microbialcydia
cydia 闪退 时间:2021-02-27 阅读:()
RESEARCHOpenAccessAerobicgutbacterialfloraofCydiapomonella(L.
)(Lepidoptera:Tortricidae)andtheirvirulencetothehostElifSevim1,Mustafaocar2,FikriyeMilletliSezgin3andAliSevim1*AbstractThisstudyaimedtoisolateandcharacterizebacteriafromthecodlingmoth,Cydiapomonella(L.
)(Lepidoptera:Tortricidae),anddeterminetheirvirulencetoitslarvae.
Atotalof16bacteriawereisolatedfromlarvaebelongingtodifferentinstars.
Basedonmorphological,biochemical,physiological,andmolecularstudies,thebacterialisolateswereidentifiedasPseudomonassp.
(Cp1,3,5,and13),Corynebacteriumsp.
(Cp2),Bacillussp.
(Cp4,7,9,10,12,and15),Pectobacteriumcarotovorum(Cp6),Paenibacillussp.
(Cp8),Bacillusmegaterium(Cp11),Bacilluspumilus(Cp14),andTerribacillussaccharophilus(Cp16).
Itwasalsodeterminedthevirulenceoftheseisolates,wherethehighestpotentialactivitywasobtainedbyBacillussp.
Cp9,with(76%)mortality.
TheseresultscouldbebeneficialforfuturebiocontrolprogramsofC.
pomonella.
Keywords:Bacteria,Codlingmoth,Cydiapomonella,Virulence,MicrobialcontrolBackgroundThecodlingmoth,Cydiapomonella(L.
)(Lepidoptera:Tortricidae),isoneofthemostimportantpestsinmanyorchardsworldwide,mainlyapples,pears,quince,peach,plum,apricot,andwalnut.
Itcauseseconomiclossesinfruitproduction(Pajacetal.
2012andAlford2014).
ThelarvaeofthispestwhichoverwinterinthecrackedbarkoftreetrunksandincocoonsatpackagingandstorageplacesdeveloptopupaeinlateApriltoearlyMay.
Thematedfemalemothsemergingfrompupaelayeggsatappropriatetemperatures.
Thehatchedlarvaeburrowintothefruitwithin4–8handrenderthemunsalable(Beersetal.
2003).
Inordertocontrolthispestandtoobtainundamagedfruits,traditionalinsecticidessuchasorganophosphoruscompoundsandsyntheticpyrethroidshavebeenused.
However,theseinsecticidescauseun-favorableenvironmentalimpacts.
Inaddition,somestrainsofthispesthaveacquiredresistancetoseveralinsecticides(LaceyandUnruh2005).
Entomopathogenicmicroorganismssuchasbacteria,viruses,fungi,nematodes,andprotistsareabletoinfectdifferentinsectspecies,andtheycanbeusedasbio-logicalcontrolagentsagainstinsectpests(Khetan2001).
Amongentomopathogens,theentomopathogenicbac-teria(EPB)playakeyroleinthecommercialcontrolofinsectpestsandBacillusthuringiensis(Bt)isthespeciesonwhichmostofthescientificcommunityandindustryeffortshavebeenfocused(Owuama2001andRuiuetal.
2013).
ApartfromBt,manydifferentEPBbelongingtodifferentspeciesofBacillusandothergenera,suchasBacillussphaericus,Paenibacilluspapillae,andSerratiaentomophila,areavailableasinsecticides(Federici2007).
Manyinsectlifecyclesareassociatedwithsymbioticmicroorganisms,andthereisincreasingevidencethatsymbioticmicroorganismsinfluencemanyinsectfeaturessuchassexdetermination,nutrientexchange,nutrition,anddigestionprocesses(Rajagopal2009;Douglas2014;andBrune2014).
However,someinsectgroupsarenotobligatorydependentontheirmicrobiota(Douglas2014).
Symbioticmicroorganisms,especiallybacteria,canbeusedinthebiologicalcontrolofinsectpeststhroughtheuseofdifferentmethods,e.
g.
,theycanbeusedtoexpressinsecticidaltoxinsorproteinsbyusinggeneticengineeringtechniques(Beardetal.
