suit55xoxo.com
55xoxo.com 时间:2021-04-07 阅读:()
RESEARCHOpenAccessRedescription,molecularcharacterisationandtaxonomicre-evaluationofauniqueAfricanmonitorlizardhaemogregarineKaryolysusparadoxa(Dias,1954)n.
comb.
(Karyolysidae)CourtneyA.
Cook1*,EdwardC.
Netherlands1,2andNicoJ.
Smit1AbstractBackground:WithintheAfricanmonitorlizardfamilyVaranidae,twohaemogregarinegenerahavebeenreported.
ThesecomprisefivespeciesofHepatozoonMiller,1908andaspeciesofHaemogregarinaDanilewsky,1885.
EventhoughotherhaemogregarinegenerasuchasHemoliviaPetit,Landau,Baccam&Lainson,1990andKaryolysusLabbé,1894havebeenreportedparasitisingotherlizardfamilies,thesehavenotbeenfoundinfectingtheVaranidae.
ThegenusKaryolysushastodatebeenformallydescribedandnamedonlyfromlizardsofthefamilyLacertidaeandtotheauthors'knowledge,thisincludesonlyninespecies.
Molecularcharacterisationusingfragmentsofthe18Sgenehasonlyrecentlybeencompletedforbuttwoofthesespecies.
Todate,threeHepatozoonspeciesareknownfromsouthernAfricanvaranids,oneoftheseHepatozoonparadoxa(Dias,1954)sharesmorphologicalcharacteristicsaliketospeciesofthefamilyKaryolysidae.
Thus,thisstudyaimedtomorphologicallyredescribeandcharacteriseH.
paradoxamolecularly,soastodetermineitstaxonomicplacement.
Methods:SpecimensofVaranusalbigularisalbigularisDaudin,1802(Rockmonitor)andVaranusniloticus(LinnaeusinHasselquist,1762)(Nilemonitor)werecollectedfromtheNdumoGameReserve,SouthAfrica.
Uponcaptureanimalswereexaminedforhaematophagousarthropods.
Bloodwascollected,thinbloodsmearsprepared,stainedwithGiemsa,screenedandmicrographsofparasitescaptured.
HaemogregarinemorphometricdatawerecomparedwiththedatafornamedhaemogregarinesofAfricanvaranids.
PrimersetHepF300andHepR900wasemployedtotargetafragmentofthe18SrRNAgeneandresultingsequencescomparedwithotherknownhaemogregarinesequencesselectedfromtheGenBankdatabase.
Results:Hepatozoonparadoxawasidentifiedinfectingtwooutofeight(25%)V.
a.
albigularisandasingle(100%)V.
niloticusexamined.
PhylogeneticanalysesrevealedthatH.
paradoxaclusteredwiththe'Karyolysus'clade,andnotwiththoseofreptilianHepatozoonspp.
Conclusions:InadditiontothisbeingthefirstmorphologicalandmolecularcharacterisationofahaemogregarinewithintheAfricanVaranidae,itisthefirstreportofaspeciesofKaryolysusinfectingthemonitorlizardfamily.
Furthermore,thisconstitutesnowonlythethirddescribedandnamedKaryolysusspeciesforwhichthereisanucleotidesequenceavailable.
Keywords:Haemogregarinetaxonomy,Hepatozoon,Monitorlizard,Haematozoa,Hemolivia*Correspondence:apicomplexan@yahoo.
co.
zaEqualcontributors1UnitforEnvironmentalSciencesandManagement,North-WestUniversity,Potchefstroom,SouthAfricaFulllistofauthorinformationisavailableattheendofthearticle2016TheAuthor(s).
OpenAccessThisarticleisdistributedunderthetermsoftheCreativeCommonsAttribution4.
0InternationalLicense(http://creativecommons.
org/licenses/by/4.
0/),whichpermitsunrestricteduse,distribution,andreproductioninanymedium,providedyougiveappropriatecredittotheoriginalauthor(s)andthesource,providealinktotheCreativeCommonslicense,andindicateifchangesweremade.
TheCreativeCommonsPublicDomainDedicationwaiver(http://creativecommons.
org/publicdomain/zero/1.
0/)appliestothedatamadeavailableinthisarticle,unlessotherwisestated.
Cooketal.
Parasites&Vectors(2016)9:347DOI10.
1186/s13071-016-1600-8BackgroundWithintheapicomplexanorderAdeleiorina,representa-tivesoftwohaemogregarinegenera,HepatozoonMiller,1908andKaryolysusLabbé,1894,arecommonlyre-portedinfectingsaurians.
ThegenusHemoliviaPetit,Landau,Baccam&Lainson,1990onthecontrary,eventhoughreportedparasitisingsaurianhosts,hasbutasingledescribedspeciesHemoliviamariaeSmallridge&Paperna,1997[1,2].
RepresentativesofHepatozoonarethemostcommonandarecosmopolitanparasitesfoundparasitisingawiderangeofvertebratehostsfromamphibiansandreptilestobirdsandmammals[3,4].
Karyolysus,conversely,isknownmainlyasasaurianhaemogregarinegenusthatprimarilyparasitiseslizardsofthefamilyLacertidae,buthasalsobeenreportedfromlizardsoftheScincidae[1,5–7].
Besidesthisdiscrepancyinvertebratehostpreferenceofthespeciesintheabovehaemogregarinegenera,speciesinthesegeneraalsodemonstratedifferentdevelopmentalpatterns.
Eventhoughspeciesofallthreeofthehaemogregarinegeneramaybetransmittedtothesaurianhostthroughthein-gestionoftheinfectedinvertebratevector,Hepatozoonspp.
maybetransmittedthroughawiderangeofarthro-podvectors(mosquitoestoticks),whilsttransmissionofHemoliviaspp.
andKaryolysusspp.
hasbeenrecordedonlythroughatickandmitevector,respectively[1].
Whilstmorethan30Hepatozoonspp.
havebeenre-cordedfromsauriansthroughoutAfrica[8],Karyolysusspp.
havemainlybeenreportedfromlacertidlizardsofEuropeandAsia[7].
UntilSmith's[3]revisionoftheHepatozoidae,thegenusKaryolysuscomprised11species,Smith[3]reassigningtwoofthesetoHepatozoon,nowHepatozoonberestnewi(Finkelstein,1907)andHepatozoonbicapsulata(Franca,1910).
Totheauthors'knowledge,todate,ninespeciesofKaryolysusareconsid-eredvalid:KaryolysuslacertaeDanilewsky,1886;Karyoly-suslacazeiLabbé,1894;KaryolysusbiretortusNicolle,1904;KaryolysuszuluetaiReichenow,1920;KaryolysussubtilisRicci,1954;KaryolysusoctocromosomiAlvarez-Calvo,1975;KaryolysuslatusSvahn,1975;KaryolysusminorSvahn,1975;andtheonlyspeciesreportedfromsub-SaharanAfricaKaryolysuspoleensisMutinga&Dipeolu,1989[3,6,7].
Descriptionsofhaemogregarinespecieswereuntilrecentlybasedonmorphologicalchar-acteristicsandlife-cycledata[7].
ThisisparticularlytrueofthehaemogregarinesdescribedfromsaurianswithinsouthernAfrica.
Haemogregarines,specificallyspeciesofHepatozoon,havebeencommonlyrecordedbloodpara-sitesofsouthernAfricansauriansincludingthosefromlizardgeneraofthefamiliesCordylidaeandVaranidae[8,9].
WithintheAfricanVaranidae,sixspeciesofHepa-tozoonhavebeendescribed,threeofthesefromsouthernAfrica(Table1),thelatterthreecomprisingHepatozoonvarani(Laveran,1905)fromVaranusniloticus(Linnaeus,1762)inSouthAfrica[3,6],Hepatozooncamarai(Dias,1954)andHepatozoonparadoxa(Dias,1954)fromVaranusalbigularisalbigularisDaudin,1802inMozambique[3,6].
TheaimofthepresentstudywasthustoprovideamorphologicalredescriptionofH.
paradoxaandmo-leculardataaidinginthecorrecttaxonomicplacementofthisparasite.
MethodsStudyarea,Varanusspp.
collectionandbloodpreparationSpecimensofVaranusalbigularisalbigularisandVaranusniloticuswerecollectedindaylightduringthesummermonthsofNovember2013,FebruaryandNovember2014,andFebruary2015intheNdumoGameReserve(NGR)(26°52′00.
0″S,32°15′00.
0″E),north-easternKwaZulu-Natal(KZN),SouthAfrica,borderingsouthernMozambique[10].
Lizardswererestrainedbyhandwhilstbloodandanyhaematopha-gousathropodswerecollectedinsitu.
Asmallvolumeofblood(approximatelyonedrop)wascollectedfromtheventralcaudalveinusinganappropriatelygauged(de-pendingonthesizeofthelizard)sterileneedleand1mlsyringe.
Asmallportionofthecollectedbloodwasusedtoprepare2–3duplicatethinbloodsmearsandthere-mainderdroppedintoanequalvolumeof70%ethanolforfuturemolecularanalysis.
Thinbloodsmearsonceair-driedinadustproofcontainerwerefixedinabsolutemethanolandstainedthereafterusingamodifiedsolu-tionofGiemsastain(FLUKA,Sigma-Aldrich,Steinheim,Germany)accordingtothemethodsof[11,12].
ScreeningofVaranusspp.
bloodsmearsSmearswerescreenedundera100*oilimmersionob-jectiveonaNikonEclipseE800compoundmicroscope(Nikon,Amsterdam,TheNetherlands)andimageswerecapturedwithanattachedNikondigitalcameraandac-companyingsoftware.
HaemogregarineswereidentifiedtospecieslevelbycomparingmorphometricdatatothatofpreviousstudiesonAfricanVaranusspp.
haemogre-garines[9,13–15](seeTable1).
Parasitaemiawascalcu-latedper100erythrocytes,withc.
104erythrocytesexaminedperbloodsmear[16–18].
DNAextraction,PCRamplificationand18SrDNAsequenceanalysisEthanol-preservedbloodsampleswereusedformolecu-larwork.
GenomicDNAofhaemogregarinespecieswasextractedfromthesamplesusingarapidDNAextrac-tionmethodasdetailedintheKAPAExpressExtractKit(KapaBiosystems,CapeTown,SouthAfrica).
Basedonpreviousstudies,amplifyingfragmentsofthe18SrRNAgeneofreptilehaemogregarinesofthegeneraKaryolysusCooketal.
Parasites&Vectors(2016)9:347Page2of13[7],Hemolivia[12]andHepatozoon[19],identificationoftheparasiteofthetwoVaranusspecies,twoV.
aalbigularisandoneV.
niloticus(n=3)fromthecurrentstudywascompletedusingtheprimersetHepF300(5′-GTTTCTGACCTATCAGCTTTCGACG-3′)andHepR900(5′-CAAATCTAAGAATTTCACCTCTGAC-3′).
ThePCRreactionswereruntargetingafragment(approximately600nt)ofthe18SrRNAgene[20].
ConditionsforPCRwereasfollows:initialdenatur-ationat95°Cfor3min,followedby35cycles,entailinga95°Cdenaturationfor30s,annealingat60°Cfor30swithanendextensionat72°Cfor1min,andfollowingthecyclesafinalextensionof72°Cfor10minasdetailedaccordingtopreviousmethods[12,18].
PCRreactionswereperformedwithvolumesof25μl,using12.
5μlThermoScientificDreamTaqPCRmastermix(2*)(2*DreamTaqbuffer,0.
4mMofeachdNTP,and4mMMgCl2),1.
25μlofeachprimer,andatleast25ngDNA.
ThefinalreactionvolumewasmadeupwithPCR-gradenucleasefreewater(ThermoScientific,Vilnius,Lithuania).
ReactionswereundertakeninaBio-RadC1000TouchThermalCyclerPCRmachine(Bio-Rad,HemelHempstead,UK).
Resultingampliconswerevisualizedunderultravioletlightona1%agarosegelstainedwithgelredusingaBio-RadGel-DocXR+imagingsystem(Bio-Rad,HemelHempstead,UK).
TwoPCRproductsfromeachsampleweresenttoacommercialsequencingcompany(InqabaBiotechnicalIn-dustries(Pty)Ltd,Pretoria,SouthAfrica)forpurificationandsequencinginbothdirections.
Resultantsequenceswereassembled,andchromatogram-basedcontigsweregeneratedandtrimmedusingGeneiousVer.
7.
1[21].
Se-quenceswereidentifiedusingtheBasicLocalAlignmentSearchTool(BLAST)[22],anddepositedintheNCBIGenBankdatabaseunderaccessionnumbersKX011039andKX011040.
ComparativesequencesforspeciesofHemolivia,Hepatozoon,Karyolysus,Haemogregarina,DactylosomaLabbé1894andBabesiosomaJakowska&Nigrelli,1956parasitisingreptiles,amphibians,mammalsandticksweredownloadedfromGenBankandalignedtothese-quencesgeneratedinthisstudy.
AdelinadimidiataSchneider,1875,AdelinagrylliButaeva,1996(GenBank:DQ096835–DQ096836)andKlossiahelicinaSchneider,1875(GenBank:HQ224955)werechosenastheout-grouptorootthephylogeny.
SequenceswerealignedusingtheMUSCLEalign-menttool[23]implementedinGeneious7.
1.
TheTable1SpeciesofhaemogregarinesofthegenusHepatozoondescribedfromAfricanvaranidsSpeciesType-hostType-localityOtherhosts(localities)Peripheralgamont/nucleusdimensionsReferenceHepatozoonborreli(Nicolle&Comte,1906)Smith,1996bVaranusgriseus(Daudin,1803)Tunisia7–8*2/1–3*1–2[3]b,[6,14]Hepatozooncamarai(Dias,1954)Smith,1996bVaranusalbigularisalbigularis(Daudin,1802)Mozambique2formsobserved:banana-shaped:11.
75*5.
00;longcurved:14.
25–18.
25*1.
25–4.
25/nucleusirregular[3]b,[9]Hepatozoonparadoxa(Dias,1954)Smith,1996bV.
a.
albigularis1MozambiqueV.
niloticus(Kenya)2;V.
a.
albigularisandV.
niloticus(SouthAfrica)36.
75–7.
50*4.
25–5.
50/nucleusnotvisible1;8.
1*5.
2/nucleusirregularornotvisible2;6.
99*4.
39/nucleusnotvisible3[9]1,[30]2,thisstudy3,[3]bHaemogregarinaroshdyiRamdan,Sauod,Mohammed&Fawzi,1996[probablyHepatozoonroshdyi(Ramdan,Sauod,Mohammed&Fawzi,1996)]V.
griseusEgypt13–20*1.
5–2.
5/6.
0–8.
5*1.
5–2.
5[40]aHepatozoontoddi(Wolbach,1914)Smith,1996bVaranusniloticus(Linnaeus,1762)TheGambia10.
3*2.
5/notgiven[3]b,[6,15]Hepatozoonvarani(Laveran,1905)Smith,1996bV.
niloticus1SouthAfricaV.
niloticusandV.
griseus(Senegal)2;V.
niloticus()(PortugueseGuinea)3;V.
niloticus(TheGambia)4;V.
niloticus(Senegal)5;VaranusarenariusDuméril&Bibron,1836()(FrenchWestAfrica)6;V.
griseus(FrenchSudan)7;V.
niloticus(Kenya)814*3/notgiven1;11–15*3/notgiven2;12.
0*13/3.
5–4.
5*3.
03;10.
3*2.
5/notgivena4;twoforms:12–14*2/notgiven,10–12*4–5/5–6*4–55;reportonly6;reportonly7;12.
7*4.
6/notgiven8[13]1,[41]2,[15]3,[42]3,[43]4,[44]5,[45]6,[46]7,[30]8,[3]baHepatozoon(syn.
Haemogregarina)toddi(Wolbach,1914)whenfirstdiscoveredwastentativelythoughttobeHepatozoon(syn.
Haemogregarina)varani(Laveran,1905)bSmith[3]duringasystematicrevisionofspeciesoftheHepatozoidaetransferredmanyoftheabovespeciesfromthegenusHaemogregarinaDanilewsky,1885tothegenusHepatozoonMiller,19081–8CorrespondingdescriptionorreportofthehaemogregarinespeciesanditsbibliographicreferenceCooketal.
Parasites&Vectors(2016)9:347Page3of13alignmentconsistedof47sequences,manuallytrimmedtoatotallengthof968nt.
Uncorrectedpair-wisedis-tances(p-distance),basepairdifferencesaswellaspar-simonyinformativesitesandthenumberthereofwereidentifiedordeterminedwiththeMEGA6bioinformat-icssoftwareprogram[24]forthealigned18SrDNAse-quencesbetweenallavailablespeciesappearinginthephylogeneticanalyses.
ToinferphylogeneticrelationshipsofthealigneddatasetbothBayesianinference(BI)andMaximumlikelihood(ML)methodswereused.
