AmMidlNat77:234–238DeBanoLF(2000)Theroleofreandsoilheatingonwaterrepellencyinwildlandenvironments:areview.
JHydrol231–232:195–206DeBanoLF,RiceRM,ConradCE(1979)Soilheatinginchaparralres:effectsonsoilproperties,plantnutrients,erosionandrunoff.
USDAForestService,PSW-145DelBarrioJ,Luis-CalabuigE,TarregaR(1999)VegetativeresponseofArctostaphylosuva-ursitoexperimentalcut-tingandburning.
PlantEcol145:191–195DrewaPB,PlattWJ,MoserEB(2002)Fireeffectsonres-proutingofshrubsinheadwatersofsoutheasternlongleafpinesavannas.
Ecology83:755–767DunneJ,DennisA,BartolomeJW(1991)ChaparralresponsetoaprescribedreintheMountHamiltonrange,SantaClaracounty,California.
Madrono38:21–29EllisBA,KummerowJ(1988)N2(C2H2–C2H4)xationintwospeciesofCeanothusseedlingsinsecondyearpostrechaparral.
PlantSoil109:207–213FernandezC,VegaJA,FonturbelT,JimenezE,PerezJR(2008)Immediateeffectsofprescribedburning,choppingandclearingonrunoff,inltrationanderosioninashrublandareainGalicia(NWSpain).
LandDegradDev19:502–515FitzgeraldSM,TannerGW(1992)AviancommunityresponsetoreandmechanicalshrubcontrolinsouthFlorida.
JRangeManag45:369–400FlorenceSR,FlorenceMA(1988)PrescribedburningeffectsinCentralCaliforniachaparral.
Rangelands10:138–140FrazerJM,DavisSD(1988)Differentialsurvivalofchaparralseedlingsduringtherstsummerdroughtafterwildre.
Oecologia76:215–221GillAM,GrovesRH(1981)Fireregimesinheathlandsandtheirplant-ecologicaleffects.
In:SpechtRL(ed)Eco-systemsoftheworld:heathlandandrelatedshrublands.
Elsevier,Amsterdam,pp61–82GillAM,StephensSL(2009)Scienticandsocialchallengesforthemanagementofre-pronewildland–urbaninter-faces.
EnvironResLett4:034014GodronM,GuillermJL,PoissonetJ,PoissonetP,ThiaultM,TrabaudL(1981)Dynamicsandmanagementofvegeta-tion:effectsofreandgrazingonQuercuscocciferagarrigue.
In:DiCastriF,GoodhallDW,SpechtRL(eds)Mediterranean-typeshrublands.
Elsevier,Amsterdam,pp317–347GratiniL,AmadoriM(1991)Post-reresproutingofshrubbyspeciesinMediterraneanmaquis.
Vegetatio96:137–143GuoQ(2001)Earlypost-resuccessioninCaliforniachapar-ral:changesindiversity,density,coverandbiomass.
EcolRes16:471–485HanesTL(1971)SuccessionafterreinthechaparralofsouthernCalifornia.
EcolMonogr41(1):27–42HolmgrenM,SeguraAM,FuentesER(2000)LimitingmechanismsintheregenerationofChileanmatorral.
PlantEcol147:49–57KeeleyJE(1977)Seedproduction,seedpopulationsinsoil,andseedlingproductionafterrefortwocongenericpairsofsproutingandnonsproutingchaparralshrubs.
Ecology58:820–829KeeleyJE(1987)RoleofreinseedgerminationofwoodytaxainCaliforniachaparral.
Ecology68:434–443KeeleyJE(1991)SeedgerminationandlifehistorysyndromesintheCaliforniachaparral.
BotRev57:81–116LeFerD,ParkerVT(2005)Theeffectofseasonalityonburnonseedgerminationinchaparral:theroleofsoilmoisture.
Madrono52:166–174LloretF,Lopez-SoriaL(1993)ResproutingofEricamultioraafterexperimentalretreatments.
JVegSci4:367–374ManierDJ,HobbsNT(2007)Largeherbivoresinsagebrushsteppeecosystems:livestockandwildungulatesinuencestructureandfunction.
Oecologia152:739–750MarcosE,TarregaR,Luis-CalabuigE(2004)InteractionsbetweenMediterraneanshrubspecieseightyearsafterexperimentalre.
PlantEcol170:235–241MillsJN(1983)Herbivoryandseedlingestablishmentinpost-resouthernCaliforniachaparral.
Oecologia60:267–270MillsJN(1986)HerbivoresandearlypostresuccessioninSouthernCaliforniachaparral.
Ecology67:1637–1649MooneyHA,ParsonsDJ(1973)StructureandfunctionoftheCaliforniachaparral:anexamplefromSanDimas.
In:DiCastriF,MooneyHA(eds)Mediterraneantypeecosys-tems.
Springer,NewYork,pp83–112MorenoJM,OechelWC(1991)FireintensityandherbivoryeffectsonpostreresproutingofAdenostomafascicula-tuminsouthernCaliforniachaparral.
Oecologia85:429–433O'NeilSE,ParkerVT(2005)FactorscontributingtothesoilseedbanksizeoftwoobligateseedingCeanothusspeciesinNorthernCalifornia.
Madrono52:182–190OdionDC,DavisFW(2000)Fire,soilheating,andthefor-mationofvegetationpatternsinchaparral.
EcolMonogr70:149–169314PlantEcol(2010)210:303–315123PottsJB,StephensSL(2009)Invasiveandnativeplantresponsestoshrublandfuelreduction:comparingpre-scribedre,mastication,andtreatmentseason.
BiolConserv142:1654–1657PrattRB,JacobsenAL,MohlaR,EwersFW,DavisSD(2008)Linkagebetweenwaterstresstoleranceandlifehistorytypeinseedlingsofninechaparralspecies.
JEcol96:1252–1265RadosevichSR,ConardSG(1980)Physiologicalcontrolofchamiseshootgrowthafterre.
AmJBot67:1442–1447RundelPW,ParsonsDJ(1980)Nutrientchangesintwochaparralshrubsalongare-inducedagegradient.
AmJBot67:51–58SaruwatariMW,DavisSD(1989)Tissuewaterrelationsofthreechaparralshrubspeciesafterwildre.
Oecologia80:303–308SeligmanNG,HenkinZ(2000)RegenerationofadominantMediterraneandwarf-shrubafterre.
JVegSci11:893–902StephensSL,MartinRE,ClintonNE(2007)PrehistoricreareaandemissionsfromCalifornia'sforests,woodlands,shrublandsandgrasslands.
ForEcolManag251:205–216StephensSL,WeiseDR,FryDL,KeifferRJ,DawsonJ,KooE,PottsJ,PagniP(2008)MeasuringtherateofspreadofchaparralprescribedresinnorthernCalifornia.
FireEcol4:74–86StephensSL,MoghaddasJJ,EdiminsterC,FiedlerCE,HasseS,HarringtonM,KeeleyJE,McIverJD,MetlenK,SkinnerCN,YoungbloodA(2009)Firetreatmenteffectsonvegetationstructure,fuels,andpotentialreseverityinwesternU.
S.
forests.
EcolAppl19:305–320SweeneyJR(1956)Responsesofvegetationtore:astudyoftheherbaceousvegetationfollowingchaparralres.
UnivCalifPublBot28:143–249TarregaR,Luis-CalabuigE,AlonsoI(1997)Space-timehet-erogeneityintherecoveryafterexperimentalburningandcuttinginaCistuslaurifoliusshrubland.
PlantEcol129:179–187ThomasCM,DavisSD(1989)Recoverypatternsofthreechaparralshrubspeciesafterwildre.
Oecologia80:309–320TylerCM(1995)Factorscontributingtopostreseedlingestablishmentinchaparral:directandindirecteffectsofre.
JEcol83:1009–1020TylerCM(1996)Relativeimportanceoffactorscontributingtopostreseedlingestablishmentinmaritimechaparral.
Ecology77:2182–2195WellsPV(1969)TherelationbetweenmodeofreproductionandextentofspeciationinwoodygeneraoftheCaliforniachaparral.
Evolution23:264–267ZammitCA,ZedlerPH(1988)Theinuenceofdominantshrubs,reandtimesincereonsoilseedbanksinmixedchaparral.
Vegetatio75:175–187ZammitCA,ZedlerPH(1994)Organizationofthesoilseedbankinmixedchaparral.
Vegetatio111:1–16PlantEcol(2010)210:303–315315123
PottsEvaMarinoScottL.
StephensReceived:1August2009/Accepted:7March2010/Publishedonline:23March2010TheAuthor(s)2010.
ThisarticleispublishedwithopenaccessatSpringerlink.
comAbstractFuelmanagementtechniquesarecom-monlyusedinshrublandstoreducewildrerisk.
However,moreinformationabouttheecologicaleffectsofthesetreatmentsisneededbymanagersandecologists.
Inanefforttoaddressthisneed,weperformedareplicated(4replicatespertreatment)48-haexperimentinnorthernCaliforniachaparraldominatedbyAdenostomafasciculatumtodeterminetheeffectsoftwofuelreductiontypes(prescribedreandmastication)andthreedifferentseasonsoftreat-ment(fall,winter,andspring)onshrubcover,height,andseedlingdensity.