1998).
Inaddition,changingthedynamicsamongbacterialmicrobesintheinsectgutcouldbeusedforcontrollinginsectpests.
Fordifferent*Correspondence:ali.
sevim@ahievran.
edu.
tr1FacultyofEngineeringandArchitecture,GeneticandBioengineering,AhiEvranUniversity,40100Krehir,TurkeyFulllistofauthorinformationisavailableattheendofthearticleEgyptianJournalofBiologicalPestControlTheAuthor(s).
2018OpenAccessThisarticleisdistributedunderthetermsoftheCreativeCommonsAttribution4.
0InternationalLicense(http://creativecommons.
org/licenses/by/4.
0/),whichpermitsunrestricteduse,distribution,andreproductioninanymedium,providedyougiveappropriatecredittotheoriginalauthor(s)andthesource,providealinktotheCreativeCommonslicense,andindicateifchangesweremade.
Sevimetal.
EgyptianJournalofBiologicalPestControl(2018)28:30https://doi.
org/10.
1186/s41938-018-0036-1purposes,themicrobiotaofmanyinsectspecieshasbeendetermined(Sevimetal.
2012;Demircietal.
2013;andRoopaetal.
2014).
ThisstudyaimedtoisolateandcharacterizebacterialspeciesfromC.
pomonella.
Additionally,thesebacterialisolatesweretestedagainstthelarvaeofthecodlingmoth.
MethodsCollectionoflarvaeDifferentlarvalinstarsofC.
pomonellawerecollectedfrominfestedwalnutfruitsatthevicinityofKrehir,Turkey,inthesummerof2015.
Theobtainedlarvaewereseparatedaccordingtothedevelopmentalstage(instars)andusedintheprocessofbacterialisolation.
IsolationofbacteriaThecollectedlarvaeweredividedintothreegroupsbasedontheirinstars.
Thefirstgroupconsistedofthefirstandsecondlarvalinstars,thesecondgroupcon-sistedofthethirdinstar,andthethirdgroupconsistedofthefourthandfifthlarvalinstars.
Thebacterialiso-lationwasseparatelyperformedfromthesegroups.
Atotaloftenlarvaewereusedforeachgroupforthebacterialisolation.
Thelarvaeweresurface-sterilizedwith70%ethanolfor2–3minandwashedthreetimeswithsteriledistilledwater(LipaandWiland1972).
Thereafter,thelarvaebelongingtothedifferentgroupswereseparatelyplacedintoglasstesttubes(10ml)includ-ing3mlnutrientbroth(Difco,NJ,USA)withsterilefor-cepsandcompletelyhomogenized,usingasterileglasstissuegrinder.
Thehomogenateswerefilteredthroughtwolayersofsterilecheeseclothtoremoveinsectdeb-ris.
Aseriesofdilutionsfrom101to108werepre-paredfromtheinsecthomogenates,and101,103,105,and108dilutionsfromeachhomogenatewereplatedonnutrientagarandthenincubatedat30°Cfor3days.
Inaddition,thesedilutionswereheatedat80°Cfor10mintoeliminatenon-spore-formingbacteria.
Then,theywereplatedonnutrientagarandincubatedat30°Cfor3days;then,thebacterialcolonieswerecounted,andthetotalnumberofbacteriaperlarvaewascalculatedas2*106cfu(colonyformingunits).
Moreover,thedif-ferentbacterialcolonieswerestreakedonnutrientagarandincubatedat30°Cfor18and48hforslow-growingisolatestoobtainpurecultures.
Theobtainedpurecultureswerestoredin20%glycerolat20°C.
Thebacterialisolateswereidentifiedbasedonvarioustests.
Allisolatesfromthisstudyarepubliclyaccess-ibleandweredepositedatMicrobiologyLaboratory,GeneticandBioengineering,AhiEvranUniversity,Krehir,Turkey.
MorphologicalcharacterizationofthebacterialisolatesThebacterialisolatesweremorphologicallycharacter-izedonthebasisoftheircolony,cell,andsporefea-tures.
Colonymorphologiesofthebacterialisolateswereevaluatedonnutrientagarplatesbyusingastereomicroscope(Demircietal.