Acomprehensivemodeltestwaspreformedtodeterminethemostsuit-ablenucleotidesubstitutionmodel,accordingtotheAkaikeinformationcriterionusingjModelTest2.
1.
7[25,26].
ThebestmodelidentifiedwastheTransver-sionModelpluswithestimatesofinvariablesitesandadiscreteGammadistribution(TVM+I+Γ).
ThismodelwassubstitutedwiththeGeneralTimeReversiblemodel(GTR+I+Γ)forphylogeneticanalysis,asthiswasthemostap-propriatemodelavailablewiththebestAICcscore.
TheBIanalysiswasimplementedfromwithinGeneious7.
1usingMrBayes3.
2.
2[27].
Theanalysiswasruntwiceover10milliongenerationsfortheMarkovChainsMonteCarlo(MCMC)algorithm.
TheMarkovchainwassam-pledevery100cycles,andtheMCMCvariantcontained4chainswithatemperatureof0.
2.
Thelog-likelihoodvaluesofthesamplepointwereplottedagainstthegenerationtimeandthefirst25%ofthetreeswerediscardedas'burn-in'withno'burn-in'samplesbeingretained.
TheMLanalysiswasperformedusingRAxMLVer.
8.
1.
22[28]implementedintheraxmlGUIVer.
1.
3[29].
Thealpha-parameterselectedwastheGTR+I+Γmodel,withsupportassessedusing1,000rapidbootstrapinferences.
Resultingtreeswerecombinedina50%majorityconsensustree.
EthicsstatementThisstudyreceivedtherelevantethicalapproval(North-WestUniversityethicsapprovalno:NWU-00005-14-S3).
ResultsPrevalence,parasitaemiaandgeneralobservationsofH.
paradoxainperipheralbloodsmearsSixadultandtwojuvenileVaranusalbigularisalbigu-laris(Fig.
1a,b)andoneadultVaranusniloticuswerecaptured,sampledforbloodparasitesandexaminedforpossiblehaematophagousarthropods.
Thetwojuvenileswerefoundtobenegativeforbloodparasites,whilst4/6(67%)adultV.
a.
albigularisand1/1(100%)adultV.
niloticuswerefoundpositiveforhaemogregarineinfec-tions.
Twoadult(33%)(withaparasitaemiaofc.
5and20%,respectively)V.
a.
albigularisandtheV.
niloticus(withaparasitaemiaofc.
0.
2%)wereparasitisedbyahaemogregarinealiketoHepatozoonparadoxadescribedbyDias[9](Figs.
1c–fand2b–i).
Oneofthetwo(17%)V.
a.
albigularis(parasitaemiaof5%)andthesingleV.
niloticuswerefoundtohaveaco-infectionwithanotherhaemogregarineofadifferentHepatozoonspp.
(Cook,Netherlands&Smit,unpublishedobservations)Fig.
1Karyolysusparadoxa(Dias,1954)invaranidlizardVaranusalbigularisalbigularisDaudin,1802.
a–bVaranusalbigularisalbigularis.
bTicksofthespeciesAmblyommaexornatuminfestingtheareaabovetheeyes,theperipheryofthemouthanddeepintothenostrils(arrows).
c–fPeripheralbloodstagesofK.
paradoxacapturedfromtheneohapantotypeslide(NMBP410).
cPossibleraretrophozoitestage,notethattheyounghosterythrocytecytoplasmandnucleusarestillintactandthattheparasitenucleusisvisibleandgranular.
d–fMaturegamontswithinanerythrocyteinwhichshrinkageofthehostcellisapparentandthenucleusdestroyedresultinginaheavilyvacuolatedappearance.
dMaturegamontinwhichfoldingofthegamontmaybeseenwithinthethickcapsule(arrow).
Scale-bar:10μmCooketal.
Parasites&Vectors(2016)9:347Page4of13(parasitaemiaofc.
0.
4and3%,respectively).
ThelatterunidentifiedHepatozoonspecieswasalsofoundinfectingtheremainingtwoparasitisedV.
a.
albigularis(parasit-aemiaofc.
0.
1and0.
5%,respectively).
AllspecimensofbothspeciesofVaranuswereinfestedwithadultandjuvenilestagesoftheLeguantickAmblyommaexornatum(Koch,1844)(Fig.
1b),thetwojuvenileV.
a.
albigularisandV.
niloticusdemonstratinglowerdensitiesoftheseectoparasites.
Theseectoparasiteswerefoundallovertheanimal,withthehighestdensitiesinthenostrils,oftenblockingthem,alsoontheareasurroundingtheeyes,andalongtheedgesofthemouth.
Itwasnotuncommontofinddeadticksandtheirremainsdeepwithinthenostril,theirabilitytoexitblockedbylaterarrivals.
Squashesofnymphalandadultfemaleandmaleticksprovidednoparasiticstages.
Nootherhaematophagousarthropods,includ-ingmites,wereobserved.
StagesoftheH.
paradoxa-likehaemogregarineobservedinperipheralbloodsmearsfromthisstudywerecomparedmorphologicallywiththoseobservedinpreviousbloodparasitestudiesofAfricanvaranids[9,13–15](Table1).
Insizeandmorphology,theH.
paradoxa-likestagesobservedduringthecurrentstudyconformedtothosede-scribedbyDias[9]andBall[30](seeTable1,Figs.
1a–f,2b–iand3a–d,respectively).
Twostagesoftheparasitewereobserved:astageunreportedbyDias[9],butpossiblybyBall[30],ararepossibletrophozoitestage(Figs.
1cand3a),andwhatwasidentifiedasamatureintra-erythrocyticgamontstage(Figs.
1d–f,2b–iand3d,e).
Karyolysusparadoxa(Dias,1954)Cook,Netherlands&Smit,2016SynsHaemogregarinaparadoxaDias,1954;HepatozoonparadoxaSmith,1996.
Type-host:VaranusalbigularisalbigularisDaudin,1802,Squamata:Varanidae[9].
Otherhosts:Varanusniloticus(LinnaeusinHasselquist,1762),Squamata:Varanidae[30];presentstudy.
Vector:Unknown.
Type-locality:NdumoGameReserve(26°54′18.
5″S,32°19′24.
7″E),KwaZulu-Natal,SouthAfrica(presentstudy).
Otherlocalities:Maputo,Mozambique[9].
Ball[30]alsodescribedahaemogregarinemorphologicallyaliketoK.
paradoxainMarimonti,nearMeru,Kenya.
Fig.
2IllustrationofHaemogregarinaparadoxaDias,1954inVaranusalbigularisalbigularisDaudin,1802[9].
a-iRedrawnandadaptedfromDias(1954).
IllustrationsrepresentingtheoriginaldescriptionofKaryolysusparadoxa(syns.
HepatozoonparadoxaandHaemogregarinaparadoxa)exVaranusalbigularisalbigularisfromMozambique.
aHealthynon-parasitisederythrocyte.
b-iParasitisederythrocytes,notetheshrinkageofthehostcell,theheavyvacuolizationofthehostcellnucleus,andthethickcapsulesurroundingthegamontwhichresultsinthegamontnucleusbeinginvisible.
Scale-bar:10μmCooketal.
Parasites&Vectors(2016)9:347Page5of13Type-material:Neohapantotype,1*bloodsmearfromthetype-hostVaranusalbigularisalbigularisandnewdes-ignatedlocality(26°54′18.
5″S,32°19′24.
7″E),depositedintheprotozoancollectionoftheNationalMuseum,Bloem-fontein,SouthAfricaunderaccessionnumberNMBP410.
OthervouchermaterialdepositedthatincludesstagesofK.
paradoxa,1*bloodsmearfromVaranusniloticus,depositedintheprotozoancollectionoftheNationalMu-seum,Bloemfontein,SouthAfricaunderaccessionnum-berNMBP411.
RepresentativeDNAsequences:Twosequencesrepre-sentinga611and613ntfragmentofthe18SrRNAgeneofK.
paradoxaisolatedfromthetype-hostVaranusalbi-gularisalbigularis,depositedintheNCBIGenBankdatabaseundertheaccessionnumbersKX011039andKX011040,respectively.
Redescription(Figs.
1,2and3)Trophozoite.
Rare,ovoid,withvacuolatedcytoplasm,measuring6.
5–6.
9*4.
3–4.
7(6.
7*4.
5)μm(n=2);nucleuswithloosechromatin,stainingpink(Fig.
1c).
Bothtrophozoitesparasitisingyoungerythrocytes,nohostcelldistortionvisible.
Maturegamont.
Roundedinshape,gamontseeminglyfoldedwithinwithawell-developedcapsule(Fig.
1d–f),measuring6.
3–7.
9*3.
6–5.
2(7.
0*4.
4)μm(n=20).
Cytoplasmstainingwhitish-blue;nucleusnotvisible.
Notabledestructionofhostcellcytoplasmandkaryolysisofthehostcellnucleus,causinganobservableheavilyvacuolatedandfoamyappearance(Fig.
1d–f).
RemarksThehaemogregarinedescribedinthisstudyfromSouthAfricanVaranusalbigularisalbigularisandVaranusniloticus(Fig.
1d–f)wasfoundtobemorphologicallysimilartoHepatozoonparadoxadescribedbyDias[9]fromaspecimenofV.
a.
albigularisinneighbouringMozambique(Fig.
2b–i).
Itsharedanumberofuniquecharacteristicsincludingdestructionoftheinfectedhosterythrocyte,consistingofdehaemoglobinisationresultinginshrinkageofthehostcellanddestructionofthehostcellnucleus(characteristicofanumberofspeciesofKaryolysus[1])resultinginaheavilyvacuolatedappear-ance(Figs.
1d–fand2b–i).
Additionally,thehaemogre-garineinthisstudyagreeswellwiththesizeofH.
paradoxaintheoriginaldescriptionofDias[9](mean7.
0*4.
4vs7.
0*4.
9μm)(Table1).
ThesameuniquecharacteristicswerereportedofahaemogregarinefoundinfectingaV.
niloticusbyBall[30]fromKenya,measuringonaverage8.
1*5.
2μm(Fig.
3c,d).
However,inBall's[30]study,additional,pre-sumablyyounger,stageswereobserved(similartotheyoungtrophozoitestagefoundinourstudy)(Figs.
1cand3a).
Ball[30]alsonotedasinglepossiblydividingstageofthesetrophozoites(Fig.
3b).
Incellsparasitisedbyallthesepossibletrophozoitestages,thehosterythro-cyteshowednoshrinkageasofyet,butwasaccordingtoBall'sreportabnormalinshapeandstaining.
Atfirst,Ball[30]didassumethisparasitetorepresentyoungerstagesofanotherhaemogregarinethathasbeenreportedparasitisingAfricanvaranidsHepatozoonvarani(Lave-ran,1905)Smith,1996(Table1).
However,basedontheeffectsoftheparasiteresemblingH.
paradoxaasde-scribedbyDias[9],itsdestructionofthehostcellandthehostcell'snucleus,heconcludedthatthisparasitewasnotH.
varani.
Overall,fortheK.
paradoxade-scribedinthisstudyandtheparasitesdescribedintheothertwostudies[9,30],thenucleusandcytoplasmwasnotvisibleowingtowhatappearedtobeathicken-closingcapsuleasseenwiththegamontsofspeciesofHemolivia,see[12,31](Figs.
1d–f,2b–iand3c,d).
OnlyonrareoccasioninthepresentstudyandthatofBall's[30]wastheparasiteseentobefoldingFig.
3IllustrationofanunknownhaemogregarinebyBall(1967)[30]inVaranusniloticus(LinnaeusinHasselquist,1762).
a–dRedrawnandadaptedfromBall(1967)(hisFigs.
19–22).
IllustrationsrepresentinganunknownhaemogregarinefoundasaconcurrentinfectiontoHepatozoonvarani(Laveran,1905)(syn.
Haemogregarinavarani)inaVaranusniloticusfromKenya.
aPossibletrophozoitestageoryoungstage,shrinkageandvacuolationofthehostcellandhostcellnucleusnotyetapparent.
bPossibledividingstage.
c–dMaturegamontstages,notetheshrinkageandheavyvacuolationofthehostcellerythrocyte.
cMaturegamontfoldedwithinacapsulewithpossiblenucleus.
dFoldingofthegamontandthegamontnucleusnotobservablethroughthecapsule.
Scale-bar:10μm(eventhoughnotprovidedbyBall,wehaveprovidedascalematchingtotheintextdescription)Cooketal.
Parasites&Vectors(2016)9:347Page6of13overonitself(Figs.
1dand3c).
Otherwise,theonlyevidenceofthisbehaviourwasacrescentshapedstainatthecentreoftheovalparasite(asseeninallthreereports)(Figs.
1,2and3).
Itisbasedontheaboveuniquecharacteristics,particularlyofthema-turegamontstages,thatwesuggestallthreereportsareofthesameparasitespeciesK.
paradoxa.
Nohapantotype,accordingtotheInternationalCodeofZoologicalNomenclature(ICZN)Article73.
3,wasdesignatedanddepositedduringtheoriginaldescriptionofK.
paradoxabyDias[9].
Ball[30]alsodidnotidentifytheparasitetotaxonlevelanddidnotdepositanyvouchermaterial.
Furthermore,oureffortstolocateanyoriginalspecimensorvouchermaterialwereunsuccessful.
Inthisstudy,K.
paradoxawascollectedfromNdumoGameReserve,northernKwaZulu-Natal,SouthAfrica,borderingthesouthofMozambique.
TheoriginaldescriptionbyDias[9]wascollectedinthevicinityofMaputointhesouthernpartsofMozambiqueapproximately300kmfromtheNGR.
Additionally,K.
paradoxainthepresentstudywascol-lectedfromthesamehostspeciesVaranusalbigularisalbigularisasintheoriginaldescriptionbyDias[9].
Basedontheabove,thematuregamontsizecomparisons,andtheuniquecharacteristicsofthematuregamontsofK.
paradoxa(destructionandshrinkageoftheinfectedhosterythrocyte,destructionofthehostcellnucleusresultinginvacuolation,andthethicknon-stainingcapsule)asdescribedaboveforallthreereportsofthisparasite,whichincludestheoriginaldescriptionbyDias[9],andinaccordancewithICZNArticle75.
3,wedesignateaneohapantotype.
Thepresentstudyalsoincludesboththedescriptionofanadditionalstageoftheparasite(atrophozoite)andprovidessequencedata(fragment18SrDNA),whichwasnotprovidedbyDias[9]inhisori-ginaldescriptionofthisparasitespecies.
Thisneoha-pantotypeisdepositedintheprotozoancollectionoftheNationalMuseum,Bloemfontein,SouthAfricaunderaccessionnumberNMBP410.
SequenceidentificationandphylogeneticanalysisAmpliconsof611and613ntforthe18SrRNAgeneofK.
paradoxawereobtainedfromtheV.
a.
albigulariswiththeseeminglypureandhighestparasitaemiainfec-tionusingprimersetsHepF300andHepR900.
NoK.
paradoxaisolatewasobtainedfromtheV.
niloticus,likelyduetoitslowparasitaemiaincomparisontotheconcurrentinfectionoftheotherunidentifiedHepato-zoonspecies(unpublisheddata).
Thedetailsofthespe-ciesusedinthephylogeneticanalysesandpresentedintheconsensustreeareprovidedinTable2.
ThetopologyofboththeBIandMLanalyseswereoverallsimilar,withdiscretemonophyleticcladesofknownandlikelybelong-ingtoKaryolysusspecies,Hepatozoonspp.
ofmammals('intraleucocytic'Hepatozoonspp.
),theherpatofauna('intraerythrocytic'Hepatozoonspp.
),Hemoliviaspp.
,Haemogregarinaspp.
,andtheDactylosomatidae(Fig.
4).
OurresultsshowedthatK.
paradoxaclusteredwithinamajormonophyleticcladecontainingbothknown(mor-phologicallyandecologicallyconfirmed)andlikelybe-longingtoKaryolysusspecies,and'intraleucocytic'HepatozoonspeciesofmammalsthatweresistertotheKaryolysusclade.
Thismajorcladewasdistinctfromthemajormonophyleticcladecontainingtheherpatofaunal'intraerythrocytic'Hepatozoonspp.
andHemoliviaspp.
Theparasitecollectedinthepresentstudythatmor-phologicallyconformedtotheoriginaldescriptionofHepatozoonparadoxaformedasistertaxontothelarger'Karyolysus'clade(containingtheknownandlikelybe-longingtoKaryolysusspecies)inthephylogeneticana-lysis(Fig.
4).
AccordingtoevolutionarydivergenceestimatesthepresentmaterialwasmostcloselyrelatedtoknownKaryolysusspp.
(at97.
7%,p-distance=0.
02)thantospeciesofthegeneraHepatozoon,HemoliviaandHaemogregarina(Table3).
DiscussionVaranusalbigularisalbigularisandVaranusniloticusareknowntodisplaysomewhatdifferenthabitatprefer-ences,preferringmoreterrestrialandaquaticenviron-mentsrespectively[32].