Exclosures(2.
5m2each)werealsousedtoassessherbivoryeffects.
Bythethirdpost-treatmentyear,prescribedretreatmentshadhighershrubcover(71±2%)thanmastication(43±4%).
Therewasnotreatmenteffectonshrubheight,speciesrichness,orcomposition.
Seedlingdensitywasinitiallyhigherinprescribedretreatments(31±4seed-lingsm-2)thanmastication(3±0seedlingsm-2);however,prescribedretreatmentsexperiencedsubstantialmortality,especiallyspringburning,result-inginlowerdensities3yearsaftertreatments(18±0seedlingsm-2afterfallandwinterrecomparedto2±0seedlingsm-2afterspringre).
A.
fasciculatumremainedthedominantshrubspeciesafterthetreatments,andCeanothuscuneatusrecruit-mentwashigherinfallburning.
Deerherbivoryonlyaffectedshrubheight,especiallyinmasticatedunits,resultinginheightsof55±2cminunexclosedareascomparedto66±4cminsideexclosuresbythethirdpost-treatmentyear.
Overall,ourndingssuggestthatfueltreatmentsplayanimportantroleinshrublandcommunitydynamics,atleastintheshort-term,withimplicationsforre-treatmentfrequency,communitystructure,andwildlifehabitat.
KeywordsPrescribedburningMasticationResproutingFirehazardHerbivoryChamiseAdenostomafasciculatumCeanothuscuneatusIntroductionFuelsreductiontreatmentshavebecomeacommonpracticeforreducingthenegativeimpactsofwildretohumancommunities(GillandStephens2009;Stephensetal.
2009),butunfortunately,managersarelackingspecicdetailsregardingtrade-offsandJ.
B.
PottsE.
MarinoS.
L.
Stephens(&)DivisionofEcosystemScience,DepartmentofEnvironmentalScience,PolicyandManagement,UniversityofCalifornia,137MulfordHall,Berkeley,CA94720-3114,USAe-mail:sstephens@berkeley.
eduE.
MarinoLaboratoriodeIncendiosForestales,CentrodeInvestigacionForestal,InstitutoNacionaldeInvestigacionyTecnologaAgrariayAlimentaria,Ctra.
LaCorunakm7.
5,28040Madrid,Spain123PlantEcol(2010)210:303–315DOI10.
1007/s11258-010-9758-1effectsofdifferingtreatmenttypesandseasonsinshrub-dominatedecosystems.
Toaddressexistingknowledgegaps,weexaminedshrubcommunityresponsetoveofthemostcommonlypracticedfuelreductiontreatmentsinshrublandsecosystems.
Wefocusouranalysisonsproutingshrubspeciesduetotheiruniquelifehistorystrategies(BondandvanWilgen1996)andoverwhelminginuenceonthebroaderchaparralplantcommunity.
Fuelreduction,byitsinherentnatureofbeingadisturbance,willaffecteachplantspeciesdifferentlydependinguponthespecies'abilitytosurviveandregenerate.
Shrublandplantspeciescanbegroupedintothreecategories,orfunctionalgroups,dependingupontheirpost-disturbanceregenerationresponse:obligateseed-ers,obligatesprouters,andfacultativeseeders(Keeley1991;Wells1969).
Therstfunctionalgroup,obligateseeders,primarilyreproducefromalong-livedre-stimulatedsoil-stored(e.
g.
,Ceanothuscuneatus,Arc-tostaphylosmanzanita)orcanopy-stored(e.
g.
Bank-sia)seedbanksandlackanysproutingability.
Duetotheirdependencyonre-relatedseedgerminationcues,veryfewobligateseederseedlingsareexpectedtoemergeaftermechanicaltreatments.
Incontrast,obligatesproutingspecies(thesecondfunctionalgroup)regenerateprimarilybysproutingvigorouslyfromadventitiousbuds(e.
g.
,Arctostaphylosglandul-osa,Quercusberberidifolia).
Thesespeciesareexpectedtoregeneratewellafterbothreandmasti-cationtreatments,butfewerresproutingspeciesmaysproutfollowinghighintensityres,sincehighlevelsofsoilheatingmaydamagemeristematictissues.
Obligatesprouterscanalsoreproducefromseed,butmostspeciesonlyproduceafewseedsthatareintolerantofheatexposure(e.
g.
,Quercusspp.
),andonlyasmallnumberofspecies(e.
g.
,A.
glandulosa)produceseedsthatrequirerecues,suchassmoke,heat,andcharatetostimulategermination(Keeley1987,1991).
Thethirdfunctionalgroup,facultativeseeders,reproducesbothsexuallybyseedandvege-tativelybysprouting(e.
g.
,Adenostomafasciculatum,chamise).
Theseedbankoffacultativeseedersisoftenpolymorphic,containingbothre-stimulatedandre-sensitiveseedstoensuregerminationafterandbetweenreevents(Keeley1991;ZammitandZedler1988),therefore,weexpectthisgrouptohaveastrongrecoveryafterdisturbance.
Seasonaltimingoffueltreatmentsisanimportantmanagementconsiderationduetoitsinuenceonthereproductivesuccessofeachfunctionalgroup.
Forexample,vegetativetissuesvaryintheirsensitivityandregenerationabilityindifferentseasons.
Inawetseasonre,lowreintensitiesmayfailtoreleaseseeddormancyformanyobligateseeders,butvegetativegrowthfromresproutersmaybemoreprolicsincelessbudtissueisdamaged.
Conversely,burningduringactivegrowthcanincreasemortalityduetoincreasedtissuesensitivity(FlorenceandFlorence1988).
Seasonaltimingisalsoanimportantdeterminantofseedsurvival.
Manysoft-coatedseeds(facultativeseeders,obligatesprouters)absorbwaterafterrainfallandbecomemorevulnerabletoheat(LeFerandParker2005).
Warmer,drierseasonprescribedres,incontrast,mayproducehighenoughtemperaturestocrackthehardseedcoatsofmostobligateseeders,butthesehighreintensitiesmayalsocausegreatermortalityforsproutingshrubspeciesandheatsensitiveseeds.
Theseasonoftreatmentwillalsostronglyaffectgerminantsurvival.
Aseedthatbreaksdormancyafterafalltreatmentwillhaveseveralmonthstoestablisharootsystembeforethesummerdrought,whereasaspringgermi-nantmaynothaveadequatetimetodeveloproots,andismoresusceptibletodrought-relatedmortality(FlorenceandFlorence1988).
Prescribedresandmasticationcanalsoinuencenon-nativeplantestablishmentandtheassociatedresourcecompeti-tion(PottsandStephens2009).
Shrubgrowthpatternscanbealteredduetodifferentphysiologicalactivityduringtheburntime(Drewaetal.
2002;RadosevichandConard1980;BondandvanWilgen1996).
Lowerresproutingshrubcoverafterspringtreatmentswouldbeexpectedbecausecarbohydrateandwaterreserveshavealreadybeenpartiallyexpended.
Firetreatmentsnotonlyaffectabovegroundvegetationbutalsocanchangesoilproperties.
Whenorganicmatterisconsumedbyre,plantnutrientsarereleasedandbecomehighlyavailableforplantgrowth(DeBanoetal.
1979).
Soilheatingbyrecanalsochangewaterrepellency(DeBano2000)andtherefore,prescribedburningmayhaveadifferentimpactonseedlingrecruitment.
Communitycompositionafterfueltreatmentsmayaffectfuturerepotential.
AdenostomaisconsideredmoreammablethanCeanothus(Biswell1974).
AshiftincommunitycompositionthatfavorsAdenostomamayultimatelycreateahigherriskreenvironment.
304PlantEcol(2010)210:303–315123Herbivorescanhaveastrongselectivepressureonspeciescompositionandabundanceinshrublands(ManierandHobbs2007),particularlyafterdistur-bancewhenregeneratingplantsareeasilyaccessibleandprovidehighlevelsofnutrition.
Obligateseederspeciesthatdependexclusivelyontheirseedbankforregenerationareparticularlyvulnerabletoseedpredators,whileobligatesproutingspeciescanbeheavilyinuencedbyvegetativebrowsers(O'NeilandParker2005).
Duetotheabundanceoflargeherbivoresinourresearchvicinity,includingblack-taileddeer(Odo-coileushemionus),rabbits(LepuscalifornicusandSylvilagus),androdents(Dipodomysagilis,Pero-myscusspp.
,Neotomaspp.
),thereisthepotentialforapronouncedherbivoreimpactontheregeneratingplantcommunity.
Herbivoresmayhavedifferenteffectsonplantrecoveryineachfueltreatment,sincethetypeandseasonoftreatmentcanresultinauniqueinteractionbetweenplantphenologyandbrowsingintensity.
Forexample,springtreatmentswouldtop-killmanysproutingspeciesatthetimeoflowcarbohydrateandwaterreserves,leavingtheplantwithfewresourcestoregenerateinthecoinci-dentperiodofheaviestbrowsingintensity(MorenoandOechel1991).