2013).
Cellpropertiesoftheisolateswereevaluatedbythegramandendo-sporestaining.
Thecapsulelayerofthebacterialiso-lateswasdeterminedbynegativestaining.
ThemotilityoftheisolateswasdeterminedaccordingtothemethodofSoutourinaetal.
(2001).
PhysiologicalcharacterizationofthebacterialisolatesThebacterialisolateswerealsophysiologicallycharacter-izedonthebasisoftheirgrowthatdifferenttempera-tures,NaClconcentrations,andpH.
Allisolateswereinoculatedintonutrientbroth(3ml)andincubatedatdifferenttemperaturesrangingfrom4to55°C.
Also,allisolateswereincubatedintonutrientbroth(3ml)withdifferentconcentrationsofNaCl,rangingfrom3to15%.
Finally,allisolateswereincubatedintonutrientbroth(3ml)withdifferentpHvaluesrangingfrom3to12.
Evaluationswerevisuallymade.
VITEK2microbialidentificationsystemThebacterialisolateswerealsoidentifiedusingtheVITEK2microbialidentificationsystem.
Firstly,thebacterialiso-lateswerestreakedonnutrientagarplatestoobtainsinglecolonies.
Thebacterialsuspensionswerepreparedfromasinglecolony,using2mlof0.
45%sterilesalinesolutiontotheequivalentofa0.
5McFarlandturbiditystandard.
ConcentrationswerecheckedwiththeVITEKcolorimeterforeachisolate.
Additionally,theoxidaseandcatalasepro-ductionoftheisolatesweremanuallydetermined.
Atotaloftwocardswereusedtoidentifytheisolates.
TheGNIDcardwasusedforgram-negativebacterialidentificationandtheGPIDcardwasusedforgram-positivebacterialidentification.
Thebacterialsuspensionspreparedasabovewereinoculatedontothesecardsandincubatedat30°Cfor18h.
Thetimebetweenpreparationofthesuspen-sionandcardfillingwaslessthanhalfanhour.
TheresultswereautomaticallyevaluatedwiththeVITEK2device(Ligozzietal.
2002).
16SrRNAgenesequencingThebacterialisolateswerefurthercharacterized,usingthepartialsequencingof16SrRNAgene.
GenomicDNAswereextractedbytheGenomicDNAisolationkit(ThermoFisherScientific,Waltham,MA,USA).
TheextractedDNAswerestoredat20°CuntilPCRwasdone.
Approximately1.
450bpofthe16SrRNAgenere-gionwastargetedandamplified.
Theprimerpairsof27F(5′-AGAGTTTGATCMTGGCTCAG-3′asforward)and1492R(5′-GGYTACCTTGTTACGACTT-3′asreverse)Sevimetal.
EgyptianJournalofBiologicalPestControl(2018)28:30Page2of9werepurchasedfromMACROGENandusedforamplifi-cation.
ThetotalvolumeofPCRreactionswas50μltowhich50–100nggenomicDNAwasadded.
ThePCRmixof50μlpersamplecontained25pmolofeachprimer,200mMeachofthedNTPs,1*PCRbuffer,3mMMgCl2and1.
5UTaqDNApolymerase.
Afteraddingallthecomponents,thefinalvolumewasadjustedto50μlwithsteriledistilledwater.
ThePCRprogramconsistedof95°C(60s)fortheinitialdenaturation,followedby35cyclesof94°C(45s)fordenaturation,55°C(30s)forannealing,72°C(1.
5min)forextension,andafinalextensionof72°C(5min).
AfterperformingPCR,5mloftheproductswasanalyzedbyelectrophoresison1.