BothV.
a.
albigularisandV.
niloticuscanbefoundthroughoutSouthAfricafromthemoretropicalIndianOceancoastalbeltintheEast,westtothemarginsofthemorearidNorthernandWesternCapeprovinces[32].
Bothspeciesoftenoccursympatri-cally,particularlyinNdumoGameReserve,renderingthefindingofthesameparasiteinboththesespeciesnotsurprising.
AsthemorphologicalanddevelopmentalcharacteristicstypicalofHaemogregarinaspp.
hadneverbeenobservedinhaemogregarinesoftheherpatofauna,K.
paradoxawastransferredalongwithmanyotherspeciesfromtheherpa-tofauna,birdsandmammals,fromthegenusHaemogre-garina(Haemogregarinidae)tothegenusHepatozoon(Hepatozoidae)bySmith[3]duringasystematicreviewoftheHepatozoidae.
However,duringthefirstdetailedrevi-sionofK.
paradoxaprovidedhereinthepresentstudy,morphologicallythisspeciessharesmorecharacteristicswithmembersofthefamilyKaryolysidae.
Karyolysusparadoxaperipheralbloodgamontsappeartobeencapsu-latedasintheHemoliviaanddestroythehostcellnucleusasinanumberofspeciesofKaryolysus(asmentionedaboveinRemarks)[1,7].
However,itisimperativetotakeintoaccountthatthesemorphologicalfeatures,specificallyforthelattergenus,arenotalwayspresent[7].
ThephylogeneticpositionofK.
paradoxawasshowntobeatthebaseofacladecontainingunde-scribedspeciesofHepatozoon,manyofthesetheCooketal.
Parasites&Vectors(2016)9:347Page7of13Table2Listoforganismsusedinthephylogeneticanalysesofthisstudy,withassociatedhost,hostfamilyandhostcommonname,GenBankaccessionnumbersandreferencesCladeOrganismHostFamilyCommonnameAccessionnumberReferenceKaryolysusKaryolysuslatusPodarcismuralisLacertidaeCommonwalllizardKJ461939[7]Hepatozoonsp.
AlgyroidesmarchiLacertidaeSpanishkeeledlizardJX531933[34]Hepatozoonsp.
PodarcisvaucheriLacertidaeAndalusianwalllizardHQ734803[33]KaryolysuslacazeiLacertatrilineataLacertidaeBalkanemeraldlizardKJ461942[7]Karyolysussp.
ZootocaviviparaLacertidaeViviparouslizardKJ461945[7]Hepatozoonsp.
PodarcisbocageiLacertidaeBocage'swalllizardJX531952[34]Hepatozoonsp.
PodarcisbocageiLacertidaeBocage'swalllizardJX531954[34]Hepatozoonsp.
TimontangitanusLacertidaeMoroccaneyedlizardHQ734799[33]Hepatozoonsp.
AtlantolacertaandreanskyiLacertidaeAndreansky'slizardHQ734798[33]Karyolysussp.
Ophionyssussp.
exLacertaviridisLacertidaeEuropeangreenlizardKJ461944[7]Hepatozoonsp.
PsammophisschokariLamprophiidaeSchokarisandracerKC696565[47]Hepatozoonsp.
HemorrhoishippocrepisColubridaeHorseshoesnakeJX244269[35]Hepatozoonsp.
EumecesalgeriensisScincidaeAlgerianskinkHQ734796[33]Hepatozoonsp.
ScelarcisperspicillataLacertidaeMenorcawalllizardHQ734791[33]Hepatozoonsp.
PodarcisvaucheriLacertidaeAndalusianwalllizardHQ734804[33]Hepatozoonsp.
PodarcishispanicusLacertidaeIberianwalllizardJX531917[34]Hepatozoonsp.
PodarcislilfordiLacertidaeLilford'swalllizardJX531920[34]Karyolysus(syn.
Hepatozoon)paradoxaVaranusalbigularisVaranidaeRockmonitorlizardKX011039ThisstudyKaryolysus(syn.
Hepatozoon)paradoxaVaranusalbigularisVaranidaeRockmonitorlizardKX011040ThisstudyIntraleucocyticHepatozoonHepatozoonamericanumCanisfamiliarisCanidaeDomesticdogAF176836[48]HepatozooncanisCanisfamiliarisCanidaeDomesticdogAY461378[49]HepatozoonursiUrsusthibetanusjaponicusUrsidaeJapaneseblackbearEU041718[50]Hepatozoonsp.
MartesmartesMustelidaePinemartenEF222257[51]HepatozoonfelisFeliscatusFelidaeDomesticcatAY620232[50]HepatozoonHepatozooncatesbianaeLithobatescatesbeianusRanidaeAmericanbullfrogHQ224954[49]HepatozoonixoxoSclerophrys(syn.
Amietophrynus)maculatusBufonidaeFlat-backedtoadKP119772[52]HepatozoontheileriAmietiaqueckettiPyxicephalidaeQueckett'sriverfrogKP119773[18]HepatozoonsipedonNerodiasipedonColubridaeNorthernwatersnakeJN181157[49]HepatozoonfitzsimonsiKinixyszombensisTestudinidaeBell'shingebacktortoiseKR069084[12]Cooketal.
Parasites&Vectors(2016)9:347Page8of13Table2Listoforganismsusedinthephylogeneticanalysesofthisstudy,withassociatedhost,hostfamilyandhostcommonname,GenBankaccessionnumbersandreferences(Continued)CladeOrganismHostFamilyCommonnameAccessionnumberReferenceHepatozoonayorgborPythonregius(aLamprophis(syn.
Boaedon)fuliginosus)Pythonidae;aLamprophiidaeBallpython;brownhousesnakeEF157822[53]HepatozoondomergueiFurcifersp.
ChamaeleonidaeChameleonspeciesKM234649[54]HepatozoonseychellensisGrandisoniaalternansIndotyphlidaeCaecilianKF246566[55]HemoliviaHemoliviamauritanicaTestudograecaTestudinidaeMediterraneanspur-thighedtortoiseKF992700[36]HemoliviaparvulaKinixyszombensisTestudinidaeBell'shingebacktortoiseKR069082[12]HemoliviamariaeEgerniastokesiiScincidaeGidgeespiny-tailedskinkKF992711[36]Hemoliviasp.
RhinoclemmyspulcherrimamanniGeoemydidaePaintedwoodturtleKF992714[36]HemoliviastellataAmblyommarotundatumexRhinellamarinaBufonidaeCanetoadKP881349[37]HaemogregarinidaeHaemogregarinasp.
_3140PelusiossubnigerPelomedusidaeEastAfricanblackmudturtleKF257925[56]HaemogregarinasacaliaeSacaliaquadriocellataGeoemydidaeFour-eyedturtleKM887507[57]HaemogregarinapellegriniMalayemyssubtrijugaGeoemydidaeSnail-eatingturtleKM887508[57]HaemogregarinaballiChelydraserpentinaserpentinaChelydridaeCommonsnappingturtleHQ224959[49]HaemogregarinastepanowiMauremyscaspicaGeoemydidaeCaspianturtleKF257926[56]DactylosomatidaeBabesiosomastableriLithobatesseptentrionalisRanidaeMinkfrogHQ224961[49]DactylosomaranarumPelophylaxlessonae(syn.
esculentus)RanidaePoolfrogHQ224957[49]OutgroupKlossiahelicinaCepaeanemoralisHelicidaeGrovesnailHQ224955[49]AdelinadimidiataScolopendracingulataScolopendridaeMegarianbandedcentipedeDQ096835[58]AdelinagrylliGryllusbimaculatusGryllidaeFieldcricketDQ096836[58]aExperimentallaboratoryanimalCooketal.
Parasites&Vectors(2016)9:347Page9of13resultsofmolecularHepatozoonspp.
surveys[33,34],andknownspeciesofKaryolysus[7].
Thismaysug-gestthattheseundescribedHepatozoonspp.
mightra-therbespeciesofKaryolysus.
TheKaryolysusspp.
cladeispartofalargercladeincludingasistercladeofHepatozoonspp.
frommammals.
Thistopology,asseeninthisstudy,hasbeenobservedinanumberofotherstudies[7,33–37].
Karadjianetal.
[37],intheirattempttounderstandtherelationshipsofthediffer-enthaemogregarinegenera,particularlyinrespecttotheconundrumofthepolyphyleticHepatozoonclade,proposed,basedontheirphylogeneticfindings,thatanumberofHepatozoonspecies(mostofwhichhavenotbeenmorphologicallydescribed)mightratherrep-resentspeciesofKaryolysus.
Giventhatitwasonlysincetherecentidentification,descriptionandmolecularchar-acterisationofKaryolysusspp.
by[7],itisonlynowthatwearebeginningtorealizethispossibility.
Moreover,itappearsthatthediversityofsquamatesparasitisedbylikelyspeciesofKaryolysusisfastincreasing,ascenarioFig.
4PhylogeneticanalysisofKaryolysusparadoxa(Dias,1954)basedon18SrDNAsequences.
Bayesianinference(BI)andMaximumLikelihood(ML)analysisshowingthephylogeneticrelationshipsfortwoKaryolyusparadoxaisolatesfromthecurrentstudyrepresentedinbold,17KaryolysusandHepatozoonspecies(representingtheKaryolysuscladeinblue),fivemammal-infectingHepatozoonspecies(representingthe'intraleucocytic'Hepatozooncladeinpurple),eightherpatofaunal-infectingHepatozoonspecies(representingthe'intraerythrocytic'Hepatozooncladeingreen),fiveHemoliviaspecies(representingtheHemoliviacladeinorange),fiveHaemogregarinaspecies(representingtheHaemogregarinidaecladeinyellow),aBabesiosomaandDactylosomaspecies(representingtheDactylosomatidaecladeinpink)andtwoAdelinaandoneKlossiaspecies(usedastheoutgroupingrey).
AllcomparativesequencesweredownloadedfromtheGenBankdatabase.
TreetopologiesforboththeBIandMLtreeswereidentical;thenodalsupportvalues(BI/ML)arerepresentedontheBItreeCooketal.
Parasites&Vectors(2016)9:347Page10of13whichislikelyonlytointensifyinfuture.
Arecentmo-lecularHepatozoonspp.
surveyby[38]showshaemogre-garineisolatesfromgeckosofthegenusTarentolaGray,1825(Phyllodactylidae)arealsofallingwithinwhatmaybeseenasthe'Karyolysus'clade;acladethatatpresentalsoincludeshaemogregarinesisolatedfromspeciesofthefamiliesColubridaeandLamprophiidae(bothsnakes),LacertidaeandVaranidae(bothlizards),andScincidae(skinks).
Furthermore,thepresentstudyshowsthatK.
paradoxaismostcloselyrelatedtoknownKaryolysusspecies,followedbyspeciesofthe'intraleucocytic'Hepatozooncladeandthenspeciesofthe'intraerythrocytic'Hepato-zoonandHemoliviaclades.
KaryolysusandHemoliviamorphologicallystillbelongwithinthesamefamily(Karyolisidae),however,withtheuseofthe18SrRNAgenethesetwogenerainthisstudyandinothersgenerallyfallindifferentmajorclades.
Karyolysusclustersinamajorcladewiththe'intraleucocytic'mammalHepatozoon,whilstHemoliviaclustersinamajorcladewiththe'intraerythrocytic'herpatofaunalHepatozoon;thisfindingisapparentinthepresentstudy,andalsoin[36,37].
Itisclearthattherelationshipbetweenthesetwogeneramaypossiblyonlyberesolvedbyusingamulti-geneapproachasin[39].
ConclusionsBasedonthemorphologyandthemolecularfindingspresentedinthisstudy,werecommendthefollowingnomenclaturalcorrection:Karyolysusparadoxa(Dias,1954)(syn.
Hepatozoonparadoxa(Dias,1954)Smith,1996,HaemogregarinaparadoxaDias,1954)inthevara-nidlizardsVaranusalbigularisalbigularis(type-host),andVaranusniloticus.
OurresultsshowedthatKaryoly-susparadoxaisascloselyrelatedtospecieswithinitscurrentgenericassignmentinthe'intraerythrocytic'her-patofaunalHepatozoonasitiswiththemoredistantlyrelatedspeciesoftheHaemogregarina.
Besidesthisstudyrepresentingthefirstmorpho-logicalandmolecularreportofahaemogregarinewithinanAfricanvaranid,itisthefirstreportofaspe-ciesofKaryolysusinfectingahostoftheVaranidae.
Furthermore,itrepresentsthethirddescribedandnamedKaryolysusspp.
forwhichthereisanucleotidesequenceavailable.
Itishopedthatthisstudywillen-couragefurthermolecularworkontheKaryolysidae,particularlythegenusKaryolysus.
ThisstudyalsoextendsthehostanddistributionrangeofK.
paradoxafromonlyasinglespecimenofV.
a.
albigularisinMozambiquetoanadditionaltwospecimensinSouthAfrica,aswellasincludingV.
niloticusasanadditionalhostbothinSouthAfricaandKenya.
ThedistributionrangeofK.
paradoxafallswithinsubtropicalareasinSouthAfrica,MozambiqueandKenya,andassuchitwouldbein-terestingtoseeifthisparticularparasiteisrestrictedtosubtropicalareassuchasisthecasewithHemoliviaparvula(Dias,1953)foundparasitisingKinixyszombensisHewitt,1931tortoisesofSouthAfricaandMozambique;see[12],orifitismorewidelydistributedthroughoutdif-ferentbiomesasisthecasewithHepatozoonfitzsimonsi(Dias,1953)foundparasitisingseveraltortoisespeciesinSouthAfricaandMozambique;see[19].
Eventhoughticksquashesdidnotresultinanyob-servableparasiticstagesfuturestudieswillfocusonidentifyingpossiblevectors.
Parasiticstagesfoundinthesepossiblevectorswillbeidentifiedtospecieslevelbasedonbothmorphologicalandmolecularfindings.
WiththeconundrumofthelargerHepatozooncladebeingpolyphyleticandabsorbingtheHemoliviaandKaryolysus,itisimportanttoincreasethenumberoftaxafromwhichwecanworkandaskdeeperphylogen-eticquestions.
However,besidesthemolecularcharac-terisationofthesespecies,itisstillimportanttofocusontheirmorphologyandwherepossibleattempttoelu-cidatetheirlife-cyclesinordertoresolvethecomplextaxonomyoftheseorganisms.
Moreimportantly,itisnecessarytoincludeanotherfasterevolvinggeneorevenmitochondrialgenomesofthesegroupsfollowing[39]beforewecanmakeanywell-informeddecisions.
AcknowledgementsWewouldliketothankNdumoGameReserveandEzemveloKZNWildlife,foraccesstositesandresearchpermits(OP839/2014;OP1262/2015;OP2492/2015)forsamplecollection.
Inaddition,wewouldliketothankTable3RepresentationofevolutionarydivergenceofthedifferentcladesinrelationtoKaryolysusparadoxa(Dias,1954)CladeSimilarityp-distanceNucleotidedifferenceParsimonyinformativesites(%)KnownKaryolysus(4)97.
70.
0214(11–18)73'Intraleucocytic'Hepatozoon(5)95.
70.
04(0.
03–0.
05)26(18–45)29Hepatozoon(8)94.
40.
05(0.
03–0.
09)33(21–56)46Hemolivia(5)97.
00.
0318(17–20)66Haemogregarina(5)94.
70.
0532(29–34)68SummativerepresentationofcladesshowninFig.
4,showingcladeswithnumberofsequencesinparentheses,theaveragepercentsimilarity,averageuncorrectedp-distancewithrangeinparentheses,averagebasepairdifferencewithrangeinparentheses,andpercentparsimonyinformativesites,allcomparedtothetwosequencesasshowninFig.
4ofKaryolysus(syn.
Hepatozoon)paradoxafromVaranusalbigularisalbigularis.
NotethatonlytheknownKaryolysusspecieshavebeenusedfromtheKaryolysuscladeCooketal.
Parasites&Vectors(2016)9:347Page11of13Microbiology,UnitforEnvironmentalSciencesandManagement,(NWU-PC),fortheuseoftheirfacilities.
ThefinancialassistanceoftheNationalResearchFoundation(NRF)ofSouthAfricatoCAC(NRFScarceSkillsPostdoctoralScholarship-GrantSFP13090332476),andtheFlemishInteruniversityCouncil(VLIR)toECN(VLIR-OUSproject–ZEIN21013PR396)isacknowledged.
Opinionsexpressedandconclusionsarrivedat,arethoseoftheauthorsandarenotnecessarilytobeattributedtotheNRForVLIR.
Authors'contributionsAllauthorsconceivedanddesignedtheproject,participatedingeneraldataanalysisandindraftingthemanuscript.
CACandECNcarriedoutthefieldwork,preparedandexaminedbloodsmears,preparedlightmicrographsandcompiledallmeasurementdata.