Herbivorepreferencesforparticularplantspeciesmayalsoinuencethecharacteroftherecoveringshrubcommunity.
Mills(1986)foundthatmamma-lianherbivoressignicantlydecreasedtheabundanceandsurvivorshipofCeanothusseedlingsafterre.
Intheir3-yearstudy,herbivoreselectivityforCeano-thusincreasedovertimeandgaveAdenostomaacompetitiveadvantage.
Adenostoma,however,mayalsosuccumbtodeerbrowsingwithsharplyreducedresproutingvigor(Davis1967).
Althoughmanyimportantstudieshaveexaminedplantrecoveryafterre,amajorityofresearchlackstreatmentreplication(BeyersandWakeman2000;Dunneetal.
1991),managementappropriatespatialscales(MorenoandOechel1991;Tyler1995),oreldsettings(Keeley1987;LeFerandParker2005).
Manyoftheeldstudieshavebeenperformedopportunis-ticallyafterwildresratherthanreplicatedprescribedres(MorenoandOechel1991;Tyler1995;ZammitandZedler1988).
Furthermore,masticationstudiesarerareinthepeer-reviewedliterature,andcomparative,replicatedstudiesofprescribedreandmasticationhaverarelybeenconductedinthechaparralecosystem.
Thegoalofourstudyistoexaminetheecologicalresponsesofthemostcommonfueltreatmentsinchaparralincluding(a)fallprescribedre,(b)winterprescribedre,(c)springprescribedre,(d)fallmastication(slashing),and(e)springmastication.
Inourresearch,weaddressthreequestions:(1)Doesfueltreatmenttypeand/orseasonaffectsproutingshrubcover,(2)Doesfueltreatmenttypeand/orseasonaffectshrubseedlingsurvival,and(3)Doesherbivoryaffectsproutingandseedingspeciesrecoveryaftertreat-ments.
Informationfromthisstudyshouldbeusefultomanagersandecologistsinterestedinshrublandeco-systemsinMediterraneanclimates.
MethodsStudysiteWeconductedourstudyinnorthernCalifornia'sCoastRangechaparral,approximately50kminlandfromthePacicOceanand175kmnorthofSanFrancisco,CA(39°N,123°W).
StudysitesarerepresentativeofCalifornianAdenostomafascicula-tum(chamise)chaparral,withchamisebeingthedominantspecies([65%oftheoverstorycover),andCeanothuscuneatus(buckbrush)andArctostaphylos(manzanita)speciesoccupyingmoreminoroverstorycomponents.
Pre-treatmentvegetationaveraged1–2minheightandnearly100%shrubcover.
Nearbyvegetationtypesincludemixedoakwoodlandonmesic,north-facingslopesandknobconepine(Pinusattenuata)standsonridgetopsandeast-facingslopes.
Chamise-dominatedchaparraloccupieslargeareasinthenortherncoastalrangesofCalifornia.
TheresearchareaexperiencesatypicalMediter-raneanclimatewithhot,drysummersandcool,wetwinters.
Maximummeantemperaturesvarywidelythroughouttheyear,averaging34°Cinsummerand7°Cinwinter.
The30-yearrainfallaverageis100cm,withnearlyallprecipitationoccurringbetweenOctoberandMay.
Annualrainfallduringour2001–2005studyaveraged99.
0cmandrangedfrom90.
7to124.
5cm.
Treatmentareasarelocated700–1,000mabovesealevelonsouthandwestaspectswithslopesof25–55%.
Soilsarederivedfromweatheredsandstoneandshaleandareclassiedasshallow,rocky,andmoderatelyacidic.
Fire-returnintervalsfortheseecosystemsbeforeresuppressionPlantEcol(2010)210:303–315305123wereapproximately30–70years(Stephensetal.
2007).
Fireandothermajordisturbanceswereabsentfromthestudyareaforatleast40yearspriortotreatment.
ExperimentaldesignEachofthevefuelreductiontreatmentswasreplicatedfourtimes;inaddition,fourcontrolplotswerealsoinstalledinthestudyarea(totalof24experimentalunits).
Eachexperimentalunitwasapproximately2ha,andunitsweredistributedacrossa\2kmdistance.
Treatmenttype(prescribedreormastication)andseason(fall/winter/spring)wererandomlyassignedtotheexperimentalunits,althoughmasticationwasrestrictedtositeswithslightlymoregradualslopesduetolimitedequipmentmaneuverabilityinsteepterrain.
Treatmentseasonsweredenedas:Fall(3–20November),Winter(8–18January),andSpring(31March–3Aprilforpre-scribedre,and23April–2Juneforspringmastica-tion).
Springmasticationtreatmentswereconductedslightlylaterthanspringretreatmentsbecauseroadswerenotdryenoughforthemasticatortoaccess.
Wedidnotexpectthisdiscrepancyinspringtreatmenttimingtosignicantlyaffectourndings,sincepreviousanalysisshowedthattreatmenttype(pre-scribedreormastication)overshadowedtheeffectoftreatmentseason(PottsandStephens2009).
Weperformedfuelreductiontreatmentsbetween2001and2003attheUniversityofCaliforniaHoplandResearchandExtensionCenterandadjacentUSBureauofLandManagementSouthCowMoun-tainRecreationArea.
Ourinitialgoalwastocompleteallfourreplicatesofeachtreatmentwithinthesameyear,butunfavorableweatherconditionsandequip-mentavailabilityforcedustosplitreplicatesover2years.
Prescribedreswereignitedwithdriptorchestoproduceupslopeheadres(Stephensetal.
2008).
Masticationwasperformedbyatrackbulldozerwithafrontmountedrotatingtootheddrumwhichshred-dedabovegroundbiomassintoapatchylayer(\5cmdeep)ofwoodydebristhatremainedonthesoilsurface.
Particlesizerangedfrom10to40cminlengthby5–10cminwidth.
Sub-surfacesoilandrootsystemswerenotdisturbedbythemasticator.
Treat-mentheterogeneitywasminimalinmasticationtreatments,sincetheequipmentoperatorperformedsystematicpassesthroughthevegetation.
Vegetationmortalitywas90–100%inbothreandmasticationtreatments.
Wedidnotimplementawintermastica-tiontreatmentduetounpredictableroadaccessandpotentialsoildamagecausedbytheheavymachineryinwetconditions.
MonitoringWeinstalled15permanentlinetransectsineachexperimentalunitpriortoreormasticationtomeasureshrubcoverusingthelineinterceptmethod.
Transectswere15minlengthandrandomlyplacedthroughouttheexperimentalunit;coverwasmeasuredbyplacingatapealongthetransectandcontinuouslyrecordingcoverbyspeciesorbareground.
Shrubheight(cm)wasmeasuredat5mintervalsalongeachlinetransect.
Datafromthe15linetransectswerepooledtorepresenttheexperimentalunit.
Attheendpointsof10randomlyselectedtransects,weestablishedpermanent2.
5m2vegetationsamplingquadratstodetermineseedlingabundanceandcom-position.
Aswiththetransectdata,wepooledseedlingdatafromthe10quadratstorepresenttheexperimentalunit.
Wemonitoredshrubandseedlingdatapriortotreatmentandagainduringthesecondandthirdsummersaftertreatment.
Controlplots(4replicates)weremonitoredbeforetreatmentswereinstalledtodetermineabaselinerangeofvariabilityamongtreatmentunits.
Asubsetofcontrolplotswasmoni-toredeachyearandshowednosignicantchangeinheight,cover,orcomposition.
Thisndingissimilarinotherstudiesthatalsoshowverylittlechangeinmaturechaparralstandsoverthestudyperiodintermsofcoverandheight(Hanes1971).
Immediatelyafterthefueltreatment,werandomlyselectedveofthe10seedlingquadratsforherbivoreexclosures.
Weconstructedexclosuresaroundthesequadrats,includinga20cmbufferzone,foratotalexclosedareaof12.
5m2foreachexperimentalunit.
Exclosureswerebuiltwith1.
5mtallwiremeshthatwascomposedof1.
27cmsquares.
Wesupportedthemeshwithmetalt-poststhatweredrivenintothesoil.
Wedidnotaddroofcoveringsbecausewedidnotexpectherbivorestoclimborjumpoverthemeshsides,andwewantedtominimizeshading,moistureaccumulationandotherindirectexclosureeffects.
Exclosureswereinitiallydesignedtoeliminatedeer,rabbit,androdentbrowsing,buterosioncreatedgaps306PlantEcol(2010)210:303–315123beneaththewiremeshthatwerelargeenoughforsmallanimalentry.
Althoughwebelievethattheexclosuresdeterredmostsmallmammals,wecanonlybecondentthattheexclosurecompletelyexcludedblack-taileddeer.
StatisticalanalysisOne-wayanalysisofvariance(ANOVA)wasusedtocheckfordifferencesbetweencontrolandmanipulatedplotsbeforetreatments(n=4).
FactorialANOVAwasusedtoassesstreatmenttypeandherbivoryeffectonshrubheight,cover,andseedlingabundanceaftertreatments.