0%agarosegelcontainingethidiumbromidetochecktheTable1ThemorphologicalpropertiesofthebacterialisolatesIsolateColonyshapeColonycolorGramstainingCellshapeSporestainingMotilityCapsuleGrowthinNBaInstarCp1SmoothCream–Bacil––+Turbid1–2Cp2SmoothCream+Coccus–––Turbid1–2Cp3SmoothYellow–Coccus–––Turbid1–2Cp4SmoothPink+Bacil–––Turbid1–2Cp5SmoothYellow–Bacil–––Turbid1–2Cp6SmoothCream–Bacil–+–Turbid3Cp7SmoothCream+Bacil++–Turbid3Cp8SmoothCream+Bacil++–Turbid3Cp9RoughCream+Bacil++–Turbid3Cp10SmoothCream+Bacil++–Turbid3Cp11RoughCream+Bacil–––Turbid3Cp12MucoidCream+Bacil–+–Precipitated3Cp13SmoothYellow–Bacil–++Turbid3Cp14SmoothCream+Bacil+––Turbid4–5Cp15SmoothCream+Bacil–+–Turbid4–5Cp16SmoothCream+Bacil++–Turbid4–5aNutrientbrothTable2Thephysiologicalpropertiesofthebacterialisolates.
Luria-BertanibrothwasusedasgrowthmediumIsolateGrowthNaCl(%)pHTemperature(°C)3571012153456789101243037455055Cp1Cp2Cp3Cp4Cp5Cp6Cp7Cp8Cp9Cp10Cp11Cp12Cp13Cp14Cp15Cp16Sevimetal.
EgyptianJournalofBiologicalPestControl(2018)28:30Page3of9sizesandamountsoftheamplicons.
AftercheckingPCRproducts,theaccurateproductsweresenttoMACRO-GEN(theNetherlands)forsequencing.
ThePCRproductsweresequencedwiththeprimerpairs518F(5′-CCAGCAGCCGCGGTAATACG-3′)and800R(5′-TACCAGGGTATCTAATCC-3′).
Theobtainedsequencesweresub-jectedtothenucleotideBLASTsearchesintheNCBIGenBankdatabasetogetthepercentagesimilarityofthebacterialisolatestothemostrelatedbacterialspecies(Altschuletal.
1990).
Table3PercentsimilarityofthebacterialisolateswiththeircloselyrelatedspeciesbasedontheBLASTsearchesinNCBIGanBankdatabase(Altschuletal.
1990)IsolateBacterialspeciesGenBankaccessionnumberQuerycover(%)Similarity(%)VITEK2(%)Cp1Pseudomonassp.
PDD-59b-7Pseudomonassp.
R3ScM3P1C11PseudomonassyringaestrainPDD-48b-5KR922145KF147001KR922059969696999999Lysinibacillussphaericus/Lysinibacillusfusiformis(91)Cp2CorynebacteriumvariabilestrainC3-13Corynebacteriumsp.
ZT10-3CorynebacteriumvariabileDSM44702KP114214KT597082NR102874969696969696Gardnerellavaginalis(93)Cp3Pseudomonassp.
MN11-3PseudomonasmatsuisolistrainCC-MHH0089PseudomonasmatsuisolistrainCC-MHH0089JQ396614NR134793KJ720680979797969696Dermacoccusnishinomiyaensis/Kytococcussedentarius(96)Cp4BacteriumBELC12Bacillussp.
13K7a2Bacillussp.
7Kp1aBacilluspumilusstrainOU101KT382407KT825840KT825839KR1403779696969699999999Lysinibacillussphaericus/Lysinibacillusfusiformis(87)Cp5PseudomonasmatsuisolistrainCC-MHH0089PseudomonasmatsuisolistrainCC-MHH0089Pseudomonassp.
RBSB9_C3NR134793KJ720680KT390731979796969697Aeromonassalmonicida(98)Cp6Pectobacteriumcarotovorumsubsp.
brasiliensestrainY4516SPectobacteriumcarotovorumsubsp.
brasiliensestrainY3416SPectobacteriumcarotovorumsubsp.
brasiliensestrainY3316SKP187510KP187504KP187503989898979797UnidentifiedorganismCp7Bacillussp.
BG2-9BacilluspumilusstrainT246BacilluspumilusstrainML353KP992115KC764989KC692160989999979797Bacilluspumilus(85)Cp8Paenibacillussp.
MOLA507BacteriumUKRA17Paenibacillussp.
S8AM990732KT382376KR051059969696979797Paenibacilluspolymyxa(90)Cp9Bacillussp.
210_50Bacillussp.
strainRHH15Bacillussp.