ECNparticipatedinthemolecularstudiesandinthesequencealignmentandprovidedsupporttothepreparationofthemanuscript.
Allauthorsreadandapprovedthefinalmanuscript.
CompetinginterestsTheauthorsdeclarethattheyhavenocompetinginterests.
Authordetails1UnitforEnvironmentalSciencesandManagement,North-WestUniversity,Potchefstroom,SouthAfrica.
2LaboratoryofAquaticEcology,EvolutionandConservation,UniversityofLeuven,3000Leuven,Belgium.
Received:10March2016Accepted:18May2016References1.
TelfordSR.
HemoparasitesoftheReptilia:ColorAtlasandText.
NewYork:CRCPress;2009.
2.
SmallridgeCJ,PapernaI.
Thetick-transmittedhaemogregarinidoftheAustraliansleepylizardTiliquarugosabelongstothegenusHemolivia.
Parasite.
1997;4:359–63.
3.
SmithTG.
ThegenusHepatozoon(Apicomplexa:Adeleina).
JParasitol.
1996;82:565–85.
4.
MerinoS,MartínezJ,MaselloJF,BedollaY,GuillfeldtP.
FirstmolecularcharacterizationofaHepatozoonspecies(Apicomplexa:Hepatozoidae)infectingbirdsanddescriptionofanewspeciesinfectingstormpetrels(Aves:Hydrobatidae).
JParasitol.
2014;100:338–43.
5.
DaviesAJ,JohnstonMRL.
Thebiologyofsomeintraerythrocyticparasitesoffishes,amphibiansandreptiles.
AdvParasitol.
2000;45:1–107.
6.
VanAsJ,DaviesAJ,SmitNJ.
Hepatozoonlangiin.
sp.
andHepatozoonvacuolatusn.
sp.
(Apicomplexa:Adeleorina:Hepatozoidae)fromthecraglizard(Sauria:Cordylidae)PseudocordyluslangifromtheNorthEasternDrakensbergescarpment,EasternFreeState,SouthAfrica.
Zootaxa.
2013;3608:345–56.
7.
Haklová-KoíkováB,HiňanováA,MajláthI,RakaK,HarrisDJ,FldváriG,TryjanowskiP,KokoováN,MalekováB,MajláthováV.
MorphologicalandmolecularcharacterizationofKaryolysus-aneglectedbutcommonparasiteinfectingsomeEuropeanlizards.
ParasitVectors.
2014;7:555.
8.
VanAsJ,DaviesAJ,SmitNJ.
LifecycleofHepatozoonaffluomalotisp.
n.
(Apicomplexa:Haemogregarinidae)incraglizards(Sauria:Cordylidae)andinculicinemosquitoesfromSouthAfrica.
FoliaParasitol.
2015;62:008.
9.
DiasJATS.
SubsídiosparaoestudodoshematozoáriosdosrépteisdeMoambique3:DuasnovasespéciesdeHaemogregarinaparasitasdoVaranusalbigularisalbigularis(Daudin,1802).
BolSocEstMo.
1954;87:71–9.
10.
NetherlandsEC,CookCA,KrugerDJ,duPreezLH,SmitNJ.
Biodiversityoffroghaemoparasitesfromsub-tropicalnorthernKwazulu-Natal,SouthAfrica.
IntJParasitolParasitesWildl.
2015;4:135–41.
11.
CookCA,SmitNJ,DaviesAJ.
Firstrecordofanintraleucocytichaemogregarine(Adeleorina:Haemogregarinidae)fromSouthAfricantortoisesofthespeciesStigmochelyspardalis(Cryptodira:Testudinidae).
AfrZool.
2014;49:290–4.
12.
CookCA,NetherlandsEC,SmitNJ.
FirstHemoliviafromsouthernAfrica:reassigningchelonianHaemogregarinaparvulaDias,1953(Adeleorina:Haemogregarinidae)toHemolivia(Adeleorina:Karyolysidae).
AfrZool.
2015;50:165–73.
13.
LaveranA.
SurunehaemogregarinedeVaranusniloticus.
CRSeancesSocBiol.
1905;59:175–6.
14.
NicolleMMC,ComteC.
SurunehémogrégarinedeVaranusgriseus.
CRSeancesSocBiol.
1906;61:310–2.
15.
WolbachSB.
Notesonthelifecycleofahemogregarinefoundinamonitor(Veranusniloticus).
JMedRes.
1914;29:473–88.
16.
CookCA,SmitNJ,DaviesAJ.
AredescriptionofHaemogregarinafitzsimonsiDias,1953andsomecommentsonHaemogregarinaparvulaDias,1953(Adeleorina:Haemogregarinidae)fromsouthernAfricantortoises(Cryptodira:Testudinidae),withnewhostdataanddistributionrecords.
FoliaParasitol.
2009;56:173–9.
17.
CookCA,SmitNJ,DaviesAJ.
Hemproteids(Apicomplexa:Haemoproteidae)fromSouthAfricanTortoises(Cryptodiria:Testudinidae).
JParasitol.
2010;96:1168–72.
18.
NetherlandsEC,CookCA,SmitNJ,duPreezLH.
RedescriptionandmoleculardiagnosisofHepatozoontheileri(Laveran,1905)(Apicomplexa:Adeleorina:Hepatozoidae),infectingAmietiaquecketti(Anura:Pyxicephalidae).
FoliaParasitol.
2014;61:293–300.
19.
CookCA,LawtonSP,DaviesAJ,SmitNJ.
ReassignmentofthelandtortoisehaemogregarineHaemogregarinafitzsimonsiDias1953,(Adeleorina:Haemogregarinidae)tothegenusHepatozoonMiller1908(Adeleorina:Hepatozoidae)basedonparasitemorphology,lifecycleandphylogeneticanalysisof18SrDNAsequencefragments.
Parasitology.
2014;141:1611–20.
20.
UjvariB,MadsenT,OlssonM.
HighprevalenceofHepatozoonspp.
(Apicomplexa:Hepatozoidae)infectioninwaterpythons(Liasisfuscus)fromtropicalAustralia.
JParasitol.
2004;90:670–2.
21.
GeneioussoftwarecreatedbyBiomatters.
[http://www.
geneious.
com].
22.
BasicLocalAlignmentSearchTool.
[http://blast.
ncbi.
nlm.
nih.
gov/Blast.
cgi].
23.
EdgarRC.
Muscle:multiplesequencealignmentwithhighaccuracyandhighthroughput.
NucleicAcidsRes.
2004;5:1792–7.
24.
TamuraK,StecherG,PetersonD,FilipskiA,KumarS.
MEGA6:molecularevolutionarygeneticsanalysisVersion6.
0.
MolBiolEvol.
2013;30:2725–9.
25.
GuindonS,GascuelO.
Asimple,fastandaccuratemethodtoestimatelargephylogeniesbymaximum-likelihood.
SystBiol.
2003;52:696–704.
26.
DarribaD,TaboadaGL,DoalloR,PosadaD.
jModelTest2:moremodels,newheuristicsandparallelcomputing.
NatMethods.
2012;9:772.
27.
HuelsenbeckJP,RonquistF.
MrBayes:Bayesianinferenceofphylogenetictrees.
Bioinformatics.
2001;17:754–5.
28.
StamatakisA.
RAxMLversion8:atoolforphylogeneticanalysisandpost-analysisoflargephylogenies.
Bioinformatics.
2014;30:1312–3.
29.
SilvestroD,MichalakI.
raxmlGUI:agraphicalfront-endforRAxML.
OrgDiversEvol.
2012;12:335–7.
30.
BallGH.
SomebloodsporozoafromEastAfricanreptiles.
JProtozool.
1967;14:198–210.
31.
irokP,KamlerM,FryeFL,FictumP,ModrD.
EndogenousdevelopmentofHemoliviamauritanica(Apicomplexa:Adeleina:Haemogregarinidae)inthemarginatedtortoiseTestudomarginata(Reptilia:Testudinidae):evidencefromexperimentalinfection.
FoliaParasitol.
2007;54:13–8.
32.
BatesMF,BranchWR,BauerAM,BurgerM,MaraisJ,AlexanderGJ,deVilliersMS.
AtlasandredlistofthereptilesofSouthAfrica,LesothoandSwaziland.
Suricata1.
Pretoria:SouthAfricanNationalBiodiversityInstitute;2014.
33.
MaiaJPMC,HarrisDJ,PereraA.
MolecularsurveyofHepatozoonspeciesinlizardsfromNorthAfrica.
JParasitol.
2011;97:513–7.
34.
MaiaJPMC,PereraA,HarrisDJ.
MolecularsurveyandmicroscopicexaminationofHepatozoonMiller,1908(Apicomplexa:Adeleorina)inlacertidlizardsfromthewesternMediterranean.
FoliaParasitol.
2012;59:241–8.
35.
ToméB,MaiaJPMC,HarrisDJ.
Hepatozooninfectionprevalenceinfoursnakegenera:influenceofdiet,preyparasitaemialevelsorparasitetypeJParasitol.
2012;98:913–7.
36.
KvierováJ,HypaV,DvoákováN,MikulíekP,JandzikD,GeorgeGardnerM,JavanbakhtH,TiarG,irokP.
HemoliviaandHepatozoon:haemogregarineswithtangledevolutionaryrelationships.
Protist.
2014;165:688–700.
37.
KaradjianG,ChavatteJ,LandauI.
Systematicrevisionoftheadeleidhaemogregarines,withcreationofBartazoonn.
g.
,reassignmentofHepatozoonargantisGarnham,1954toHemolivia,andmoleculardataonHemoliviastellata.
Parasite.
2015;22:31.
38.
ToméB,RatoC,PereraA,HarrisDJ.
HighdiversityofHepatozoonspp.
ingeckosofthegenusTarentola.
JParasitol.
2016.
doi:10.
1645/15-908.
39.
OgedengbeJD,OgedengbeME,HafeezMA,BartaJR.
Molecularphylogeneticsofeimeriidcoccidian(Eimeriidae,Eimeriorina,Apicomplexa,Alveolata):Apreliminarymulti-geneandmulti-genomeapproach.
ParasitolRes.
2015;114:4149–41460.
40.
RamdanNF,SaoudMFA,MohammedSH,FawziSM.
OnanewhaemogregarineofVaranusgriseusfromEgypt.
QatarUnivSciJ.
1996;16:119–25.
Cooketal.
Parasites&Vectors(2016)9:347Page12of1341.
LaveranA,PettitA.
Contributionàl'étudedeshaemogregarinesdequelquessauriensd'Afrique.
BullSocPatholExot.
1909;2:506–14.
42.
FranaC.
NotessurdeshématozoairesdelaGuinéePortugaise.
ArqivRealInstitutdeBactériologieCmaraPestana.
1911;3:229–38.
43.
ToddJL,WolbachSB.
ParasiticprotozoafromTheGambia1912:secondreportoftheexpeditionoftheLiverpoolSchoolofTropicalMedicinetoTheGambia,1911.
JMedRes.
1912;26:195–218.
44.
LegerM,LegerA.
HématozoairesdesreptilesduHaut-Senegal-Niger.
BullSocPatholExot.
1914;7:488–92.
45.
RousselotR.
1943CitedbyRousselot,R.
1953.
46.
RousselotR.
NotesdeParasitologieTropicale.
I.
ParasitesdeSangdesAnimaux.
Paris:VigotFreres;1953.
47.
ToméB,MaiaJPMC,HarrisDJ.
MolecularassessmentofapicomplexanparasitesinthesnakePsammophisfromNorthAfrica:domultipleparasitelineagesreflectthefinalvertebratehostdietJParasitol.
2013;99:883–7.
48.
MathewJS,vandenBusscheRA,EwingSA,MalayerJR,LathaBR,PancieraRJ.
PhylogeneticrelationshipsofHepatozoon(Apicomplexa:Adeleorina)basedonmolecular,morphologic,andlife-cyclecharacters.
JParasitol.
2000;86:366–72.
49.
BartaJR,OgendengbeJD,MartinDS,SmithTG.
Phylogeneticpositionoftheadeleorinidcoccidia(Myzozoa,Apicomplexa,Coccidia,Eucoccidiorida,Adeleorina)inferredusing18SrDNAsequences.
JEukaryotMicrobiol.
2012;59:171–80.
50.
Criado-FornelioA,RuasJL,CasadoN,FariasNA,SoaresMP,etal.
NewmoleculardataonmammalianHepatozoonspecies(Apicomplexa:Adeleorina)fromBrazilandSpain.
IntJParasitol.
2006;92:93–9.
51.
Criado-FornelioA,BulingA,CasadoN,GimenezC,RuasJ,etal.
Molecularcharacterizationofarthropod-bornehaematozoansinwildmammalsfromBrazil,VenezuelaandSpain.
ActaParasitol.
2009;54:187–93.
52.
NetherlandsEC,CookCA,SmitNJ.
Hepatozoonspecies(Adeleorina:Hepatozoidae)ofAfricanbufonids,withmorphologicaldescriptionandmoleculardiagnosisofHepatozoonixoxosp.
nov.
parasitisingthreeAmietophrynusspecies(Anura:Bufonidae).
ParasitVectors.
2014;7:552.
53.
SlobodaM,KamlerM,BulantováJ,VotpkaJ,ModrD.
AnewspeciesofHepatozoon(Apicomplexa:Adeleorina)fromPythonregius(Serpentes:Pythonidae)anditsexperimentaltransmissionbyamosquitovector.
JParasitol.
2007;93:1189–98.
54.
MaiaJP,CrottiniA,HarrisDJ.
MicroscopicandmolecularcharacterizationofHepatozoondomerguei(Apicomplexa)andFoleyellafurcata(Nematoda)inwildendemicreptilesfromMadagascar.
Parasite.
2014;21:47.
55.
HarrisDJ,Damas-MoreiraI,MaiaJPMC,PereraA.
FirstreportofHepatozoon(Apicomplexa:Adeleorina)incaecilians,withdescriptionofanewspecies.
JParasitol.
2014;100:117–20.
56.
DvoákováN,KvierováJ,PapouekI,JavanbakhtH,TiarG,KamiH,irokP.
HaemogregarinesfromwesternPalaearcticfreshwaterturtles(generaEmys,Mauremys)areconspecificwithHaemogregarinastepanowiDanilewsky,1885.
Parasitology.
2013;141:522–30.
57.
DvoákováN,KvierováJ,HostovskM,irokP.
HaemogregarinesoffreshwaterturtlesfromSoutheastAsiawithadescriptionofHaemogregarinasacaliaesp.
n.
andaredescriptionofHaemogregarinapellegriniLaveranandPettit,1910.
Parasitology.
2015;142:816–26.
58.
KopenáJ,JirkM,OborníkM,TokarevYS,LukeJ,ModrD.
Phylogeneticanalysisofcoccidianparasitesfrominvertebrates:searchformissinglinks.
Protist.
2006;157:173–83.
Weacceptpre-submissioninquiriesOurselectortoolhelpsyoutondthemostrelevantjournalWeprovideroundtheclockcustomersupportConvenientonlinesubmissionThoroughpeerreviewInclusioninPubMedandallmajorindexingservicesMaximumvisibilityforyourresearchSubmityourmanuscriptatwww.
biomedcentral.
com/submitSubmityournextmanuscripttoBioMedCentralandwewillhelpyouateverystep:Cooketal.
Parasites&Vectors(2016)9:347Page13of13
comb.
(Karyolysidae)CourtneyA.
Cook1*,EdwardC.
Netherlands1,2andNicoJ.
Smit1AbstractBackground:WithintheAfricanmonitorlizardfamilyVaranidae,twohaemogregarinegenerahavebeenreported.
ThesecomprisefivespeciesofHepatozoonMiller,1908andaspeciesofHaemogregarinaDanilewsky,1885.
EventhoughotherhaemogregarinegenerasuchasHemoliviaPetit,Landau,Baccam&Lainson,1990andKaryolysusLabbé,1894havebeenreportedparasitisingotherlizardfamilies,thesehavenotbeenfoundinfectingtheVaranidae.
ThegenusKaryolysushastodatebeenformallydescribedandnamedonlyfromlizardsofthefamilyLacertidaeandtotheauthors'knowledge,thisincludesonlyninespecies.
Molecularcharacterisationusingfragmentsofthe18Sgenehasonlyrecentlybeencompletedforbuttwoofthesespecies.
Todate,threeHepatozoonspeciesareknownfromsouthernAfricanvaranids,oneoftheseHepatozoonparadoxa(Dias,1954)sharesmorphologicalcharacteristicsaliketospeciesofthefamilyKaryolysidae.
Thus,thisstudyaimedtomorphologicallyredescribeandcharacteriseH.
paradoxamolecularly,soastodetermineitstaxonomicplacement.
Methods:SpecimensofVaranusalbigularisalbigularisDaudin,1802(Rockmonitor)andVaranusniloticus(LinnaeusinHasselquist,1762)(Nilemonitor)werecollectedfromtheNdumoGameReserve,SouthAfrica.