IntheseANOVAtests,thetreatmentfactorhadvelevels,oneforeachtreatmenttype(FF=refall,FS=respring,FW=rewinter,MF=mas-ticationfall,MS=masticationspring);andtheex-closurefactorhadtwolevels(exclosedandunexclosed).
ThisfactorialANOVAhastheadvantagetobeabletoidentifypotentialstatisticalinteractionsbetweenfactorsaffectingshrubresponse.
Speciesrichnessandcompositionwereonlyassessedintheunexclosedareasbecauseexclosuresizewasnotlargeenoughtoaddressspeciesdiversity.
Wealsoper-formedrepeatedmeasuresANOVAtoaddresstreat-menteffect(betweengroupsfactor)onshrubcoverovertime(withinsubjectfactor)intheunexclosedareas.
Wepresentdatafromthesecondandthirdpost-treatmentyears,andexcludedatafromtherstpost-treatmentyearbecausespringtreatmentsdidnothavetimetoproducenewgrowthbeforethesummermonitoringperiod.
Weappliedpower,logarithm,andangular(arcsinesquareroot)transformationstotheseedlingdatatomeetANOVAassumptions.
Weusedanon-parametrictest(Kruskal–Wallis)whendatatrans-formationwasnotsufcienttomeetANOVAassump-tions.
TukeyHSDtestswereperformedtoidentifydifferencesbetweentreatmenttypeswhenANOVAresultsdemonstratedsignicanteffects(P\0.
05).
StatisticalanalysesweredonewiththeRstatisticalsoftwarepackage(http://www.
r-project.
org/).
ResultsShrubcoverandheightControlplotshadnosignicantdifferencesinshrubcover(F5,18=1.
55,P=0.
224)orheight(F5,18=0.
39,P=0.
846)comparedtoexperimentalplotsbeforetreatmentswereapplied.
Exclosedareasonlyhadsignicantlytallershrubsthanunexclosedareasonthethirdpost-treatmentyear(F1,30=1.
16,P=0.
290forthesecondyearandF1,30=5.
27,P=0.
028forthethirdyear;Fig.
1).
Treatmenttypehadnosignicanteffectonshrubheight(F4,30=1.
99,P=0.
122andF4,30=1.
45,P=0.
242forthesecondandthirdyear,respectively),andnointeractionbetweenfactorswasobserved.
Ingeneral,retreat-mentstendedtohavetallershrubsthanmasticationtreatmentsoutsidetheexclosuresinthethirdyear.
Regardingshrubcover,therewasnosignicantdifferencebetweenexclosedandunexclosedareasforbothpost-treatmentyears(F1,30=0.
28,P=0.
601forthesecondyearandF1,30=0.
82,P=0.
371forthethirdyear).
However,shrubcoverwassigni-cantlydifferentbytreatmenttype,withP\0.
0001inbothyears(F4,30=9.
90forthesecondyearandF4,30=11.
64forthethirdyear),andnosignicantinteractionbetweentreatmenttypeandexclosure.
Therefore,tobetteraddresscoverdifferencesbetweentreatments,wedecidedtoexcludedatafromtheexclosuresinthefollowinganalyses.
RepeatedmeasuresANOVAintheunexclosedareasconrmedahighlysignicanteffectoftreatmenttypeonshrubcover(F4,45=29.
76,P\0.
0001;Fig.
2).
Ingeneral,retreatmentshadhighercover010203040506070802yr-post3yr-postShrubheight(cm)unexclosedexclosedaaabFig.
1Averageshrubheight(±1SE)forexclosedandunexclosedareasandpost-treatmentyear(secondyearpost-treatmentandthirdyearpost-treatment).
Differentlettersdepictsignicantdifferencesinheightbetweentreatmentswithineachyearforasignicancelevela=0.
05PlantEcol(2010)210:303–315307123valuesthanmasticationtreatments,exceptforthesecondyearwheremechanicalspringandfallrewerenotsignicantlydifferentfromeachother.
Prescribedreplotshadashrubcoverrangingfrom59to72%bythesecondyearand67–76%bythethirdyear,comparedtomasticationplotcoverof29–42%and39–48%,inthesecondandthirdyears,respectively.
Treatmentseasondidnotaffectshrubrecovery.
Thechangeinshrubcoverovertime(withinsubjectfactor)washighlysignicant(F2,45=244.
98,P\0.
0001).
Shrubcoverreductionforbothpre-scribedreandmasticationwassignicant,withcovervaluesremainingsignicantlylowerthanpre-treat-mentvalues3yearsaftertreatmentswereapplied.
However,coverincreasebetweenthesecondandthethirdyearwasnotsignicantwithinanytreatmenttype,althoughMFtendedtohaveaslightlyhigherrecoveryrate(TukeyHSD,P=0.
064).
SpeciesrichnessandcompositionSpeciescompositionandrichnesswereassessedoutsideexclosures.
Priortotreatments,wefound14shrubspeciesalongthecovertransects,withanaverageof6±1speciesperexperimentalunit.
Adenostomafasciculatumwastheoverwhelminglydominantspecies(76%averagecover;Table1),followedbyCeanothuscuneatus(11%),Arctostaph-ylosspecies(6%),Heteromelesarbutifolia(1%),andQuercusspecies(\1%).
Aftertreatments,weidenti-ed16shrubspeciesalongthecovertransects.
Speciesrichnessdidnotdifferbetweentreatments(F4,15=1.
52,P=0.
247forthesecondyearandKruskal–Wallisv2=8.
03,P=0.
090,forthethirdyear)withanaverageof4±1speciesinthesecondyearand5±1speciesinthethirdyear.
Adenostomawastheprimaryspecies3yearsaftertreatments,withsimilarrelativecoverbetweentreatments(F4,15=0.
45,P=0.
774)rangingfrom91to95%,butdifferenttotalcoverbetweentreatments(F4,15=17.
86,P\0.
0001),withretreatmentsshowingsignicanthighervaluesofthisspeciescomparedtomastication.
Theremaining15speciesoccupied\1%covereachbythethirdpost-treatmentyear.
Ceanothuscuneatusaveragedonly0.
9%totalcoverandupto2%relativecoverafter3yearsandneverexceeded2.
5%totalcoveror6.
5%relativecoverinanyexperimentalunit.
ThefallretreatmenthadsignicantlyhigherCeanothuscuneatustotalandrelativecovercomparedtothespringandwinterre0102030405060708090100pre-treat2yr-post3yr-postShrubcover(%)FFFSFWMFMSaabbcccaabbbFig.
2Averageshrubcover(±1SE)foreachtreatment(FFrefall,FSrespring,FWrewinter,MFmasticationfall,MSmasticationspring)andmonitoringtime(pre-treatment,secondyearpost-treatmentandthirdyearpost-treatment).
Differentlettersdepictsignicantdifferencesincoverbetweentreat-mentswithineachyearforasignicancelevela=0.
05Table1Average±SEshrubcover(%)byspeciesforeachtreatmenttype(refall,respring,rewinter,masticationfall,andmasticationspring)before(pre-treatment)and3yearsaftertreatments(3yr-post)innorthernCaliforniachaparralSpeciesYearFireMasticationFallSpringWinterFallSpringAdenostomafasciculatumPre-treatment75.
1±4.
578.
1±3.
375.
1±6.
380.
7±2.
073.
9±4.
43yr-post63.
3±4.
072.
0±1.
365.
0±2.
336.
5±3.
343.
3±5.
6CeanothuscuneatusPre-treatment16.
2±2.
815.
5±3.
58.
4±4.
59.
0±2.
39.
9±3.
83yr-post0.
9±0.
30.
0±0.
00.
0±0.
00.
4±0.
20.
5±0.
2Othershrubspp.
Pre-treatment5.
6±2.
34.
4±2.
912.
8±4.
56.
6±2.
010.
1±1.
53yr-post3.
1±1.
54.
3±2.
35.
0±1.
11.
7±0.
93.
8±1.
5308PlantEcol(2010)210:303–315123treatmentsbythethirdpost-treatmentyear(F4,15=8.
25,P\0.
001intotalcoverandF4,15=8.
96,P\0.
001inrelativecover).
SeedlingestablishmentTotalshrubseedlingdensitywassignicantlydiffer-entbetweentreatmentsinboththesecondandthirdpost-treatmentyears(F4,30=29.
28,P\0.
0001forthesecondyearandF4,30=26.
15,P\0.
0001forthethirdyear),butdidnotdifferbetweenexclosedandunexclosedareas(F1,30=0.
03,P=0.
862forthesecondyear,andF1,30=0.
58,P=0.
449,forthethirdyear).
Inthesecondpost-treatmentyear,allretreatmentshadconsiderablyhigherseedlingdensitiesthanthemasticationtreatments,withanaverageof31±4seedlingsm-2(mean±standarderror)intheretreatmentscomparedto3±0seedlingsm-2inthemasticatedareas.
Bythethirdpost-treatmentyear,thespringretreatmenthadexperienceddispropor-tionatelyhighermortalityandexhibitedseedlingdensitiesthatweremuchmoresimilartothemasti-cationtreatmentsthantotheotherretreatments.