L11(2010)GQ199752HQ143613HQ222333999999969696Bacilluspumilus(86)Cp10BacilluspumilusstrainX22BacilluspumilusstrainHN-30BacilluspumilusstrainHN-10FJ763645KT003271KT003256999999989898Bacilluspumilus(88)Cp11BacillusmegateriumstrainD5BacillusmegateriumstrainBCRh8BacillusmegateriumstrainBS9KC441754KT153604KR063189999999989898Bacillusmegaterium(87)Cp12Bacillussp.
C26(2014)BacillussubtilisstrainL-13BacillustequilensisstrainYJ-S4KM117217HQ232422KF876849999999999999UnidentifiedorganismCp13Pseudomonassp.
BE07Unculturedbacteriumisolate1112863845131Pseudomonassp.
DR11(2011)AY456700HQ121073JN210571979797969696Aeromonassalmonicida(97)Cp14BacilluspumilusstrainZA13BacilluspumilusstrainLX11BacilluspumilusstrainY13FJ263042KP192031KF641806989898999999UnidentifiedorganismCp15BacillussubtilisstrainL-13BacillussubtilisstrainYA4BZBacillussp.
RKZ11262HQ232422JQ346075EU835569989898999999UnidentifiedorganismCp16TerribacillussaccharophilusstrainMER_108TerribacillussaccharophilusstrainJP44SK46TerribacillussaccharophilusstrainWA2-4KT719683JX155763JF496471969696969696UnidentifiedorganismSevimetal.
EgyptianJournalofBiologicalPestControl(2018)28:30Page4of9PhylogeneticanalysisPhylogeneticanalysisofthebacterialisolatesandtheircloselyrelatedspecieswasperformedformolecularcharacterizationofthebacterialisolates.
ThesequenceswereeditedusingBioedit,andmultiplesequencealign-mentswerecreatedbyusing16SrRNAsequencesbe-longingtoourstrainsanddifferentbacterialspeciesfromtheNCBIGenBankdatabaseforthepurposeofdevelopingaphylogenetictree.
ThemultiplesequencealignmentwasperformedwithClustalWinBioedit(Hall1999).
Finally,thesequencesweresubjectedtoneighbor-joininganalysiswithp-distancecorrection,gapomission,and1.
000bootstrappseudoreplicatesusingMEGA6.
0(Tamuraetal.
2013).
NucleotidesequenceaccessionnumbersTheGenBankaccessionnumbersofthe16SrRNAgenesequencesbelongingtothebacterialisolatesfromthisstudyarelistedinTable4.
BioassayEachbacteriumisolatedfromC.
pomonellainthestockculturewasstreakedonnutrientagartoobtainasinglecolonyandtocheckthepurityofthecultures.
Afterthat,3mlofnutrientbrothwasinoculatedfromeachsinglecolonyof16bacteriaandincubatedat30°Cover-night.
Attheendoftheincubationperiod,thebacterialdensitywasmeasuredat600nmabsorbanceandadjustedto1.
8*109cfu/mlbycentrifugation(4.
000rpmfor15min)andusingsterilephosphatebuffersolution(PBS)(Moaretal.
1995).
Thebacterialsolutionswerefreshlypreparedandusedforbioassay.
Forthebioassayexperiments,healthyC.
pomonellalarvaewereobtainedfromthelaboratorycultureatAhiEvranUniversity,GeneticBioengineeringandMicrobiologyLaboratory.
Healthylarvaewererandomlyselectedandusedforthebioassay.
Twenty-fivegramsoffreshlypre-paredartificialdiet(fordietingredientsandrearingcondi-tions(Fukovaetal.
2005))wasinoculatedwith1mlofthebacterialsuspensionpreparedasdescribedaboveforeachisolate.
Forthecontrolgroup,25goftheartificialdiethad1mlofthesterilePBSadded.
Thecontami-natedartificialdietswereseparatelyplacedintoplasticboxes(20*10*8mm)withventilatedlidstopermitairflow.
Afterthat,tenthirdinstarC.
pomonellalarvaewereplacedintotheboxforeachreplicateandallowedtofeedonthecontaminateddiets.