Uponcaptureanimalswereexaminedforhaematophagousarthropods.
Bloodwascollected,thinbloodsmearsprepared,stainedwithGiemsa,screenedandmicrographsofparasitescaptured.
HaemogregarinemorphometricdatawerecomparedwiththedatafornamedhaemogregarinesofAfricanvaranids.
PrimersetHepF300andHepR900wasemployedtotargetafragmentofthe18SrRNAgeneandresultingsequencescomparedwithotherknownhaemogregarinesequencesselectedfromtheGenBankdatabase.
Results:Hepatozoonparadoxawasidentifiedinfectingtwooutofeight(25%)V.
a.
albigularisandasingle(100%)V.
niloticusexamined.
PhylogeneticanalysesrevealedthatH.
paradoxaclusteredwiththe'Karyolysus'clade,andnotwiththoseofreptilianHepatozoonspp.
Conclusions:InadditiontothisbeingthefirstmorphologicalandmolecularcharacterisationofahaemogregarinewithintheAfricanVaranidae,itisthefirstreportofaspeciesofKaryolysusinfectingthemonitorlizardfamily.
Furthermore,thisconstitutesnowonlythethirddescribedandnamedKaryolysusspeciesforwhichthereisanucleotidesequenceavailable.
Keywords:Haemogregarinetaxonomy,Hepatozoon,Monitorlizard,Haematozoa,Hemolivia*Correspondence:apicomplexan@yahoo.
co.
zaEqualcontributors1UnitforEnvironmentalSciencesandManagement,North-WestUniversity,Potchefstroom,SouthAfricaFulllistofauthorinformationisavailableattheendofthearticle2016TheAuthor(s).
OpenAccessThisarticleisdistributedunderthetermsoftheCreativeCommonsAttribution4.
0InternationalLicense(http://creativecommons.
org/licenses/by/4.
0/),whichpermitsunrestricteduse,distribution,andreproductioninanymedium,providedyougiveappropriatecredittotheoriginalauthor(s)andthesource,providealinktotheCreativeCommonslicense,andindicateifchangesweremade.
TheCreativeCommonsPublicDomainDedicationwaiver(http://creativecommons.
org/publicdomain/zero/1.
0/)appliestothedatamadeavailableinthisarticle,unlessotherwisestated.
Cooketal.
Parasites&Vectors(2016)9:347DOI10.
1186/s13071-016-1600-8BackgroundWithintheapicomplexanorderAdeleiorina,representa-tivesoftwohaemogregarinegenera,HepatozoonMiller,1908andKaryolysusLabbé,1894,arecommonlyre-portedinfectingsaurians.
ThegenusHemoliviaPetit,Landau,Baccam&Lainson,1990onthecontrary,eventhoughreportedparasitisingsaurianhosts,hasbutasingledescribedspeciesHemoliviamariaeSmallridge&Paperna,1997[1,2].
RepresentativesofHepatozoonarethemostcommonandarecosmopolitanparasitesfoundparasitisingawiderangeofvertebratehostsfromamphibiansandreptilestobirdsandmammals[3,4].
Karyolysus,conversely,isknownmainlyasasaurianhaemogregarinegenusthatprimarilyparasitiseslizardsofthefamilyLacertidae,buthasalsobeenreportedfromlizardsoftheScincidae[1,5–7].
Besidesthisdiscrepancyinvertebratehostpreferenceofthespeciesintheabovehaemogregarinegenera,speciesinthesegeneraalsodemonstratedifferentdevelopmentalpatterns.
Eventhoughspeciesofallthreeofthehaemogregarinegeneramaybetransmittedtothesaurianhostthroughthein-gestionoftheinfectedinvertebratevector,Hepatozoonspp.
maybetransmittedthroughawiderangeofarthro-podvectors(mosquitoestoticks),whilsttransmissionofHemoliviaspp.
andKaryolysusspp.
hasbeenrecordedonlythroughatickandmitevector,respectively[1].
Whilstmorethan30Hepatozoonspp.
havebeenre-cordedfromsauriansthroughoutAfrica[8],Karyolysusspp.
havemainlybeenreportedfromlacertidlizardsofEuropeandAsia[7].
UntilSmith's[3]revisionoftheHepatozoidae,thegenusKaryolysuscomprised11species,Smith[3]reassigningtwoofthesetoHepatozoon,nowHepatozoonberestnewi(Finkelstein,1907)andHepatozoonbicapsulata(Franca,1910).
Totheauthors'knowledge,todate,ninespeciesofKaryolysusareconsid-eredvalid:KaryolysuslacertaeDanilewsky,1886;Karyoly-suslacazeiLabbé,1894;KaryolysusbiretortusNicolle,1904;KaryolysuszuluetaiReichenow,1920;KaryolysussubtilisRicci,1954;KaryolysusoctocromosomiAlvarez-Calvo,1975;KaryolysuslatusSvahn,1975;KaryolysusminorSvahn,1975;andtheonlyspeciesreportedfromsub-SaharanAfricaKaryolysuspoleensisMutinga&Dipeolu,1989[3,6,7].
Descriptionsofhaemogregarinespecieswereuntilrecentlybasedonmorphologicalchar-acteristicsandlife-cycledata[7].
ThisisparticularlytrueofthehaemogregarinesdescribedfromsaurianswithinsouthernAfrica.
Haemogregarines,specificallyspeciesofHepatozoon,havebeencommonlyrecordedbloodpara-sitesofsouthernAfricansauriansincludingthosefromlizardgeneraofthefamiliesCordylidaeandVaranidae[8,9].
WithintheAfricanVaranidae,sixspeciesofHepa-tozoonhavebeendescribed,threeofthesefromsouthernAfrica(Table1),thelatterthreecomprisingHepatozoonvarani(Laveran,1905)fromVaranusniloticus(Linnaeus,1762)inSouthAfrica[3,6],Hepatozooncamarai(Dias,1954)andHepatozoonparadoxa(Dias,1954)fromVaranusalbigularisalbigularisDaudin,1802inMozambique[3,6].
TheaimofthepresentstudywasthustoprovideamorphologicalredescriptionofH.
paradoxaandmo-leculardataaidinginthecorrecttaxonomicplacementofthisparasite.
MethodsStudyarea,Varanusspp.
collectionandbloodpreparationSpecimensofVaranusalbigularisalbigularisandVaranusniloticuswerecollectedindaylightduringthesummermonthsofNovember2013,FebruaryandNovember2014,andFebruary2015intheNdumoGameReserve(NGR)(26°52′00.
0″S,32°15′00.
0″E),north-easternKwaZulu-Natal(KZN),SouthAfrica,borderingsouthernMozambique[10].
Lizardswererestrainedbyhandwhilstbloodandanyhaematopha-gousathropodswerecollectedinsitu.
Asmallvolumeofblood(approximatelyonedrop)wascollectedfromtheventralcaudalveinusinganappropriatelygauged(de-pendingonthesizeofthelizard)sterileneedleand1mlsyringe.
Asmallportionofthecollectedbloodwasusedtoprepare2–3duplicatethinbloodsmearsandthere-mainderdroppedintoanequalvolumeof70%ethanolforfuturemolecularanalysis.
Thinbloodsmearsonceair-driedinadustproofcontainerwerefixedinabsolutemethanolandstainedthereafterusingamodifiedsolu-tionofGiemsastain(FLUKA,Sigma-Aldrich,Steinheim,Germany)accordingtothemethodsof[11,12].
ScreeningofVaranusspp.
bloodsmearsSmearswerescreenedundera100*oilimmersionob-jectiveonaNikonEclipseE800compoundmicroscope(Nikon,Amsterdam,TheNetherlands)andimageswerecapturedwithanattachedNikondigitalcameraandac-companyingsoftware.
HaemogregarineswereidentifiedtospecieslevelbycomparingmorphometricdatatothatofpreviousstudiesonAfricanVaranusspp.
haemogre-garines[9,13–15](seeTable1).
Parasitaemiawascalcu-latedper100erythrocytes,withc.
104erythrocytesexaminedperbloodsmear[16–18].
DNAextraction,PCRamplificationand18SrDNAsequenceanalysisEthanol-preservedbloodsampleswereusedformolecu-larwork.
GenomicDNAofhaemogregarinespecieswasextractedfromthesamplesusingarapidDNAextrac-tionmethodasdetailedintheKAPAExpressExtractKit(KapaBiosystems,CapeTown,SouthAfrica).
Basedonpreviousstudies,amplifyingfragmentsofthe18SrRNAgeneofreptilehaemogregarinesofthegeneraKaryolysusCooketal.
Parasites&Vectors(2016)9:347Page2of13[7],Hemolivia[12]andHepatozoon[19],identificationoftheparasiteofthetwoVaranusspecies,twoV.
aalbigularisandoneV.
niloticus(n=3)fromthecurrentstudywascompletedusingtheprimersetHepF300(5′-GTTTCTGACCTATCAGCTTTCGACG-3′)andHepR900(5′-CAAATCTAAGAATTTCACCTCTGAC-3′).
ThePCRreactionswereruntargetingafragment(approximately600nt)ofthe18SrRNAgene[20].
ConditionsforPCRwereasfollows:initialdenatur-ationat95°Cfor3min,followedby35cycles,entailinga95°Cdenaturationfor30s,annealingat60°Cfor30swithanendextensionat72°Cfor1min,andfollowingthecyclesafinalextensionof72°Cfor10minasdetailedaccordingtopreviousmethods[12,18].
PCRreactionswereperformedwithvolumesof25μl,using12.
5μlThermoScientificDreamTaqPCRmastermix(2*)(2*DreamTaqbuffer,0.
4mMofeachdNTP,and4mMMgCl2),1.
25μlofeachprimer,andatleast25ngDNA.
ThefinalreactionvolumewasmadeupwithPCR-gradenucleasefreewater(ThermoScientific,Vilnius,Lithuania).
ReactionswereundertakeninaBio-RadC1000TouchThermalCyclerPCRmachine(Bio-Rad,HemelHempstead,UK).
Resultingampliconswerevisualizedunderultravioletlightona1%agarosegelstainedwithgelredusingaBio-RadGel-DocXR+imagingsystem(Bio-Rad,HemelHempstead,UK).
TwoPCRproductsfromeachsampleweresenttoacommercialsequencingcompany(InqabaBiotechnicalIn-dustries(Pty)Ltd,Pretoria,SouthAfrica)forpurificationandsequencinginbothdirections.
Resultantsequenceswereassembled,andchromatogram-basedcontigsweregeneratedandtrimmedusingGeneiousVer.
7.
1[21].
Se-quenceswereidentifiedusingtheBasicLocalAlignmentSearchTool(BLAST)[22],anddepositedintheNCBIGenBankdatabaseunderaccessionnumbersKX011039andKX011040.
ComparativesequencesforspeciesofHemolivia,Hepatozoon,Karyolysus,Haemogregarina,DactylosomaLabbé1894andBabesiosomaJakowska&Nigrelli,1956parasitisingreptiles,amphibians,mammalsandticksweredownloadedfromGenBankandalignedtothese-quencesgeneratedinthisstudy.
AdelinadimidiataSchneider,1875,AdelinagrylliButaeva,1996(GenBank:DQ096835–DQ096836)andKlossiahelicinaSchneider,1875(GenBank:HQ224955)werechosenastheout-grouptorootthephylogeny.
SequenceswerealignedusingtheMUSCLEalign-menttool[23]implementedinGeneious7.
1.
TheTable1SpeciesofhaemogregarinesofthegenusHepatozoondescribedfromAfricanvaranidsSpeciesType-hostType-localityOtherhosts(localities)Peripheralgamont/nucleusdimensionsReferenceHepatozoonborreli(Nicolle&Comte,1906)Smith,1996bVaranusgriseus(Daudin,1803)Tunisia7–8*2/1–3*1–2[3]b,[6,14]Hepatozooncamarai(Dias,1954)Smith,1996bVaranusalbigularisalbigularis(Daudin,1802)Mozambique2formsobserved:banana-shaped:11.
75*5.
00;longcurved:14.
25–18.
25*1.
25–4.
25/nucleusirregular[3]b,[9]Hepatozoonparadoxa(Dias,1954)Smith,1996bV.
a.
albigularis1MozambiqueV.
niloticus(Kenya)2;V.
a.
albigularisandV.
niloticus(SouthAfrica)36.
75–7.
50*4.
25–5.
50/nucleusnotvisible1;8.
1*5.
2/nucleusirregularornotvisible2;6.
99*4.
39/nucleusnotvisible3[9]1,[30]2,thisstudy3,[3]bHaemogregarinaroshdyiRamdan,Sauod,Mohammed&Fawzi,1996[probablyHepatozoonroshdyi(Ramdan,Sauod,Mohammed&Fawzi,1996)]V.
griseusEgypt13–20*1.
5–2.
5/6.
0–8.
5*1.
5–2.
5[40]aHepatozoontoddi(Wolbach,1914)Smith,1996bVaranusniloticus(Linnaeus,1762)TheGambia10.
3*2.
5/notgiven[3]b,[6,15]Hepatozoonvarani(Laveran,1905)Smith,1996bV.
niloticus1SouthAfricaV.
niloticusandV.
griseus(Senegal)2;V.
niloticus()(PortugueseGuinea)3;V.
niloticus(TheGambia)4;V.
niloticus(Senegal)5;VaranusarenariusDuméril&Bibron,1836()(FrenchWestAfrica)6;V.
griseus(FrenchSudan)7;V.
niloticus(Kenya)814*3/notgiven1;11–15*3/notgiven2;12.
0*13/3.
5–4.
5*3.
03;10.
3*2.
5/notgivena4;twoforms:12–14*2/notgiven,10–12*4–5/5–6*4–55;reportonly6;reportonly7;12.
7*4.
6/notgiven8[13]1,[41]2,[15]3,[42]3,[43]4,[44]5,[45]6,[46]7,[30]8,[3]baHepatozoon(syn.
Haemogregarina)toddi(Wolbach,1914)whenfirstdiscoveredwastentativelythoughttobeHepatozoon(syn.
Haemogregarina)varani(Laveran,1905)bSmith[3]duringasystematicrevisionofspeciesoftheHepatozoidaetransferredmanyoftheabovespeciesfromthegenusHaemogregarinaDanilewsky,1885tothegenusHepatozoonMiller,19081–8CorrespondingdescriptionorreportofthehaemogregarinespeciesanditsbibliographicreferenceCooketal.
Parasites&Vectors(2016)9:347Page3of13alignmentconsistedof47sequences,manuallytrimmedtoatotallengthof968nt.
Uncorrectedpair-wisedis-tances(p-distance),basepairdifferencesaswellaspar-simonyinformativesitesandthenumberthereofwereidentifiedordeterminedwiththeMEGA6bioinformat-icssoftwareprogram[24]forthealigned18SrDNAse-quencesbetweenallavailablespeciesappearinginthephylogeneticanalyses.
ToinferphylogeneticrelationshipsofthealigneddatasetbothBayesianinference(BI)andMaximumlikelihood(ML)methodswereused.
Acomprehensivemodeltestwaspreformedtodeterminethemostsuit-ablenucleotidesubstitutionmodel,accordingtotheAkaikeinformationcriterionusingjModelTest2.
1.
7[25,26].
ThebestmodelidentifiedwastheTransver-sionModelpluswithestimatesofinvariablesitesandadiscreteGammadistribution(TVM+I+Γ).
ThismodelwassubstitutedwiththeGeneralTimeReversiblemodel(GTR+I+Γ)forphylogeneticanalysis,asthiswasthemostap-propriatemodelavailablewiththebestAICcscore.
TheBIanalysiswasimplementedfromwithinGeneious7.
1usingMrBayes3.
2.
2[27].
Theanalysiswasruntwiceover10milliongenerationsfortheMarkovChainsMonteCarlo(MCMC)algorithm.
TheMarkovchainwassam-pledevery100cycles,andtheMCMCvariantcontained4chainswithatemperatureof0.
2.
Thelog-likelihoodvaluesofthesamplepointwereplottedagainstthegenerationtimeandthefirst25%ofthetreeswerediscardedas'burn-in'withno'burn-in'samplesbeingretained.
TheMLanalysiswasperformedusingRAxMLVer.
8.
1.
22[28]implementedintheraxmlGUIVer.
1.
3[29].
Thealpha-parameterselectedwastheGTR+I+Γmodel,withsupportassessedusing1,000rapidbootstrapinferences.
Resultingtreeswerecombinedina50%majorityconsensustree.
EthicsstatementThisstudyreceivedtherelevantethicalapproval(North-WestUniversityethicsapprovalno:NWU-00005-14-S3).
ResultsPrevalence,parasitaemiaandgeneralobservationsofH.
paradoxainperipheralbloodsmearsSixadultandtwojuvenileVaranusalbigularisalbigu-laris(Fig.
1a,b)andoneadultVaranusniloticuswerecaptured,sampledforbloodparasitesandexaminedforpossiblehaematophagousarthropods.