Attheendofthethirdpost-treatmentyear,thefallandwinterprescribedretreatmentshadsignicantlyhigherseedlingdensities,with19±3and18±4seedlingsm-2,respectively,comparedto3±1seedlingsm-2inthespringretreatmentand2±0seedlingsm-2inthemasticatedareas.
Adenostomawasthemostcommonshrubseed-linginallexperimentalunits(Fig.
3).
Adenostomaseedlingdensitysignicantlydifferedbetweentreatmentsinbothpost-treatmentyears(F4,15=9.
16,P\0.
001forthesecondyearandF4,15=15.
10,P\0.
0001forthethirdyear).
Inthesecondpost-treatmentyear,Adenostomaseed-lingdensitywasconsiderablyhigherintheretreatmentsthaninmasticationtreatments,althoughaverageseedlingdensityforthespringburnunitswaslargelyboostedbyasingletreatmentreplicatewithhighseedlingnumbers.
Bythethirdpost-treatmentyear,highseedlingmortalityinspringburnunitsreducedAdenostomaseedlingstosimilardensitiesasinmasticationtreatments.
FallandwinterretreatmentsstillhadhigherAdenostomaseedlingabundancecomparedtotheothertreatments.
Ceanothuscuneatuswasthesecondmostabundantplantspeciesinthepost-treatmentunits.
Thisspeciesalsoshowedsignicantlydifferentseedlingdensitiesbetweentreatmentsinboththesecondandthirdpost-treatmentyears(F4,15=7.
59,P=0.
001andF4,15=3.
09,P=0.
048,respectively;Fig.
3).
Inthesecondpost-treatmentyear,prescribedrehadhigherCeanothuscuneatusseedlingdensitythanmastication,butonlyresultsfromMFweresignicantlylower.
AlthoughMStendedtohavehigherseedlingdensitythanMF,therewasnosignicanteffectofseasoninmasticationtreatments.
Bythethirdtreatmentyear,alltreatmentshadsimilarseedlingdensity,exceptFFthathadsignicantlyhigherdensityofCeanothuscuneatusseedlingsthanFS.
DiscussionTreatmenttypeSignicantdifferencesinchaparralrecoverybetweentreatmentswereobserved,inbothtermsofresprout-ingandseedgermination.
Ingeneral,prescribedretreatmentsresultedinhighershrubcoverandseedlingdensitythanmasticationtreatments3yearsaftertreatments.
Highershrubcoverafterburningcom-paredtomechanicalremovalwasalsoobservedinotherstudiesindifferentshrublandecosystemsincludingshrublandsinFlorida(FitzgeraldandTanner1992),QuercuscocciferagarrigueinFrance(Godronetal.
1981),andMediterraneanheathland(Calvoetal.
1998,2002)andCistuslaurifoliusshrublandinSpain(Tarregaetal.
1997).
Conversely,shrubheightandspeciesrichnesswasnotsigni-cantlydifferentbetweentreatmentsinthisstudy.
Thelackofshrubheightdifferencebetweentreatmentsmaysuggestthatwoodyspeciesrecoveryduringtherstyearafteradisturbanceismainlyinvestedinreoccupyingthebaresurroundingspace,intheabsenceofcompetitionforlight(Marcosetal.
2004).
Calvoetal.
(1998)observedthatthemostpronouncedincreaseinheightoccurredoncetheshrubshaveoccupiedpracticallyalltheirinitialspace,achievingsimilarheightvalues9yearsaftertreatments.
Bytheendofourstudy(3yearsaftertreatments)shrubcoverwasapproximately70%inretreatmentsand40%inmasticatedareas,whichmayexplaintheabsenceofdifferentheightresponseamongtreatments.
Recoverytoinitialshrubcovervaluesoccurred4yearsafterburninginCaliforniaPlantEcol(2010)210:303–315309123chaparral(Dunneetal.
1991)ormechanicalandretreatmentsinMediterraneanheathland(Calvoetal.
1998)andgarrigue(Godronetal.
1981).
GratiniandAmadori(1991)reportedarapidresproutingofmaquisduringtherst4–5yearsafterawildreinsouthwesternItaly,withvegetationstructureandheightsimilartothoseofinitialcommunity8yearsafterre.
Shrubgerminationwasstimulatedintheretreatmentunits,resultinginhigherseedlingdensitiescomparedtomasticatedunits.
SimilarresultswerereportedbyBaezaandRoy(2008)inaMediterraneangorseshrublandinsoutheasternSpain.
Althoughsomeauthorsreportedthatincreasedsoilheatingbyinsolationaftershrubremovalwasenoughtostim-ulateseedgerminationinchaparral(ChristensenandMuller1975),weexpectedlowerseedlingdensitiesinthemasticationtreatmentsbecausethelackofrestimuliwoulddecreasethegerminationofspeciesthatrequireheat,char,orsmoketoreleasedormancy(Keeley1991).
Despitechaparralshrub-seedlingdensitybeinghigherinretreatments2yearsaftertreatments,weobservedhighermortalityratesinburnedplots,especiallyinspringres.
Ourstudy'sseedlingabundancepatternsaresimilartotrendsfoundinotherchaparralresearch(BeyersandWakeman2000;Guo2001;ZammitandZedler1988)whereseedlingnumbersareinitiallyveryhighaftertreatmentandthensuffersignicantmortalityinsubsequentyears.
Drasticmortalityratesintheearlyrecoveryyearscanbecausedbyacombinationoffactors,includingcompetition,herbivory,andtheinsufcientrootdevelopmenttohandlesummerdroughtconditions.
Seedbanklosscanlargelybeattributedtoerosion(Keeley1977)andcoincidenceofbaresoilandheavyrainmayincreaseerosion(Fernandezetal.
2008;GillandGroves1981).
Fernandezetal.
(2008)reportedthatfueltreatmentsaffectedtheinitialrunoff,inl-tration,anderosioninagorseshrublandcommunityinnorthwestSpain,especiallyafterprescribedburn-ingcomparedtomastication.
Initiallowerseedlingdensityinmasticationareasmayexplainthenegligiblemortalityobservedduetoincreasedsoilwaterresourcescausedbyfewerplants(MorenoandOechel1991).
However,Holmgrenetal.
(2000)reportedhigherseedlingsurvivalinburnedplotswhencomparedtomechanicallyclearedplotsinChileanshrubland.
Theseauthorsobservedthatearlyseedlingsurvivalwashigherundertheshadeoflargeshrubs.
Initialgerminationandseedlingsurvivalafterfueltreatmentsiscriticalinchaparralasseedlingrecruitmentisrareuntilthenextdisturbanceoccurs(Keeley1987).
Despitethehighseedlingmortalityobservedinretreatments,ourresultsstilldemonstratedhigherseedlingdensitiesinfallandwinterburningscomparedtospringburningandmastication3yearsafterthetreatments.
Adenostomafasciculatum01020304050FFFSFWMFMSbbbaabBBAAACeanothuscuneatus0123452yr-post3yr-post2yr-post3yr-postDensity(seedlingsm-2)Density(seedlingsm-2)FFFSFWMFMSBAABABABbbbaabFig.
3Averageseedlingdensity(±1SE)forAdenostomafasciculatumandCeanothuscuneatusbytreatmenttype(FFrefall,FSrespring,FWrewinter,MFmasticationfall,MSmasticationspring)andpost-treatmentyear.
Differentlettersdepictsignicantlydifferentseedlingdensitybetweentreat-mentsforthesecondyear(lowercases)andthirdyear(uppercase)forasignicancelevela=0.
05310PlantEcol(2010)210:303–315123Greatershrubrecoveryfollowingremaybeexplainedbythelargenutrientreleasethatresultsfrombiomasscombustionandashdeposition(Chris-tensen1973;ChristensenandMuller1975;DeBanoetal.
1979;RundelandParsons1980).
Masticatedsites,incontrast,haveaslownutrientreleaseastheshreddedbiomassdecaysandthismaycontributetotheslowerregrowth.
Eventhoughregenerationmechanismsofchaparralspeciesareconsideredadaptedtore,theresultsofourandotherstudiesinMediterraneanshrublandssuggestthatthesecom-munitieshaveahighresiliencetodisturbance,eithercausedbyreorbymechanicaltreatments(Calvoetal.
1998;DelBarrioetal.
1999;BondandMidgley2001;PottsandStephens2009).
Furthermore,non-redisturbancesaredemonstratedtoplayanimpor-tantroleintheevolutionofsomechaparralcommu-nities(Ackerly2004).
Thepresenceofre-followingspeciesinmasticatedareas(PottsandStephens2009)maybeattributedtopolymorphicseedbanks(Keeley1991;Sweeney1956;ZammitandZedler1994)and/orsufcientlyhighsolarheatingthatovercomesphysicalseeddormancy(BaskinandBaskin1998;ChristensenandMuller1975).
Calvoetal.
(2002)observedthatdifferencesinshrubcoverafterdiffer-entfueltreatments(includingburningandcutting)duringtherstyearofsuccessioninaMediterraneanheathlandtendedtobeeliminatedafter12years,andmostofthespeciestendedtorecovertotheirinitialcovervalues.