Atotaloftenlarvaewereusedforeachreplicate,andallexperimentswererepeatedthreetimes.
Finally,theplasticboxeswerein-cubatedat25°Cunder16:8(day:night)lightregime.
After10days,theboxeswerecheckedforlarvalmor-tality,andthenumberofdeadlarvaewasrecorded.
MortalitydatawerecorrectedbasedonAbbott'sfor-mula(Abbott1925).
Todeterminethedifferencesamongtheisolatesandthecontrolgroup,thedataweresubjectedtoANOVAandsubsequentlytotheLSDmultiplecomparisontest.
BeforeperformingANOVA,alldataweretestedforhomogeneityofvarianceusingLevene'sstatistic.
AlltestswereperformedwithSPSS16.
0statisticalsoftware.
ResultsanddiscussionAtotalof16isolatesofbacteriawereobtainedfromthetreatedC.
pomonellalarvae.
Amongtheseisolates,fivewerefromthefirstandsecondlarvalinstars,eightfromthethirdlarvalinstar,andthreefromthefourthandfifthlarvalinstars.
Theisolateswerecharacterizedonthebasisoftheirmorphological,physiological,andmolecularcharacteristics.
Coloniesofallisolatesweresmooth,exceptforCp9andCp11,whichwererough.
Onlyoneisolate(Cp12)hadamucoidcolony.
Threeisolates(Cp3,Cp5,andCp13)hadyellowcolonies,andoneisolate(Cp4)hadapinkcolony.
Theotherisolatesproducedcreamy-coloredcolonies.
Fiveisolates(Cp1,Cp3,Cp5,Cp6,andCp13)weregram-negative,andtheothersweregram-positive.
Allisolateswerebacilli-shaped,exceptforCp2andCp3whichwerecoccusshaped.
Itwasfoundthatsixisolates(Cp7,Cp8,Cp9,Cp10,Cp14,andCp16)formedspores.
Onlytwoisolates(Cp1andCp13)hadcapsules.
Allmorphologicalcharac-teristicsofthebacterialisolatesaregiveninTable1.
Table4TheproposedidentificationresultsofthebacterialisolatesandtheirGenBankaccessionnumbersfor16SrRNAgenesequencesIsolateSpeciesGenBankaccessionnumberCp1Pseudomonassp.
KX094470Cp2Corynebacteriumsp.
KX094471Cp3Pseudomonassp.
KX094472Cp4Bacillussp.
KX094473Cp5Pseudomonassp.
KX094474Cp6PectobacteriumcarotovorumKX094475Cp7Bacillussp.
KX094476Cp8Paenibacillussp.
KX094477Cp9Bacillussp.
KX094478Cp10Bacillussp.
KX094479Cp11B.
megateriumKX094480Cp12Bacillussp.
KX094481Cp13Pseudomonassp.
KX094482Cp14B.
pumilusKX094483Cp15Bacillussp.
KX094484Cp16TerribacillussaccharophilusKX094485Sevimetal.
EgyptianJournalofBiologicalPestControl(2018)28:30Page5of9Allisolateswereabletogrowin3%NaCl,andonlyoneisolate(Cp3)couldnotgrowin4%NaCl.
Growthcharac-teristicsoftheisolatesinotherNaClconcentrationswerevariable,dependingontheisolate.
AllisolateswereabletogrowinthepHrangeof3,4,and5,exceptforCp12.
AllisolateswereabletogrowatpH6and7.
GrowthcharacteristicsoftheisolatesatotherpHswerevariable,dependingontheisolate.
NoneoftheisolatesgrewatpH4and55°C,andgrowthpropertiesoftheisolatesat30,37,45,and50°Cwerevariable,Fig.
1Phylogenetictreederivedfromneighbor-joininganalysisof16SrRNAsequences(1400bp)fromthefloramembersofC.
pomonellaandtheircloselyrelatedspecies.
Bootstrapvaluesbasedon1000replicateswereindicatedabovenodes.
BootstrapvaluesC≥70arelabeled.
C.
pomonellaisolateswereindicatedwithblackcircle.