Thetwojuvenileswerefoundtobenegativeforbloodparasites,whilst4/6(67%)adultV.
a.
albigularisand1/1(100%)adultV.
niloticuswerefoundpositiveforhaemogregarineinfec-tions.
Twoadult(33%)(withaparasitaemiaofc.
5and20%,respectively)V.
a.
albigularisandtheV.
niloticus(withaparasitaemiaofc.
0.
2%)wereparasitisedbyahaemogregarinealiketoHepatozoonparadoxadescribedbyDias[9](Figs.
1c–fand2b–i).
Oneofthetwo(17%)V.
a.
albigularis(parasitaemiaof5%)andthesingleV.
niloticuswerefoundtohaveaco-infectionwithanotherhaemogregarineofadifferentHepatozoonspp.
(Cook,Netherlands&Smit,unpublishedobservations)Fig.
1Karyolysusparadoxa(Dias,1954)invaranidlizardVaranusalbigularisalbigularisDaudin,1802.
a–bVaranusalbigularisalbigularis.
bTicksofthespeciesAmblyommaexornatuminfestingtheareaabovetheeyes,theperipheryofthemouthanddeepintothenostrils(arrows).
c–fPeripheralbloodstagesofK.
paradoxacapturedfromtheneohapantotypeslide(NMBP410).
cPossibleraretrophozoitestage,notethattheyounghosterythrocytecytoplasmandnucleusarestillintactandthattheparasitenucleusisvisibleandgranular.
d–fMaturegamontswithinanerythrocyteinwhichshrinkageofthehostcellisapparentandthenucleusdestroyedresultinginaheavilyvacuolatedappearance.
dMaturegamontinwhichfoldingofthegamontmaybeseenwithinthethickcapsule(arrow).
Scale-bar:10μmCooketal.
Parasites&Vectors(2016)9:347Page4of13(parasitaemiaofc.
0.
4and3%,respectively).
ThelatterunidentifiedHepatozoonspecieswasalsofoundinfectingtheremainingtwoparasitisedV.
a.
albigularis(parasit-aemiaofc.
0.
1and0.
5%,respectively).
AllspecimensofbothspeciesofVaranuswereinfestedwithadultandjuvenilestagesoftheLeguantickAmblyommaexornatum(Koch,1844)(Fig.
1b),thetwojuvenileV.
a.
albigularisandV.
niloticusdemonstratinglowerdensitiesoftheseectoparasites.
Theseectoparasiteswerefoundallovertheanimal,withthehighestdensitiesinthenostrils,oftenblockingthem,alsoontheareasurroundingtheeyes,andalongtheedgesofthemouth.
Itwasnotuncommontofinddeadticksandtheirremainsdeepwithinthenostril,theirabilitytoexitblockedbylaterarrivals.
Squashesofnymphalandadultfemaleandmaleticksprovidednoparasiticstages.
Nootherhaematophagousarthropods,includ-ingmites,wereobserved.
StagesoftheH.
paradoxa-likehaemogregarineobservedinperipheralbloodsmearsfromthisstudywerecomparedmorphologicallywiththoseobservedinpreviousbloodparasitestudiesofAfricanvaranids[9,13–15](Table1).
Insizeandmorphology,theH.
paradoxa-likestagesobservedduringthecurrentstudyconformedtothosede-scribedbyDias[9]andBall[30](seeTable1,Figs.
1a–f,2b–iand3a–d,respectively).
Twostagesoftheparasitewereobserved:astageunreportedbyDias[9],butpossiblybyBall[30],ararepossibletrophozoitestage(Figs.
1cand3a),andwhatwasidentifiedasamatureintra-erythrocyticgamontstage(Figs.
1d–f,2b–iand3d,e).
Karyolysusparadoxa(Dias,1954)Cook,Netherlands&Smit,2016SynsHaemogregarinaparadoxaDias,1954;HepatozoonparadoxaSmith,1996.
Type-host:VaranusalbigularisalbigularisDaudin,1802,Squamata:Varanidae[9].
Otherhosts:Varanusniloticus(LinnaeusinHasselquist,1762),Squamata:Varanidae[30];presentstudy.
Vector:Unknown.
Type-locality:NdumoGameReserve(26°54′18.
5″S,32°19′24.
7″E),KwaZulu-Natal,SouthAfrica(presentstudy).
Otherlocalities:Maputo,Mozambique[9].
Ball[30]alsodescribedahaemogregarinemorphologicallyaliketoK.
paradoxainMarimonti,nearMeru,Kenya.
Fig.
2IllustrationofHaemogregarinaparadoxaDias,1954inVaranusalbigularisalbigularisDaudin,1802[9].
a-iRedrawnandadaptedfromDias(1954).
IllustrationsrepresentingtheoriginaldescriptionofKaryolysusparadoxa(syns.
HepatozoonparadoxaandHaemogregarinaparadoxa)exVaranusalbigularisalbigularisfromMozambique.
aHealthynon-parasitisederythrocyte.
b-iParasitisederythrocytes,notetheshrinkageofthehostcell,theheavyvacuolizationofthehostcellnucleus,andthethickcapsulesurroundingthegamontwhichresultsinthegamontnucleusbeinginvisible.
Scale-bar:10μmCooketal.
Parasites&Vectors(2016)9:347Page5of13Type-material:Neohapantotype,1*bloodsmearfromthetype-hostVaranusalbigularisalbigularisandnewdes-ignatedlocality(26°54′18.
5″S,32°19′24.
7″E),depositedintheprotozoancollectionoftheNationalMuseum,Bloem-fontein,SouthAfricaunderaccessionnumberNMBP410.
OthervouchermaterialdepositedthatincludesstagesofK.
paradoxa,1*bloodsmearfromVaranusniloticus,depositedintheprotozoancollectionoftheNationalMu-seum,Bloemfontein,SouthAfricaunderaccessionnum-berNMBP411.
RepresentativeDNAsequences:Twosequencesrepre-sentinga611and613ntfragmentofthe18SrRNAgeneofK.
paradoxaisolatedfromthetype-hostVaranusalbi-gularisalbigularis,depositedintheNCBIGenBankdatabaseundertheaccessionnumbersKX011039andKX011040,respectively.
Redescription(Figs.
1,2and3)Trophozoite.
Rare,ovoid,withvacuolatedcytoplasm,measuring6.
5–6.
9*4.
3–4.
7(6.
7*4.
5)μm(n=2);nucleuswithloosechromatin,stainingpink(Fig.
1c).
Bothtrophozoitesparasitisingyoungerythrocytes,nohostcelldistortionvisible.
Maturegamont.
Roundedinshape,gamontseeminglyfoldedwithinwithawell-developedcapsule(Fig.
1d–f),measuring6.
3–7.
9*3.
6–5.
2(7.
0*4.
4)μm(n=20).
Cytoplasmstainingwhitish-blue;nucleusnotvisible.
Notabledestructionofhostcellcytoplasmandkaryolysisofthehostcellnucleus,causinganobservableheavilyvacuolatedandfoamyappearance(Fig.
1d–f).
RemarksThehaemogregarinedescribedinthisstudyfromSouthAfricanVaranusalbigularisalbigularisandVaranusniloticus(Fig.
1d–f)wasfoundtobemorphologicallysimilartoHepatozoonparadoxadescribedbyDias[9]fromaspecimenofV.
a.
albigularisinneighbouringMozambique(Fig.
2b–i).
Itsharedanumberofuniquecharacteristicsincludingdestructionoftheinfectedhosterythrocyte,consistingofdehaemoglobinisationresultinginshrinkageofthehostcellanddestructionofthehostcellnucleus(characteristicofanumberofspeciesofKaryolysus[1])resultinginaheavilyvacuolatedappear-ance(Figs.
1d–fand2b–i).
Additionally,thehaemogre-garineinthisstudyagreeswellwiththesizeofH.
paradoxaintheoriginaldescriptionofDias[9](mean7.
0*4.
4vs7.
0*4.
9μm)(Table1).
ThesameuniquecharacteristicswerereportedofahaemogregarinefoundinfectingaV.
niloticusbyBall[30]fromKenya,measuringonaverage8.
1*5.
2μm(Fig.
3c,d).
However,inBall's[30]study,additional,pre-sumablyyounger,stageswereobserved(similartotheyoungtrophozoitestagefoundinourstudy)(Figs.
1cand3a).
Ball[30]alsonotedasinglepossiblydividingstageofthesetrophozoites(Fig.
3b).
Incellsparasitisedbyallthesepossibletrophozoitestages,thehosterythro-cyteshowednoshrinkageasofyet,butwasaccordingtoBall'sreportabnormalinshapeandstaining.
Atfirst,Ball[30]didassumethisparasitetorepresentyoungerstagesofanotherhaemogregarinethathasbeenreportedparasitisingAfricanvaranidsHepatozoonvarani(Lave-ran,1905)Smith,1996(Table1).
However,basedontheeffectsoftheparasiteresemblingH.
paradoxaasde-scribedbyDias[9],itsdestructionofthehostcellandthehostcell'snucleus,heconcludedthatthisparasitewasnotH.
varani.
Overall,fortheK.
paradoxade-scribedinthisstudyandtheparasitesdescribedintheothertwostudies[9,30],thenucleusandcytoplasmwasnotvisibleowingtowhatappearedtobeathicken-closingcapsuleasseenwiththegamontsofspeciesofHemolivia,see[12,31](Figs.
1d–f,2b–iand3c,d).
OnlyonrareoccasioninthepresentstudyandthatofBall's[30]wastheparasiteseentobefoldingFig.
3IllustrationofanunknownhaemogregarinebyBall(1967)[30]inVaranusniloticus(LinnaeusinHasselquist,1762).
a–dRedrawnandadaptedfromBall(1967)(hisFigs.
19–22).
IllustrationsrepresentinganunknownhaemogregarinefoundasaconcurrentinfectiontoHepatozoonvarani(Laveran,1905)(syn.
Haemogregarinavarani)inaVaranusniloticusfromKenya.
aPossibletrophozoitestageoryoungstage,shrinkageandvacuolationofthehostcellandhostcellnucleusnotyetapparent.
bPossibledividingstage.
c–dMaturegamontstages,notetheshrinkageandheavyvacuolationofthehostcellerythrocyte.
cMaturegamontfoldedwithinacapsulewithpossiblenucleus.
dFoldingofthegamontandthegamontnucleusnotobservablethroughthecapsule.
Scale-bar:10μm(eventhoughnotprovidedbyBall,wehaveprovidedascalematchingtotheintextdescription)Cooketal.
Parasites&Vectors(2016)9:347Page6of13overonitself(Figs.
1dand3c).
Otherwise,theonlyevidenceofthisbehaviourwasacrescentshapedstainatthecentreoftheovalparasite(asseeninallthreereports)(Figs.
1,2and3).
Itisbasedontheaboveuniquecharacteristics,particularlyofthema-turegamontstages,thatwesuggestallthreereportsareofthesameparasitespeciesK.
paradoxa.
Nohapantotype,accordingtotheInternationalCodeofZoologicalNomenclature(ICZN)Article73.
3,wasdesignatedanddepositedduringtheoriginaldescriptionofK.
paradoxabyDias[9].
Ball[30]alsodidnotidentifytheparasitetotaxonlevelanddidnotdepositanyvouchermaterial.
Furthermore,oureffortstolocateanyoriginalspecimensorvouchermaterialwereunsuccessful.
Inthisstudy,K.
paradoxawascollectedfromNdumoGameReserve,northernKwaZulu-Natal,SouthAfrica,borderingthesouthofMozambique.
TheoriginaldescriptionbyDias[9]wascollectedinthevicinityofMaputointhesouthernpartsofMozambiqueapproximately300kmfromtheNGR.
Additionally,K.
paradoxainthepresentstudywascol-lectedfromthesamehostspeciesVaranusalbigularisalbigularisasintheoriginaldescriptionbyDias[9].
Basedontheabove,thematuregamontsizecomparisons,andtheuniquecharacteristicsofthematuregamontsofK.
paradoxa(destructionandshrinkageoftheinfectedhosterythrocyte,destructionofthehostcellnucleusresultinginvacuolation,andthethicknon-stainingcapsule)asdescribedaboveforallthreereportsofthisparasite,whichincludestheoriginaldescriptionbyDias[9],andinaccordancewithICZNArticle75.
3,wedesignateaneohapantotype.
Thepresentstudyalsoincludesboththedescriptionofanadditionalstageoftheparasite(atrophozoite)andprovidessequencedata(fragment18SrDNA),whichwasnotprovidedbyDias[9]inhisori-ginaldescriptionofthisparasitespecies.
Thisneoha-pantotypeisdepositedintheprotozoancollectionoftheNationalMuseum,Bloemfontein,SouthAfricaunderaccessionnumberNMBP410.
SequenceidentificationandphylogeneticanalysisAmpliconsof611and613ntforthe18SrRNAgeneofK.
paradoxawereobtainedfromtheV.
a.
albigulariswiththeseeminglypureandhighestparasitaemiainfec-tionusingprimersetsHepF300andHepR900.
NoK.
paradoxaisolatewasobtainedfromtheV.
niloticus,likelyduetoitslowparasitaemiaincomparisontotheconcurrentinfectionoftheotherunidentifiedHepato-zoonspecies(unpublisheddata).
Thedetailsofthespe-ciesusedinthephylogeneticanalysesandpresentedintheconsensustreeareprovidedinTable2.
ThetopologyofboththeBIandMLanalyseswereoverallsimilar,withdiscretemonophyleticcladesofknownandlikelybelong-ingtoKaryolysusspecies,Hepatozoonspp.
ofmammals('intraleucocytic'Hepatozoonspp.
),theherpatofauna('intraerythrocytic'Hepatozoonspp.
),Hemoliviaspp.
,Haemogregarinaspp.
,andtheDactylosomatidae(Fig.
4).
OurresultsshowedthatK.
paradoxaclusteredwithinamajormonophyleticcladecontainingbothknown(mor-phologicallyandecologicallyconfirmed)andlikelybe-longingtoKaryolysusspecies,and'intraleucocytic'HepatozoonspeciesofmammalsthatweresistertotheKaryolysusclade.
Thismajorcladewasdistinctfromthemajormonophyleticcladecontainingtheherpatofaunal'intraerythrocytic'Hepatozoonspp.
andHemoliviaspp.
Theparasitecollectedinthepresentstudythatmor-phologicallyconformedtotheoriginaldescriptionofHepatozoonparadoxaformedasistertaxontothelarger'Karyolysus'clade(containingtheknownandlikelybe-longingtoKaryolysusspecies)inthephylogeneticana-lysis(Fig.
4).
AccordingtoevolutionarydivergenceestimatesthepresentmaterialwasmostcloselyrelatedtoknownKaryolysusspp.
(at97.
7%,p-distance=0.
02)thantospeciesofthegeneraHepatozoon,HemoliviaandHaemogregarina(Table3).
DiscussionVaranusalbigularisalbigularisandVaranusniloticusareknowntodisplaysomewhatdifferenthabitatprefer-ences,preferringmoreterrestrialandaquaticenviron-mentsrespectively[32].
BothV.
a.
albigularisandV.
niloticuscanbefoundthroughoutSouthAfricafromthemoretropicalIndianOceancoastalbeltintheEast,westtothemarginsofthemorearidNorthernandWesternCapeprovinces[32].
Bothspeciesoftenoccursympatri-cally,particularlyinNdumoGameReserve,renderingthefindingofthesameparasiteinboththesespeciesnotsurprising.
AsthemorphologicalanddevelopmentalcharacteristicstypicalofHaemogregarinaspp.
hadneverbeenobservedinhaemogregarinesoftheherpatofauna,K.
paradoxawastransferredalongwithmanyotherspeciesfromtheherpa-tofauna,birdsandmammals,fromthegenusHaemogre-garina(Haemogregarinidae)tothegenusHepatozoon(Hepatozoidae)bySmith[3]duringasystematicreviewoftheHepatozoidae.
However,duringthefirstdetailedrevi-sionofK.
paradoxaprovidedhereinthepresentstudy,morphologicallythisspeciessharesmorecharacteristicswithmembersofthefamilyKaryolysidae.
Karyolysusparadoxaperipheralbloodgamontsappeartobeencapsu-latedasintheHemoliviaanddestroythehostcellnucleusasinanumberofspeciesofKaryolysus(asmentionedaboveinRemarks)[1,7].
However,itisimperativetotakeintoaccountthatthesemorphologicalfeatures,specificallyforthelattergenus,arenotalwayspresent[7].
ThephylogeneticpositionofK.
paradoxawasshowntobeatthebaseofacladecontainingunde-scribedspeciesofHepatozoon,manyofthesetheCooketal.