Thenumberofshrubspeciesdidnotdifferbetweenreandmechanicaltreatmentsinourstudy.
AsimilarresultwasfoundbyFitzgeraldandTanner(1992)inchaparralandbyCalvoetal.
(2005)inMediterraneanshrublandecosystems.
Regardingspe-ciescomposition,Adenostomaremainedthedomi-nantspeciesafterdisturbance,showinghighertotalcoverafterrethanmechanicaltreatments,butsimilarrelativecoverbetweentreatments.
Thispatternwasexpectedconsideringpre-treatmentAd-enostomadominanceandthespecies'abilitytobothresproutaftertop-killandgerminatefromasoil-storedseedbank(OdionandDavis2000).
Theversatileadaptationoffacultativeseedersmaybeanadvantageforrecoveringinawiderangeofenviron-mentalconditionsoccurringafterdisturbances(SeligmanandHenkin2000).
Non-dominantspecies,suchasCeanothuscunea-tus,haddifferentresponsesthanthedominantspecies.
Ceanothusisofparticularlystronginteresttolandmanagersduetoitsnutritionalvaluetomanybrowsingwildlifespecies(Biswell1974),includingblack-taileddeer.
Bythethirdyearaftertreatments,Ceanothuscuneatustotalcoverwasstilllowbecausethisspeciesonlyregeneratesbygermination,show-ingslightlyhigherrelativecoverinfallburntreat-mentsthaninthespringandwinterburntreatments.
Nevertheless,theslowrecoveryofCeanothusmaynotbedetrimentaltothisspecies.
Onceseedlingsareestablishedandnitrogenavailabilitydecreasesfol-lowingtheinitialushimmediatelypost-re,nitro-gen-xingspecies,suchasCeanothus,shouldhaveanincreasingcompetitiveadvantageovernon-sym-bioticshrubs(EllisandKummerow1988),especiallyinnitrogen-limitedecosystemslikechaparral(RundelandParsons1980).
InAdenostomadominatedchap-arral,Davis(1967)reportedCeanothuscoverincreased10yearsafterreinsouthernCalifornia.
TreatmentseasonSeasonoftreatmentdidnotsignicantlyaffectshrubresproutinginourstudy.
Resultsontreatmentseasoneffectsarevariedintheliterature.
FitzgeraldandTanner(1992)alsofoundnoeffectsofseasononeitherreormechanicaltreatmentsinshrublandsinFlorida.
Theseauthorsarguedthatsoilmoistureaffectspost-treatmentrecoveryandexplainedtheabsenceofdifferenceintheirstudytorelativelylowmoisturelevelsinbothseasonswhenthetreatmentswereapplied(winterandsummer).
Dunneetal.
(1991)alsoreportednonegativeeffectoffallburnsonchaparralrecoveryandGodronetal.
(1981)didnotobserveanydifferenceinvegetationcoverbetweenfallandspringburntreatmentsinFrenchgarrigue.
RadosevichandConard(1980)reportednoeffectonpost-reAdenostomashootgrowthbetweenburningseasonsinnorthernCaliforniachaparral.
Theseauthorsarguedthatpost-regrowthisnotdependentonthewaterstatusoftheshrubbeforetheburningtime,butonsubsequentsuitableenviron-mentalconditions.
Conversely,MorenoandOechel(1991)reportedthatincreasingreintensityincreasedplantmortality,reducedthenumberofresproutsperplant,anddelayedthetimeofresproutinginchaparral.
SimilarresultswerefoundbyLloretandLopez-Soria(1993)intheMediterraneanshrubEricamultiorawhenPlantEcol(2010)210:303–315311123applyinghigherintensityretoindividualplants.
FlorenceandFlorence(1988)alsoreportedincreasedAdenostomamortalityathighreintensitywhenburnswereconductedduringthegrowingorower-ingseason.
However,Canadelletal.
(1991)didnotobservedifferencesinthebiomassofresproutsofindividualshrubsburnedatdifferentreintensitiesinaMediterraneanshrubland.
OurndingsconictwithsomepreviousresearchthatfoundlowershrubsproutingafterspringprescribedreinsouthernCaliforniachaparral(BeyersandWakeman2000).
Althoughseasonoftreatmentdidnothaveastronginuenceonshrubreprouting,seasonoftreatmentdidhaveasignicantinuenceonseedlingdensity.
Fallandwinterretreatmentshadhigherseedlingdensitiesthananyothertreatment.
WeexpectedhighseedlingdensitiesinfallretreatmentsbecausethistreatmentbestapproximatesCaliforniachaparralwildreregimesbeforeresuppression(Stephensetal.
2007).
However,weweresurprisedtoobservesubstantialseedlingrecruitmentafterthewinterretreatments,sincemoistsoilconditionsatthetimeofburninghavebeenshowntohinderseedlingsuccessforsomespecies(LeFerandParker2005).
Labora-torygerminationresearchbyLeferandParker(2005)hasdemonstratedthatAdenostomaseedsaresensitivetoheatundermoistsoilconditionsduetotheamountofwaterabsorbedintotheseedandthelackofahardseedcoat.
Adenostomaseedlingdensitiesdidnotvarybetweenthethreeburningseasons2yearsaftertreatments,andremainednearlyidenticalinthefallandwinterretreatmentsbythethirdyearaftersubsequentmortality.
Masticationtreatmentsalsoshowedsimilarseedlingdensitiesforthisspeciesbetweenfallandspring.
Weexpectedthespringtreatmentstohavethelowestseedlingdensities,sincegerminatingplantshaveaninsufcienttimeandresourcestodevelopdrought-survivingrootsystemspriortothehot,dryMediterraneansummer(MooneyandParsons1973;Bondetal.
1984;SaruwatariandDavis1989).
Ourseedlingmortalitydataconrmthishypothesis.
ThehighestseedlingmortalitywasobservedinspringburningforbothAdenostomaandCeanothusseed-lingsindistinctively(85and87%,respectively).
Fallburning,whererebehaviorandeffectsareexpectedtobemoresimilartowildreinchaparralcommunities,resultedinmorethan50%seedlingmortalityforAdenostomacomparedto\23%forCeanothus.
Winterburningresultedinlowerseed-lingmortalityinAdenostoma(33%)comparedtoCeanothus(72%).
Although,theabilityofaseedtodevelopadequaterootspriortosummerdroughtmaybethestrongestinuenceonseedlingmortality,functionalgroupcharacteristicsmayalsobeanimportantfactorindeterminingseedlingmortalityandsurvival.
Consis-tentwithourndings,otherstudieshavereportedhighermortalityinfacultativeseederspeciescom-paredtoobligateseederspeciesafterwildre(FrazerandDavis1988;ThomasandDavis1989)andfallburning(OdionandDavis2000)insouthernCalifor-niachaparral,suggestingthatthelackofresproutingabilityamongobligateseedersisoffsetbyanenhancedabilitytoestablishseedlingsafterre.
Anotherchaparralstudyreportedthatobligateseed-ersseedlingarebetteradaptedtowaterstress,demonstratinghighersurvivalratesinopengapsandwithoutshadingfromadjacentshrubcanopies(Prattetal.
2008).
Regardingmastication,althoughfallandspringtreatmentshadsimilarseedlingdensities,seasonseemedtoaffectseedlingmortalityinaverydifferentmanner,withfallmasticationevenshowingseedlingrecruitmentbythethirdyearforbothAdenostomaandCeanothus.
Ourresultssuggestthat,intheabsenceofre-enhancinggermination,seedlingestablishmentafterfallmasticationtreatmentsoccurredforatleast3years.
HerbivoryThisstudydemonstratedarelativeeffectofherbi-voresonvegetationregrowthatourtreatmentscale.
Shrubheightwashigherintheexclosedareas3yearsaftertreatment;however,coverwasnotsignicantlyaffected.
Althoughtreatmenttypehadnosignicanteffectonshrubheight,theherbivoryeffectwashigherinmasticationcomparedtoretreatments.
Thiscouldbeexplainedbythepresenceofplantskeletonsafterburning,notpresentaftermastication,whichmayinterferewithlargemammalmovementpreventingaccesstobasalsprouts(Davis1967).
Itisimportanttonotethatourrelativelysmalltwo-hectaretreatmentareasmayhaveleadtointensebrowsingpressureduetotheirplacementinanotherwisedensechaparralmatrix.
Largertreatmentareasmayexperiencelessbrowsingpressure.
312PlantEcol(2010)210:303–315123Ourresultssuggestthatlargeherbivoresdonotextensivelybrowseshrubseedlingsintheearlyyearsfollowingdisturbance,sinceseedlingabundancewassimilarinexclosedandunexclosedareas(atleastatthespatialscalesexaminedhere).
PreviousherbivorystudiesinCaliforniachaparralalsofoundnosignif-icantexclosureeffectsonvegetationrecovery(Tyler1995,1996).
However,theseresultsareincontrasttootherstudies(Mills1983,1986)wheredifferencesinseedlingdensitywerefoundbetweenexclosedandunexclosedareas.