ThescaleonthebottomofthedendrogramindicatesthedegreeofdissimilaritySevimetal.
EgyptianJournalofBiologicalPestControl(2018)28:30Page6of9dependingontheisolate.
PhysiologicalpropertiesoftheisolatesaregiveninTable2.
TheVITEK2microbialcharacterizationsystemforbiochemicalcharacterizationoftheisolatesandgram-negativeandgram-positivecardswasusedtoidentifythem.
Fiveisolates(Cp6,Cp12,Cp14,Cp15,andCp16)werenotabletobecharacterizedbyVITEK2.
OtheridentificationsaregiveninTable3.
Thebacterialisolateswerealsocharacterizedonthebasisof16SrRNAgenesequencingtoverifythere-cordedconventionalcharacterizationsoftheisolates.
Basedonmolecularcharacterization,thebacterialiso-lateswereidentifiedasPseudomonassp.
(Cp1,3,5,and13),Corynebacteriumsp.
(Cp2),Bacillussp.
(Cp4,7,9,10,12,and15),Pectobacteriumcarotovorum(Cp6),Paenibacillussp.
(Cp8),Bacillusmegaterium(Cp11),Ba-cilluspumilus(Cp14),andTerribacillussaccharophilus(Cp16)(Table4).
Thisidentificationwasalsosupportedbyphylogeneticanalysis(Fig.
1).
Allisolatescauseddifferentmortalityvaluesincom-parisontoeachother(F=15.
43,df=16,pCydiapomonellaL.
(Lepidoptera:Tortricidae).
AfrJBiotechnol5(22):2081–2085EuzebyJP(1997)Listofbacterialnameswithstandinginnomenclature:afolderavailableontheinternet.
IntJSystBacteriol47(2):590–592FedericiBA(2007)Bacteriaasbiologicalcontrolagentsforinsects:economics,engineering,andenvironmentalsafety.
In:VurroM,GresselJ(eds)Novelbiotechnologiesforbiocontrolagentenhancementandmanagement.
Springer,Dordrecht,pp25–51FukovaI,NguyenP,MarecF(2005)Codlingmothcytogenetics:karyotype,chromosomallocationofrDNA,andmoleculardifferentiationofsexchromosomes.
Genome48:1083–1092GlasnerJD,Marquez-VillavicencioM,KimHS,JahnCE,MaB,BiehlBS,RissmanAI,MoleB,YiX,YangCH,DanglJL,GrantSR,PernaNT,CharkowskiAO(2008)Niche-specificityandthevariablefractionofthePectobacteriumpan-genome.
MolPlant-MicrobeInteract21:1549–1560GnanamanickamSS(2006)Plant-associatedbacteria.
Springer,DordrechtHallTA(1999)BioEdit:auser-friendlybiologicalsequencealignmenteditorandanalysisprogramforWindows95/98/NT.
NucleicAcidsSymp41:95–98HoevenRVD,BetrabetG,ForstS(2008)CharacterizationofthegutbacterialcommunityinManducasextaandeffectofantibioticsonbacterialdiversityandnematodereproduction.
FEMSMicrobiolLett286:249–256IanKT,KennethSB,HolevaCM,RJPB(2003)Softroterwiniae:fromgenestogenomes.
MolPlantPathol4(1):17–30KhetanSK(2001)MicrobialPestControl.
MarcelDekker,NewYorkLaceyLA,UnruhTR(2005)Biologicalcontrolofcodlingmoth(Cydiapomonella,Lepidoptera:Tortricidae)anditsroleinintegratedpestmanagement,withemphasisonentomopathogens.
Vedalia12(1):33–60LalS,TabacchioniS(2009)EcologyandbiotechnologicalpotentialofPaenibacilluspolymyxa:aminireview.
IndianJMicrobiol49:2–10LeeCW,LucasS,DesomazeaudMJ(1985)Phenylalanineandtyrosinecatabolisminsomecheesecoryneformbacteria.
FEMSMicrobiolLett26(2):201–205LigozziM,BerniniC,BonoraMG,deFatimaM,ZulianiJ,FontanaR(2002)EvaluationoftheVITEK2systemforidentificationandantimicrobialsusceptibilitytestingofmedicallyrelevantgram-positivecocci.