Parasites&Vectors(2016)9:347Page7of13Table2Listoforganismsusedinthephylogeneticanalysesofthisstudy,withassociatedhost,hostfamilyandhostcommonname,GenBankaccessionnumbersandreferencesCladeOrganismHostFamilyCommonnameAccessionnumberReferenceKaryolysusKaryolysuslatusPodarcismuralisLacertidaeCommonwalllizardKJ461939[7]Hepatozoonsp.
AlgyroidesmarchiLacertidaeSpanishkeeledlizardJX531933[34]Hepatozoonsp.
PodarcisvaucheriLacertidaeAndalusianwalllizardHQ734803[33]KaryolysuslacazeiLacertatrilineataLacertidaeBalkanemeraldlizardKJ461942[7]Karyolysussp.
ZootocaviviparaLacertidaeViviparouslizardKJ461945[7]Hepatozoonsp.
PodarcisbocageiLacertidaeBocage'swalllizardJX531952[34]Hepatozoonsp.
PodarcisbocageiLacertidaeBocage'swalllizardJX531954[34]Hepatozoonsp.
TimontangitanusLacertidaeMoroccaneyedlizardHQ734799[33]Hepatozoonsp.
AtlantolacertaandreanskyiLacertidaeAndreansky'slizardHQ734798[33]Karyolysussp.
Ophionyssussp.
exLacertaviridisLacertidaeEuropeangreenlizardKJ461944[7]Hepatozoonsp.
PsammophisschokariLamprophiidaeSchokarisandracerKC696565[47]Hepatozoonsp.
HemorrhoishippocrepisColubridaeHorseshoesnakeJX244269[35]Hepatozoonsp.
EumecesalgeriensisScincidaeAlgerianskinkHQ734796[33]Hepatozoonsp.
ScelarcisperspicillataLacertidaeMenorcawalllizardHQ734791[33]Hepatozoonsp.
PodarcisvaucheriLacertidaeAndalusianwalllizardHQ734804[33]Hepatozoonsp.
PodarcishispanicusLacertidaeIberianwalllizardJX531917[34]Hepatozoonsp.
PodarcislilfordiLacertidaeLilford'swalllizardJX531920[34]Karyolysus(syn.
Hepatozoon)paradoxaVaranusalbigularisVaranidaeRockmonitorlizardKX011039ThisstudyKaryolysus(syn.
Hepatozoon)paradoxaVaranusalbigularisVaranidaeRockmonitorlizardKX011040ThisstudyIntraleucocyticHepatozoonHepatozoonamericanumCanisfamiliarisCanidaeDomesticdogAF176836[48]HepatozooncanisCanisfamiliarisCanidaeDomesticdogAY461378[49]HepatozoonursiUrsusthibetanusjaponicusUrsidaeJapaneseblackbearEU041718[50]Hepatozoonsp.
MartesmartesMustelidaePinemartenEF222257[51]HepatozoonfelisFeliscatusFelidaeDomesticcatAY620232[50]HepatozoonHepatozooncatesbianaeLithobatescatesbeianusRanidaeAmericanbullfrogHQ224954[49]HepatozoonixoxoSclerophrys(syn.
Amietophrynus)maculatusBufonidaeFlat-backedtoadKP119772[52]HepatozoontheileriAmietiaqueckettiPyxicephalidaeQueckett'sriverfrogKP119773[18]HepatozoonsipedonNerodiasipedonColubridaeNorthernwatersnakeJN181157[49]HepatozoonfitzsimonsiKinixyszombensisTestudinidaeBell'shingebacktortoiseKR069084[12]Cooketal.
Parasites&Vectors(2016)9:347Page8of13Table2Listoforganismsusedinthephylogeneticanalysesofthisstudy,withassociatedhost,hostfamilyandhostcommonname,GenBankaccessionnumbersandreferences(Continued)CladeOrganismHostFamilyCommonnameAccessionnumberReferenceHepatozoonayorgborPythonregius(aLamprophis(syn.
Boaedon)fuliginosus)Pythonidae;aLamprophiidaeBallpython;brownhousesnakeEF157822[53]HepatozoondomergueiFurcifersp.
ChamaeleonidaeChameleonspeciesKM234649[54]HepatozoonseychellensisGrandisoniaalternansIndotyphlidaeCaecilianKF246566[55]HemoliviaHemoliviamauritanicaTestudograecaTestudinidaeMediterraneanspur-thighedtortoiseKF992700[36]HemoliviaparvulaKinixyszombensisTestudinidaeBell'shingebacktortoiseKR069082[12]HemoliviamariaeEgerniastokesiiScincidaeGidgeespiny-tailedskinkKF992711[36]Hemoliviasp.
RhinoclemmyspulcherrimamanniGeoemydidaePaintedwoodturtleKF992714[36]HemoliviastellataAmblyommarotundatumexRhinellamarinaBufonidaeCanetoadKP881349[37]HaemogregarinidaeHaemogregarinasp.
_3140PelusiossubnigerPelomedusidaeEastAfricanblackmudturtleKF257925[56]HaemogregarinasacaliaeSacaliaquadriocellataGeoemydidaeFour-eyedturtleKM887507[57]HaemogregarinapellegriniMalayemyssubtrijugaGeoemydidaeSnail-eatingturtleKM887508[57]HaemogregarinaballiChelydraserpentinaserpentinaChelydridaeCommonsnappingturtleHQ224959[49]HaemogregarinastepanowiMauremyscaspicaGeoemydidaeCaspianturtleKF257926[56]DactylosomatidaeBabesiosomastableriLithobatesseptentrionalisRanidaeMinkfrogHQ224961[49]DactylosomaranarumPelophylaxlessonae(syn.
esculentus)RanidaePoolfrogHQ224957[49]OutgroupKlossiahelicinaCepaeanemoralisHelicidaeGrovesnailHQ224955[49]AdelinadimidiataScolopendracingulataScolopendridaeMegarianbandedcentipedeDQ096835[58]AdelinagrylliGryllusbimaculatusGryllidaeFieldcricketDQ096836[58]aExperimentallaboratoryanimalCooketal.
Parasites&Vectors(2016)9:347Page9of13resultsofmolecularHepatozoonspp.
surveys[33,34],andknownspeciesofKaryolysus[7].
Thismaysug-gestthattheseundescribedHepatozoonspp.
mightra-therbespeciesofKaryolysus.
TheKaryolysusspp.
cladeispartofalargercladeincludingasistercladeofHepatozoonspp.
frommammals.
Thistopology,asseeninthisstudy,hasbeenobservedinanumberofotherstudies[7,33–37].
Karadjianetal.
[37],intheirattempttounderstandtherelationshipsofthediffer-enthaemogregarinegenera,particularlyinrespecttotheconundrumofthepolyphyleticHepatozoonclade,proposed,basedontheirphylogeneticfindings,thatanumberofHepatozoonspecies(mostofwhichhavenotbeenmorphologicallydescribed)mightratherrep-resentspeciesofKaryolysus.
Giventhatitwasonlysincetherecentidentification,descriptionandmolecularchar-acterisationofKaryolysusspp.
by[7],itisonlynowthatwearebeginningtorealizethispossibility.
Moreover,itappearsthatthediversityofsquamatesparasitisedbylikelyspeciesofKaryolysusisfastincreasing,ascenarioFig.
4PhylogeneticanalysisofKaryolysusparadoxa(Dias,1954)basedon18SrDNAsequences.
Bayesianinference(BI)andMaximumLikelihood(ML)analysisshowingthephylogeneticrelationshipsfortwoKaryolyusparadoxaisolatesfromthecurrentstudyrepresentedinbold,17KaryolysusandHepatozoonspecies(representingtheKaryolysuscladeinblue),fivemammal-infectingHepatozoonspecies(representingthe'intraleucocytic'Hepatozooncladeinpurple),eightherpatofaunal-infectingHepatozoonspecies(representingthe'intraerythrocytic'Hepatozooncladeingreen),fiveHemoliviaspecies(representingtheHemoliviacladeinorange),fiveHaemogregarinaspecies(representingtheHaemogregarinidaecladeinyellow),aBabesiosomaandDactylosomaspecies(representingtheDactylosomatidaecladeinpink)andtwoAdelinaandoneKlossiaspecies(usedastheoutgroupingrey).
AllcomparativesequencesweredownloadedfromtheGenBankdatabase.
TreetopologiesforboththeBIandMLtreeswereidentical;thenodalsupportvalues(BI/ML)arerepresentedontheBItreeCooketal.
Parasites&Vectors(2016)9:347Page10of13whichislikelyonlytointensifyinfuture.
Arecentmo-lecularHepatozoonspp.
surveyby[38]showshaemogre-garineisolatesfromgeckosofthegenusTarentolaGray,1825(Phyllodactylidae)arealsofallingwithinwhatmaybeseenasthe'Karyolysus'clade;acladethatatpresentalsoincludeshaemogregarinesisolatedfromspeciesofthefamiliesColubridaeandLamprophiidae(bothsnakes),LacertidaeandVaranidae(bothlizards),andScincidae(skinks).
Furthermore,thepresentstudyshowsthatK.
paradoxaismostcloselyrelatedtoknownKaryolysusspecies,followedbyspeciesofthe'intraleucocytic'Hepatozooncladeandthenspeciesofthe'intraerythrocytic'Hepato-zoonandHemoliviaclades.
KaryolysusandHemoliviamorphologicallystillbelongwithinthesamefamily(Karyolisidae),however,withtheuseofthe18SrRNAgenethesetwogenerainthisstudyandinothersgenerallyfallindifferentmajorclades.
Karyolysusclustersinamajorcladewiththe'intraleucocytic'mammalHepatozoon,whilstHemoliviaclustersinamajorcladewiththe'intraerythrocytic'herpatofaunalHepatozoon;thisfindingisapparentinthepresentstudy,andalsoin[36,37].
Itisclearthattherelationshipbetweenthesetwogeneramaypossiblyonlyberesolvedbyusingamulti-geneapproachasin[39].
ConclusionsBasedonthemorphologyandthemolecularfindingspresentedinthisstudy,werecommendthefollowingnomenclaturalcorrection:Karyolysusparadoxa(Dias,1954)(syn.
Hepatozoonparadoxa(Dias,1954)Smith,1996,HaemogregarinaparadoxaDias,1954)inthevara-nidlizardsVaranusalbigularisalbigularis(type-host),andVaranusniloticus.
OurresultsshowedthatKaryoly-susparadoxaisascloselyrelatedtospecieswithinitscurrentgenericassignmentinthe'intraerythrocytic'her-patofaunalHepatozoonasitiswiththemoredistantlyrelatedspeciesoftheHaemogregarina.
Besidesthisstudyrepresentingthefirstmorpho-logicalandmolecularreportofahaemogregarinewithinanAfricanvaranid,itisthefirstreportofaspe-ciesofKaryolysusinfectingahostoftheVaranidae.
Furthermore,itrepresentsthethirddescribedandnamedKaryolysusspp.
forwhichthereisanucleotidesequenceavailable.
Itishopedthatthisstudywillen-couragefurthermolecularworkontheKaryolysidae,particularlythegenusKaryolysus.
ThisstudyalsoextendsthehostanddistributionrangeofK.
paradoxafromonlyasinglespecimenofV.
a.
albigularisinMozambiquetoanadditionaltwospecimensinSouthAfrica,aswellasincludingV.
niloticusasanadditionalhostbothinSouthAfricaandKenya.
ThedistributionrangeofK.
paradoxafallswithinsubtropicalareasinSouthAfrica,MozambiqueandKenya,andassuchitwouldbein-terestingtoseeifthisparticularparasiteisrestrictedtosubtropicalareassuchasisthecasewithHemoliviaparvula(Dias,1953)foundparasitisingKinixyszombensisHewitt,1931tortoisesofSouthAfricaandMozambique;see[12],orifitismorewidelydistributedthroughoutdif-ferentbiomesasisthecasewithHepatozoonfitzsimonsi(Dias,1953)foundparasitisingseveraltortoisespeciesinSouthAfricaandMozambique;see[19].
Eventhoughticksquashesdidnotresultinanyob-servableparasiticstagesfuturestudieswillfocusonidentifyingpossiblevectors.
Parasiticstagesfoundinthesepossiblevectorswillbeidentifiedtospecieslevelbasedonbothmorphologicalandmolecularfindings.
WiththeconundrumofthelargerHepatozooncladebeingpolyphyleticandabsorbingtheHemoliviaandKaryolysus,itisimportanttoincreasethenumberoftaxafromwhichwecanworkandaskdeeperphylogen-eticquestions.
However,besidesthemolecularcharac-terisationofthesespecies,itisstillimportanttofocusontheirmorphologyandwherepossibleattempttoelu-cidatetheirlife-cyclesinordertoresolvethecomplextaxonomyoftheseorganisms.
Moreimportantly,itisnecessarytoincludeanotherfasterevolvinggeneorevenmitochondrialgenomesofthesegroupsfollowing[39]beforewecanmakeanywell-informeddecisions.
AcknowledgementsWewouldliketothankNdumoGameReserveandEzemveloKZNWildlife,foraccesstositesandresearchpermits(OP839/2014;OP1262/2015;OP2492/2015)forsamplecollection.
Inaddition,wewouldliketothankTable3RepresentationofevolutionarydivergenceofthedifferentcladesinrelationtoKaryolysusparadoxa(Dias,1954)CladeSimilarityp-distanceNucleotidedifferenceParsimonyinformativesites(%)KnownKaryolysus(4)97.
70.
0214(11–18)73'Intraleucocytic'Hepatozoon(5)95.
70.
04(0.
03–0.
05)26(18–45)29Hepatozoon(8)94.
40.
05(0.
03–0.
09)33(21–56)46Hemolivia(5)97.
00.
0318(17–20)66Haemogregarina(5)94.
70.
0532(29–34)68SummativerepresentationofcladesshowninFig.
4,showingcladeswithnumberofsequencesinparentheses,theaveragepercentsimilarity,averageuncorrectedp-distancewithrangeinparentheses,averagebasepairdifferencewithrangeinparentheses,andpercentparsimonyinformativesites,allcomparedtothetwosequencesasshowninFig.
4ofKaryolysus(syn.
Hepatozoon)paradoxafromVaranusalbigularisalbigularis.
NotethatonlytheknownKaryolysusspecieshavebeenusedfromtheKaryolysuscladeCooketal.
Parasites&Vectors(2016)9:347Page11of13Microbiology,UnitforEnvironmentalSciencesandManagement,(NWU-PC),fortheuseoftheirfacilities.
ThefinancialassistanceoftheNationalResearchFoundation(NRF)ofSouthAfricatoCAC(NRFScarceSkillsPostdoctoralScholarship-GrantSFP13090332476),andtheFlemishInteruniversityCouncil(VLIR)toECN(VLIR-OUSproject–ZEIN21013PR396)isacknowledged.
Opinionsexpressedandconclusionsarrivedat,arethoseoftheauthorsandarenotnecessarilytobeattributedtotheNRForVLIR.
Authors'contributionsAllauthorsconceivedanddesignedtheproject,participatedingeneraldataanalysisandindraftingthemanuscript.
CACandECNcarriedoutthefieldwork,preparedandexaminedbloodsmears,preparedlightmicrographsandcompiledallmeasurementdata.
ECNparticipatedinthemolecularstudiesandinthesequencealignmentandprovidedsupporttothepreparationofthemanuscript.
Allauthorsreadandapprovedthefinalmanuscript.
CompetinginterestsTheauthorsdeclarethattheyhavenocompetinginterests.
Authordetails1UnitforEnvironmentalSciencesandManagement,North-WestUniversity,Potchefstroom,SouthAfrica.
2LaboratoryofAquaticEcology,EvolutionandConservation,UniversityofLeuven,3000Leuven,Belgium.
Received:10March2016Accepted:18May2016References1.
TelfordSR.
HemoparasitesoftheReptilia:ColorAtlasandText.
NewYork:CRCPress;2009.
2.
SmallridgeCJ,PapernaI.
Thetick-transmittedhaemogregarinidoftheAustraliansleepylizardTiliquarugosabelongstothegenusHemolivia.
Parasite.
1997;4:359–63.
3.
SmithTG.
ThegenusHepatozoon(Apicomplexa:Adeleina).
JParasitol.
1996;82:565–85.
4.
MerinoS,MartínezJ,MaselloJF,BedollaY,GuillfeldtP.
FirstmolecularcharacterizationofaHepatozoonspecies(Apicomplexa:Hepatozoidae)infectingbirdsanddescriptionofanewspeciesinfectingstormpetrels(Aves:Hydrobatidae).
JParasitol.
2014;100:338–43.
5.
DaviesAJ,JohnstonMRL.
Thebiologyofsomeintraerythrocyticparasitesoffishes,amphibiansandreptiles.
AdvParasitol.
2000;45:1–107.
6.
VanAsJ,DaviesAJ,SmitNJ.
Hepatozoonlangiin.
sp.
andHepatozoonvacuolatusn.
sp.
(Apicomplexa:Adeleorina:Hepatozoidae)fromthecraglizard(Sauria:Cordylidae)PseudocordyluslangifromtheNorthEasternDrakensbergescarpment,EasternFreeState,SouthAfrica.