Nevertheless,itshouldbepointedoutthat,inourstudy,theexclosureswereonlyeffectiveagainstlargemammals,i.
e.
,black-taileddeer,andprobablydidnotpreventherbivoryfromsmallermammals,suchasrabbitsormice.
ManagementimplicationsInordertoimproveblack-taileddeerhabitat,pre-scribedrewouldbemostbenecialinfall,sinceitproducessignicantlyhigherCeanothuscuneatusseedlingdensityafterthethirdpost-treatmentyear.
Ingeneral,springretreatmentcouldbedetrimentalduetothelowerseedlingsurvivalratesduetosummerdrought.
Regardingmechanicaltreatments,fallandspringmasticationresultedinsimilarseedlingdensities,althoughfalltreatmentsforbothreandmasticationreducedseedlingmortalityasseedlingshadmoretimetodevelopadequaterootssystemspriortothesummerdrought.
Fromawildlifemanagementstandpoint,retreatmentshavefaster,densershrubregrowth,whichmayencouragewildliferecoverywithinashorttimeafterfueltreatment.
However,regardingwildreprevention,masticationtreatmentsaremoreeffectiveintermsofshrubfuel-loadaccumulationbecausetheyhavedramaticallyslowershrubregrowth,andtherefore,reducepotentialreintensityincaseofawildre.
However,rehazardswouldprobablybehigherinmasticatedareascomparedtothosetreatedwithprescribedreintheshort-termbecausenewlymasticatedareashavenedeadfuelsontheground(Bradleyetal.
2006)andincreasenon-nativegrasscover(PottsandStephens2009).
Inthepast,limitedplantrecoverydatahashinderedfuelsmanagementdecisionsandcausedpotentiallydetrimentalmanagementactions.
Regionaldifferencesinplantresponse,relianceonpost-wildredata(duetoalackofprescribedredata),andthealmostcompleteabsenceofmasticationresearchhavecontributedtotheconfusion.
Despitethiscomplexdecisionenvironment,itisimportantformanagerstomakethemostresponsibledecisionsbasedontheirobjectives.
Thegoalofourresearchwastoprovidemanagersandscientistswithdatatoreducepreviousuncertainty.
Clearly,moreresearchwillbeneededtollintheremaininggapsinknowledgeandtestthendingsofthisstudyinthelong-termandinothergeographicregions.
AcknowledgmentsWethanktheUSJointFireScienceProgram(Project#00-2-02)andtheUSFSRiversideFireLab(DavidWeise)forfundingthisresearch,andourprojectcooperatorsfromtheBureauofLandManagementUkiahFieldOfce(BLM),UniversityofCaliforniaHoplandResearchandExtensionCenter(HREC),andCaliforniaDepartmentofForestryandFireProtectionMendocinoandLakeCountyUnits(CALFIRE).
WeextendspecialthankstoJ.
Dawson(BLM),R.
Keiffer(HREC),T.
Derum(CALFIRE),M.
Tolbert(CALFIRE),andW.
Baxter(CALFIRE).
Wealsoextendourgratitudetonumerouseldassistants,particularlyD.
Fry,whoalsoreviewedthismanuscript.
EvaMarino'sparticipationinthisresearchwaspossiblethroughaPh.
D.
scholarshipfundedbyINIA(InstitutoNacionaldeInvestigacionyTecnologaAgrariayAlimentaria),Spain.
OpenAccessThisarticleisdistributedunderthetermsoftheCreativeCommonsAttributionNoncommercialLicensewhichpermitsanynoncommercialuse,distribution,andreproductioninanymedium,providedtheoriginalauthor(s)andsourcearecredited.
ReferencesAckerlyD(2004)Functionalstrategiesofchaparralshrubsinrelationtoseasonalwaterdecitanddisturbance.
EcolMonogr74:25–44BaezaMJ,RoyJ(2008)Germinationofanobligateseeder(Ulexparviorus)andconsequencesforwildreman-agement.
ForEcolManag256:685–693BaskinCC,BaskinJM(1998)Seeds:ecology,biogeography,andevolutionofdormancyandgermination.
AcademicPress,SanDiegoBeyersJL,WakemanCD(2000)SeasonofburneffectsinSouthernCaliforniachaparral.
In:KeeleyJE,Baer-KeeleyM,FotheringhamCJ(eds)2ndInterfacebetweenEcologyandLandDevelopmentinCalifornia.
U.
S.
GeologicalSurveyOpenFileReport00-62,pp45–55BiswellHH(1974)Effectsofreonchaparral.
In:KozlowskiTT,AhlgrenCE(eds)Fireandecosystems.
AcademicPress,NewYorkBondWJ,MidgleyJJ(2001)Ecologyofsproutinginwoodyplants:thepersistenceniche.
TrendsEcolEvol16:45–51PlantEcol(2010)210:303–315313123BondWJ,vanWilgenBW(1996)FireandPlants.
ChapmanandHall,LondonBondWJ,VlokJ,ViviersM(1984)VariationsinseedlingrecruitmentofCapeProteaceaeafterre.
JEcol72:209–221BradleyT,GibsonJ,BunnW(2006)Fireseverityandintensityduringspringburninginnaturalandmasticatedmixedshrubwoodland.
USDAForestServiceProc.
RMRS-P-41CalvoL,TarregaR,LuisE(1998)Space-timedistributionpatternsofEricaaustralisL.
subsp.
aragonensis(Willk)afterexperimentalburning,cutting,andploughing.
PlantEcol137:1–12CalvoL,TarregaR,DeLuisE(2002)ThedynamicsofMediterraneanshrubsspeciesover12yearsfollowingperturbations.
PlantEcol160:25–42CalvoL,TarregaR,LuisE,ValbuenaL,MarcosE(2005)Recoveryafterexperimentalcuttingandburninginthreeshrubcommunitieswithdifferentdominantspecies.
PlantEcol180:175–185CanadellJ,LloretF,Lopez-SoriaL(1991)ResproutingvigouroftwoMediterraneanshrubspeciesafterexperimentalretreatments.
Vegetatio95:119–126ChristensenNL(1973)FireandthenitrogencycleinCali-forniachaparral.
Science188:66–68ChristensenNL,MullerCH(1975)EffectsofreonfactorscontrollingplantgrowthinAdenostomachaparral.
EcolMonogr45:29–55DavisJ(1967)Someeffectsofdeerbrowsingonchamisesproutsafterre.
AmMidlNat77:234–238DeBanoLF(2000)Theroleofreandsoilheatingonwaterrepellencyinwildlandenvironments:areview.
JHydrol231–232:195–206DeBanoLF,RiceRM,ConradCE(1979)Soilheatinginchaparralres:effectsonsoilproperties,plantnutrients,erosionandrunoff.
USDAForestService,PSW-145DelBarrioJ,Luis-CalabuigE,TarregaR(1999)VegetativeresponseofArctostaphylosuva-ursitoexperimentalcut-tingandburning.
PlantEcol145:191–195DrewaPB,PlattWJ,MoserEB(2002)Fireeffectsonres-proutingofshrubsinheadwatersofsoutheasternlongleafpinesavannas.
Ecology83:755–767DunneJ,DennisA,BartolomeJW(1991)ChaparralresponsetoaprescribedreintheMountHamiltonrange,SantaClaracounty,California.
Madrono38:21–29EllisBA,KummerowJ(1988)N2(C2H2–C2H4)xationintwospeciesofCeanothusseedlingsinsecondyearpostrechaparral.
PlantSoil109:207–213FernandezC,VegaJA,FonturbelT,JimenezE,PerezJR(2008)Immediateeffectsofprescribedburning,choppingandclearingonrunoff,inltrationanderosioninashrublandareainGalicia(NWSpain).
LandDegradDev19:502–515FitzgeraldSM,TannerGW(1992)AviancommunityresponsetoreandmechanicalshrubcontrolinsouthFlorida.
JRangeManag45:369–400FlorenceSR,FlorenceMA(1988)PrescribedburningeffectsinCentralCaliforniachaparral.
Rangelands10:138–140FrazerJM,DavisSD(1988)Differentialsurvivalofchaparralseedlingsduringtherstsummerdroughtafterwildre.
Oecologia76:215–221GillAM,GrovesRH(1981)Fireregimesinheathlandsandtheirplant-ecologicaleffects.
In:SpechtRL(ed)Eco-systemsoftheworld:heathlandandrelatedshrublands.
Elsevier,Amsterdam,pp61–82GillAM,StephensSL(2009)Scienticandsocialchallengesforthemanagementofre-pronewildland–urbaninter-faces.
EnvironResLett4:034014GodronM,GuillermJL,PoissonetJ,PoissonetP,ThiaultM,TrabaudL(1981)Dynamicsandmanagementofvegeta-tion:effectsofreandgrazingonQuercuscocciferagarrigue.
In:DiCastriF,GoodhallDW,SpechtRL(eds)Mediterranean-typeshrublands.
Elsevier,Amsterdam,pp317–347GratiniL,AmadoriM(1991)Post-reresproutingofshrubbyspeciesinMediterraneanmaquis.