Zootaxa.
2013;3608:345–56.
7.
Haklová-KoíkováB,HiňanováA,MajláthI,RakaK,HarrisDJ,FldváriG,TryjanowskiP,KokoováN,MalekováB,MajláthováV.
MorphologicalandmolecularcharacterizationofKaryolysus-aneglectedbutcommonparasiteinfectingsomeEuropeanlizards.
ParasitVectors.
2014;7:555.
8.
VanAsJ,DaviesAJ,SmitNJ.
LifecycleofHepatozoonaffluomalotisp.
n.
(Apicomplexa:Haemogregarinidae)incraglizards(Sauria:Cordylidae)andinculicinemosquitoesfromSouthAfrica.
FoliaParasitol.
2015;62:008.
9.
DiasJATS.
SubsídiosparaoestudodoshematozoáriosdosrépteisdeMoambique3:DuasnovasespéciesdeHaemogregarinaparasitasdoVaranusalbigularisalbigularis(Daudin,1802).
BolSocEstMo.
1954;87:71–9.
10.
NetherlandsEC,CookCA,KrugerDJ,duPreezLH,SmitNJ.
Biodiversityoffroghaemoparasitesfromsub-tropicalnorthernKwazulu-Natal,SouthAfrica.
IntJParasitolParasitesWildl.
2015;4:135–41.
11.
CookCA,SmitNJ,DaviesAJ.
Firstrecordofanintraleucocytichaemogregarine(Adeleorina:Haemogregarinidae)fromSouthAfricantortoisesofthespeciesStigmochelyspardalis(Cryptodira:Testudinidae).
AfrZool.
2014;49:290–4.
12.
CookCA,NetherlandsEC,SmitNJ.
FirstHemoliviafromsouthernAfrica:reassigningchelonianHaemogregarinaparvulaDias,1953(Adeleorina:Haemogregarinidae)toHemolivia(Adeleorina:Karyolysidae).
AfrZool.
2015;50:165–73.
13.
LaveranA.
SurunehaemogregarinedeVaranusniloticus.
CRSeancesSocBiol.
1905;59:175–6.
14.
NicolleMMC,ComteC.
SurunehémogrégarinedeVaranusgriseus.
CRSeancesSocBiol.
1906;61:310–2.
15.
WolbachSB.
Notesonthelifecycleofahemogregarinefoundinamonitor(Veranusniloticus).
JMedRes.
1914;29:473–88.
16.
CookCA,SmitNJ,DaviesAJ.
AredescriptionofHaemogregarinafitzsimonsiDias,1953andsomecommentsonHaemogregarinaparvulaDias,1953(Adeleorina:Haemogregarinidae)fromsouthernAfricantortoises(Cryptodira:Testudinidae),withnewhostdataanddistributionrecords.
FoliaParasitol.
2009;56:173–9.
17.
CookCA,SmitNJ,DaviesAJ.
Hemproteids(Apicomplexa:Haemoproteidae)fromSouthAfricanTortoises(Cryptodiria:Testudinidae).
JParasitol.
2010;96:1168–72.
18.
NetherlandsEC,CookCA,SmitNJ,duPreezLH.
RedescriptionandmoleculardiagnosisofHepatozoontheileri(Laveran,1905)(Apicomplexa:Adeleorina:Hepatozoidae),infectingAmietiaquecketti(Anura:Pyxicephalidae).
FoliaParasitol.
2014;61:293–300.
19.
CookCA,LawtonSP,DaviesAJ,SmitNJ.
ReassignmentofthelandtortoisehaemogregarineHaemogregarinafitzsimonsiDias1953,(Adeleorina:Haemogregarinidae)tothegenusHepatozoonMiller1908(Adeleorina:Hepatozoidae)basedonparasitemorphology,lifecycleandphylogeneticanalysisof18SrDNAsequencefragments.
Parasitology.
2014;141:1611–20.
20.
UjvariB,MadsenT,OlssonM.
HighprevalenceofHepatozoonspp.
(Apicomplexa:Hepatozoidae)infectioninwaterpythons(Liasisfuscus)fromtropicalAustralia.
JParasitol.
2004;90:670–2.
21.
GeneioussoftwarecreatedbyBiomatters.
[http://www.
geneious.
com].
22.
BasicLocalAlignmentSearchTool.
[http://blast.
ncbi.
nlm.
nih.
gov/Blast.
cgi].
23.
EdgarRC.
Muscle:multiplesequencealignmentwithhighaccuracyandhighthroughput.
NucleicAcidsRes.
2004;5:1792–7.
24.
TamuraK,StecherG,PetersonD,FilipskiA,KumarS.
MEGA6:molecularevolutionarygeneticsanalysisVersion6.
0.
MolBiolEvol.
2013;30:2725–9.
25.
GuindonS,GascuelO.
Asimple,fastandaccuratemethodtoestimatelargephylogeniesbymaximum-likelihood.
SystBiol.
2003;52:696–704.
26.
DarribaD,TaboadaGL,DoalloR,PosadaD.
jModelTest2:moremodels,newheuristicsandparallelcomputing.
NatMethods.
2012;9:772.
27.
HuelsenbeckJP,RonquistF.
MrBayes:Bayesianinferenceofphylogenetictrees.
Bioinformatics.
2001;17:754–5.
28.
StamatakisA.
RAxMLversion8:atoolforphylogeneticanalysisandpost-analysisoflargephylogenies.
Bioinformatics.
2014;30:1312–3.
29.
SilvestroD,MichalakI.
raxmlGUI:agraphicalfront-endforRAxML.
OrgDiversEvol.
2012;12:335–7.
30.
BallGH.
SomebloodsporozoafromEastAfricanreptiles.
JProtozool.
1967;14:198–210.
31.
irokP,KamlerM,FryeFL,FictumP,ModrD.
EndogenousdevelopmentofHemoliviamauritanica(Apicomplexa:Adeleina:Haemogregarinidae)inthemarginatedtortoiseTestudomarginata(Reptilia:Testudinidae):evidencefromexperimentalinfection.
FoliaParasitol.
2007;54:13–8.
32.
BatesMF,BranchWR,BauerAM,BurgerM,MaraisJ,AlexanderGJ,deVilliersMS.
AtlasandredlistofthereptilesofSouthAfrica,LesothoandSwaziland.
Suricata1.
Pretoria:SouthAfricanNationalBiodiversityInstitute;2014.
33.
MaiaJPMC,HarrisDJ,PereraA.
MolecularsurveyofHepatozoonspeciesinlizardsfromNorthAfrica.
JParasitol.
2011;97:513–7.
34.
MaiaJPMC,PereraA,HarrisDJ.
MolecularsurveyandmicroscopicexaminationofHepatozoonMiller,1908(Apicomplexa:Adeleorina)inlacertidlizardsfromthewesternMediterranean.
FoliaParasitol.
2012;59:241–8.
35.
ToméB,MaiaJPMC,HarrisDJ.
Hepatozooninfectionprevalenceinfoursnakegenera:influenceofdiet,preyparasitaemialevelsorparasitetypeJParasitol.
2012;98:913–7.
36.
KvierováJ,HypaV,DvoákováN,MikulíekP,JandzikD,GeorgeGardnerM,JavanbakhtH,TiarG,irokP.
HemoliviaandHepatozoon:haemogregarineswithtangledevolutionaryrelationships.
Protist.
2014;165:688–700.
37.
KaradjianG,ChavatteJ,LandauI.
Systematicrevisionoftheadeleidhaemogregarines,withcreationofBartazoonn.
g.
,reassignmentofHepatozoonargantisGarnham,1954toHemolivia,andmoleculardataonHemoliviastellata.
Parasite.
2015;22:31.
38.
ToméB,RatoC,PereraA,HarrisDJ.
HighdiversityofHepatozoonspp.
ingeckosofthegenusTarentola.
JParasitol.
2016.
doi:10.
1645/15-908.
39.
OgedengbeJD,OgedengbeME,HafeezMA,BartaJR.
Molecularphylogeneticsofeimeriidcoccidian(Eimeriidae,Eimeriorina,Apicomplexa,Alveolata):Apreliminarymulti-geneandmulti-genomeapproach.
ParasitolRes.
2015;114:4149–41460.
40.
RamdanNF,SaoudMFA,MohammedSH,FawziSM.
OnanewhaemogregarineofVaranusgriseusfromEgypt.
QatarUnivSciJ.
1996;16:119–25.
Cooketal.
Parasites&Vectors(2016)9:347Page12of1341.
LaveranA,PettitA.
Contributionàl'étudedeshaemogregarinesdequelquessauriensd'Afrique.
BullSocPatholExot.
1909;2:506–14.
42.
FranaC.
NotessurdeshématozoairesdelaGuinéePortugaise.
ArqivRealInstitutdeBactériologieCmaraPestana.
1911;3:229–38.
43.
ToddJL,WolbachSB.
ParasiticprotozoafromTheGambia1912:secondreportoftheexpeditionoftheLiverpoolSchoolofTropicalMedicinetoTheGambia,1911.
JMedRes.
1912;26:195–218.
44.
LegerM,LegerA.
HématozoairesdesreptilesduHaut-Senegal-Niger.
BullSocPatholExot.
1914;7:488–92.
45.
RousselotR.
1943CitedbyRousselot,R.
1953.
46.
RousselotR.
NotesdeParasitologieTropicale.
I.
ParasitesdeSangdesAnimaux.
Paris:VigotFreres;1953.
47.
ToméB,MaiaJPMC,HarrisDJ.
MolecularassessmentofapicomplexanparasitesinthesnakePsammophisfromNorthAfrica:domultipleparasitelineagesreflectthefinalvertebratehostdietJParasitol.
2013;99:883–7.
48.
MathewJS,vandenBusscheRA,EwingSA,MalayerJR,LathaBR,PancieraRJ.
PhylogeneticrelationshipsofHepatozoon(Apicomplexa:Adeleorina)basedonmolecular,morphologic,andlife-cyclecharacters.
JParasitol.
2000;86:366–72.
49.
BartaJR,OgendengbeJD,MartinDS,SmithTG.
Phylogeneticpositionoftheadeleorinidcoccidia(Myzozoa,Apicomplexa,Coccidia,Eucoccidiorida,Adeleorina)inferredusing18SrDNAsequences.
JEukaryotMicrobiol.
2012;59:171–80.
50.
Criado-FornelioA,RuasJL,CasadoN,FariasNA,SoaresMP,etal.
NewmoleculardataonmammalianHepatozoonspecies(Apicomplexa:Adeleorina)fromBrazilandSpain.
IntJParasitol.
2006;92:93–9.
51.
Criado-FornelioA,BulingA,CasadoN,GimenezC,RuasJ,etal.
Molecularcharacterizationofarthropod-bornehaematozoansinwildmammalsfromBrazil,VenezuelaandSpain.
ActaParasitol.
2009;54:187–93.
52.
NetherlandsEC,CookCA,SmitNJ.
Hepatozoonspecies(Adeleorina:Hepatozoidae)ofAfricanbufonids,withmorphologicaldescriptionandmoleculardiagnosisofHepatozoonixoxosp.
nov.
parasitisingthreeAmietophrynusspecies(Anura:Bufonidae).
ParasitVectors.
2014;7:552.
53.
SlobodaM,KamlerM,BulantováJ,VotpkaJ,ModrD.
AnewspeciesofHepatozoon(Apicomplexa:Adeleorina)fromPythonregius(Serpentes:Pythonidae)anditsexperimentaltransmissionbyamosquitovector.
JParasitol.
2007;93:1189–98.
54.
MaiaJP,CrottiniA,HarrisDJ.
MicroscopicandmolecularcharacterizationofHepatozoondomerguei(Apicomplexa)andFoleyellafurcata(Nematoda)inwildendemicreptilesfromMadagascar.
Parasite.
2014;21:47.
55.
HarrisDJ,Damas-MoreiraI,MaiaJPMC,PereraA.
FirstreportofHepatozoon(Apicomplexa:Adeleorina)incaecilians,withdescriptionofanewspecies.
JParasitol.
2014;100:117–20.
56.
DvoákováN,KvierováJ,PapouekI,JavanbakhtH,TiarG,KamiH,irokP.
HaemogregarinesfromwesternPalaearcticfreshwaterturtles(generaEmys,Mauremys)areconspecificwithHaemogregarinastepanowiDanilewsky,1885.
Parasitology.
2013;141:522–30.
57.
DvoákováN,KvierováJ,HostovskM,irokP.
HaemogregarinesoffreshwaterturtlesfromSoutheastAsiawithadescriptionofHaemogregarinasacaliaesp.
n.
andaredescriptionofHaemogregarinapellegriniLaveranandPettit,1910.
Parasitology.
2015;142:816–26.
58.
KopenáJ,JirkM,OborníkM,TokarevYS,LukeJ,ModrD.
Phylogeneticanalysisofcoccidianparasitesfrominvertebrates:searchformissinglinks.
Protist.
2006;157:173–83.
Weacceptpre-submissioninquiriesOurselectortoolhelpsyoutondthemostrelevantjournalWeprovideroundtheclockcustomersupportConvenientonlinesubmissionThoroughpeerreviewInclusioninPubMedandallmajorindexingservicesMaximumvisibilityforyourresearchSubmityourmanuscriptatwww.
biomedcentral.
com/submitSubmityournextmanuscripttoBioMedCentralandwewillhelpyouateverystep:Cooketal.
Parasites&Vectors(2016)9:347Page13of13
- suit55xoxo.com相关文档
- Reimer55xoxo.com
- reduction55xoxo.com
- Fully55xoxo.com
- part55xoxo.com
- Powering55xoxo.com
- 1.0055xoxo.com
云如故枣庄高防(49元)大内存2H2G49元8H8G109元
云如故是一家成立于2018年的国内企业IDC服务商,由山东云如故网络科技有限公司运营,IDC ICP ISP CDN VPN IRCS等证件齐全!合法运营销售,主要从事自营高防独立服务器、物理机、VPS、云服务器,虚拟主机等产品销售,适合高防稳定等需求的用户,可用于建站、游戏、商城、steam、APP、小程序、软件、资料存储等等各种个人及企业级用途。机房可封UDP 海外 支持策略定制 双层硬件(傲...

阿里云年中活动最后一周 - ECS共享型N4 2G1M年付59元
以前我们在参与到云服务商促销活动的时候周期基本是一周时间,而如今我们会看到无论是云服务商还是电商活动基本上周期都要有超过一个月,所以我们有一些网友习惯在活动结束之前看看商家是不是有最后的促销活动吸引力的,比如有看到阿里云年中活动最后一周,如果我们有需要云服务器的可以看看。在前面的文章中(阿里云新人福利选择共享性N4云服务器年79.86元且送2月数据库),(LAOZUO.ORG)有提到阿里云今年的云...
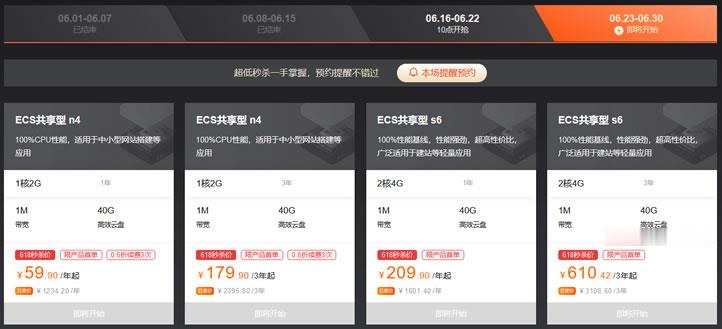
HostDare($33.79/年)CKVM和QKVM套餐 可选CN2 GIA线路
关于HostDare服务商在之前的文章中有介绍过几次,算是比较老牌的服务商,但是商家背景财力不是特别雄厚,算是比较小众的个人服务商。目前主流提供CKVM和QKVM套餐。前者是电信CN2 GIA,不过库存储备也不是很足,这不九月份发布新的补货库存活动,有提供九折优惠CN2 GIA,以及六五折优惠QKVM普通线路方案。这次活动截止到9月30日,不清楚商家这次库存补货多少。比如 QKVM基础的五个方案都...
55xoxo.com为你推荐
-
乐划锁屏乐视屏幕手机解锁忘记但是指纹可以解开怎么办?johncusack有喜欢演员JOHN CUSACK的吗?从哪部片子开始喜欢他的?至今为止他主要参与的电影作品有哪些?嘉兴商标注册怎么查商标注册日期百度关键词价格查询百度推广关键词怎么扣费?陈嘉垣陈嘉桓是谁?百度指数词百度指数为0的词 为啥排名没有m.kan84.net经常使用http://www.feikan.cc看电影的进来帮我下啊www.5any.comwww.qbo5.com 这个网站要安装播放器lcoc.toptop weenie 是什么?baqizi.cc徐悲鸿到其中一张很美的女人体画