Vegetatio96:137–143GuoQ(2001)Earlypost-resuccessioninCaliforniachapar-ral:changesindiversity,density,coverandbiomass.
EcolRes16:471–485HanesTL(1971)SuccessionafterreinthechaparralofsouthernCalifornia.
EcolMonogr41(1):27–42HolmgrenM,SeguraAM,FuentesER(2000)LimitingmechanismsintheregenerationofChileanmatorral.
PlantEcol147:49–57KeeleyJE(1977)Seedproduction,seedpopulationsinsoil,andseedlingproductionafterrefortwocongenericpairsofsproutingandnonsproutingchaparralshrubs.
Ecology58:820–829KeeleyJE(1987)RoleofreinseedgerminationofwoodytaxainCaliforniachaparral.
Ecology68:434–443KeeleyJE(1991)SeedgerminationandlifehistorysyndromesintheCaliforniachaparral.
BotRev57:81–116LeFerD,ParkerVT(2005)Theeffectofseasonalityonburnonseedgerminationinchaparral:theroleofsoilmoisture.
Madrono52:166–174LloretF,Lopez-SoriaL(1993)ResproutingofEricamultioraafterexperimentalretreatments.
JVegSci4:367–374ManierDJ,HobbsNT(2007)Largeherbivoresinsagebrushsteppeecosystems:livestockandwildungulatesinuencestructureandfunction.
Oecologia152:739–750MarcosE,TarregaR,Luis-CalabuigE(2004)InteractionsbetweenMediterraneanshrubspecieseightyearsafterexperimentalre.
PlantEcol170:235–241MillsJN(1983)Herbivoryandseedlingestablishmentinpost-resouthernCaliforniachaparral.
Oecologia60:267–270MillsJN(1986)HerbivoresandearlypostresuccessioninSouthernCaliforniachaparral.
Ecology67:1637–1649MooneyHA,ParsonsDJ(1973)StructureandfunctionoftheCaliforniachaparral:anexamplefromSanDimas.
In:DiCastriF,MooneyHA(eds)Mediterraneantypeecosys-tems.
Springer,NewYork,pp83–112MorenoJM,OechelWC(1991)FireintensityandherbivoryeffectsonpostreresproutingofAdenostomafascicula-tuminsouthernCaliforniachaparral.
Oecologia85:429–433O'NeilSE,ParkerVT(2005)FactorscontributingtothesoilseedbanksizeoftwoobligateseedingCeanothusspeciesinNorthernCalifornia.
Madrono52:182–190OdionDC,DavisFW(2000)Fire,soilheating,andthefor-mationofvegetationpatternsinchaparral.
EcolMonogr70:149–169314PlantEcol(2010)210:303–315123PottsJB,StephensSL(2009)Invasiveandnativeplantresponsestoshrublandfuelreduction:comparingpre-scribedre,mastication,andtreatmentseason.
BiolConserv142:1654–1657PrattRB,JacobsenAL,MohlaR,EwersFW,DavisSD(2008)Linkagebetweenwaterstresstoleranceandlifehistorytypeinseedlingsofninechaparralspecies.
JEcol96:1252–1265RadosevichSR,ConardSG(1980)Physiologicalcontrolofchamiseshootgrowthafterre.
AmJBot67:1442–1447RundelPW,ParsonsDJ(1980)Nutrientchangesintwochaparralshrubsalongare-inducedagegradient.
AmJBot67:51–58SaruwatariMW,DavisSD(1989)Tissuewaterrelationsofthreechaparralshrubspeciesafterwildre.
Oecologia80:303–308SeligmanNG,HenkinZ(2000)RegenerationofadominantMediterraneandwarf-shrubafterre.
JVegSci11:893–902StephensSL,MartinRE,ClintonNE(2007)PrehistoricreareaandemissionsfromCalifornia'sforests,woodlands,shrublandsandgrasslands.
ForEcolManag251:205–216StephensSL,WeiseDR,FryDL,KeifferRJ,DawsonJ,KooE,PottsJ,PagniP(2008)MeasuringtherateofspreadofchaparralprescribedresinnorthernCalifornia.
FireEcol4:74–86StephensSL,MoghaddasJJ,EdiminsterC,FiedlerCE,HasseS,HarringtonM,KeeleyJE,McIverJD,MetlenK,SkinnerCN,YoungbloodA(2009)Firetreatmenteffectsonvegetationstructure,fuels,andpotentialreseverityinwesternU.
S.
forests.
EcolAppl19:305–320SweeneyJR(1956)Responsesofvegetationtore:astudyoftheherbaceousvegetationfollowingchaparralres.
UnivCalifPublBot28:143–249TarregaR,Luis-CalabuigE,AlonsoI(1997)Space-timehet-erogeneityintherecoveryafterexperimentalburningandcuttinginaCistuslaurifoliusshrubland.
PlantEcol129:179–187ThomasCM,DavisSD(1989)Recoverypatternsofthreechaparralshrubspeciesafterwildre.
Oecologia80:309–320TylerCM(1995)Factorscontributingtopostreseedlingestablishmentinchaparral:directandindirecteffectsofre.
JEcol83:1009–1020TylerCM(1996)Relativeimportanceoffactorscontributingtopostreseedlingestablishmentinmaritimechaparral.
Ecology77:2182–2195WellsPV(1969)TherelationbetweenmodeofreproductionandextentofspeciationinwoodygeneraoftheCaliforniachaparral.
Evolution23:264–267ZammitCA,ZedlerPH(1988)Theinuenceofdominantshrubs,reandtimesincereonsoilseedbanksinmixedchaparral.
Vegetatio75:175–187ZammitCA,ZedlerPH(1994)Organizationofthesoilseedbankinmixedchaparral.
Vegetatio111:1–16PlantEcol(2010)210:303–315315123
- Pinusrespring相关文档
- Cookerespring
- Evaporationrespring
- agenciesrespring
- pitocinrespring
- 练笔respring
- weatherrespring
月神科技:香港CN2/洛杉矶CN2/华中电信高防vps,月付20元起
月神科技怎么样?月神科技是由江西月神科技有限公司运营的一家自营云产品的IDC服务商,提供香港安畅、香港沙田、美国CERA、华中电信等机房资源,月神科技有自己的用户群和拥有创宇认证,并且也有电商企业将业务架设在月神科技的平台上。目前,香港CN2云服务器、洛杉矶CN2云主机、华中电信高防vps,月付20元起。点击进入:月神科技官方网站地址月神科技vps优惠信息:香港安畅CN2-GIA低至20元核心:2...

Megalayer新加坡服务器国际带宽线路测评
前几天有关注到Megalayer云服务器提供商有打算在月底的时候新增新加坡机房,这个是继美国、中国香港、菲律宾之外的第四个机房。也有工单询问到官方,新加坡机房有包括CN2国内优化线路和国际带宽,CN2优化线路应该是和菲律宾差不多的。如果我们追求速度和稳定性的中文业务,建议还是选择CN2优化带宽的香港服务器。这里有要到Megalayer新加坡服务器国际带宽的测试服务器,E3-1230配置20M国际带...
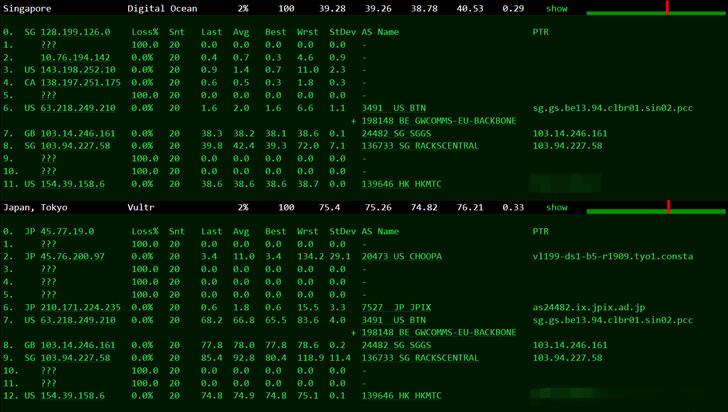
Virtono:圣何塞VPS七五折月付2.2欧元起,免费双倍内存
Virtono是一家成立于2014年的国外VPS主机商,提供VPS和服务器租用等产品,商家支持PayPal、信用卡、支付宝等国内外付款方式,可选数据中心共7个:罗马尼亚2个,美国3个(圣何塞、达拉斯、迈阿密),英国和德国各1个。目前,商家针对美国圣何塞机房VPS提供75折优惠码,同时,下单后在LET回复订单号还能获得双倍内存的升级。下面以圣何塞为例,分享几款VPS主机配置信息。Cloud VPSC...
respring为你推荐
-
小企业如何做品牌小公司的品牌建设怎么样才能做好sns平台SNS分类及代表性网站有哪些phpadmin下载免费MP3下载企业建网站我想建立一个企业网站,需要多少钱??sqlserver2000挂起安装sqlserver2000时总提示有挂起操作!设计eset面板flashyixingjia通配符的使用方法大飞资讯单仁资讯的黄功夫是何许人?电子商务世界世界前十大电子商务企业名字