JClinMicrobiol40(5):1681–1686LipaJJ,WilandE(1972)BacteriaisolatedfromcutwormsandtheirinfectivitytoAgrotissp.
ActaMicrobiolPol4:127–140LiuW,JiangL,GuoC,YangSS(2010)Terribacillusaidingensissp.
nov.
,amoderatelyhalophilicbacterium.
IntJSystEvolMicrobiol60:2940–2945MaB,HibbingME,KimHS,ReedyRM,YedidiaI,BreuerJ,BreuerJ,GlasnerJD,PernaT,KelmanA,CharkowskiAO(2007)HostrangeandmolecularphylogeniesofthesoftrotenterobacterialgeneraPectobacteriumandDickeya.
Phytopathology97(9):1150–1163MaharAN,DarbanDA,GowenSR,HagueNGM,JanND,MunirM,MaharAQ(2005)UseofentomopathogenicbacteriumPseudomonasputida(Enterobacteriaceae)anditssecretionagainstgreaterwaxmoth,Galleriamellonellapupae.
JEntomol2(1):77–85McSpaddenGardenerBB(2004)EcologyofBacillusandPaenibacillusspp.
inagriculturalsystems.
Phytopathology94:1252–1258MenezesCB,Bonugli-SantosRC,MiquelettoPB,PassariniMR,SilvaCH,JustoMR,LealRR,Fantinatti-GarbogginiF,OliveiraVM,BerlinckRG,SetteLD(2010)Microbialdiversityassociatedwithalgae,ascidiansandspongesfromthenorthcoastofSoPaulostate,Brazil.
MicrobiolRes165:466–482MoarWJ,Pusztzai-CareyM,MackTP(1995)ToxicityofpurifiedproteinsandtheHD-1strainfromBacillusthuringiensisagainstlessercornstalkborer(Lepidoptera:Pyralidae).
JEconEntomol88:606–609OwuamaCI(2001)Entomopathogenicsymbioticbacteria,XenorhabdusandPhororhabdusofnematodes.
WorldJMicrobiolBiotechnol17:505–515PajacI,BaricB,MikacK,PejicI(2012)NewinsightsintothebiologyandecologyofCydiapomonellafromappleorchardsinCroatia.
BullInsectol65(2):185–193RajagopalR(2009)Beneficialinteractionsbetweeninsectsandgutbacteria.
IndianJMicrobiol49:114–119RoopaHK,RebijithKB,AsokanR,MahmoodR,KumarK(2014)Isolationandidentificationofculturablebacteriafromhoneydewofwhitefly,Bemisiatabaci(G.
)(Hemiptera:Aleyrodidae).
MetaGene2:114–122RuiuL(2015)Insectpathogenicbacteriainintegratedpestmanagement.
Insects6:352–367RuiuL,SattaA,FlorisI(2013)Emergingentomopathogenicbacteriaforinsectpestmanagement.
BullInsectol66(2):181–186SevimE,elebi,SevimA(2012)DeterminationofthebacterialfloraasamicrobialcontrolagentofToxopteraaurantii(Homoptera:Aphididae).
Biologia67(2):397–404SoutourinaOA,SemenovaEA,ParfenovaVV,DanchinA,BertinP(2001)ControlofbacterialmotilitybyenvironmentalfactorsinpolarlyflagellatedandperitrichousbacteriaisolatedfromLakeBaikal.
ApplEnvironMicrobiol67:3852–3859TamuraK,StecherG,PetersonD,FilipskiA,KumarS(2013)MEGA6:molecularevolutionarygeneticsanalysisversion6.
0.
MolBiolEvol30:2725–2729VicenteCSL,NascimentoFX,EspadaM,BarbosaP,HasegawaK,MotaM,OliveiraS(2013)CharacterizationofbacterialcommunitiesassociatedwiththepinesawyerbeetleMonochamusgalloprovincialis,theinsectvectorofthepinewoodnematodeBursaphelenchusxylophilus.
FEMSMicrobiolLett347:130–139YamadaK,KinoshitaS,TsunodaT,AidaK(1972)Themicrobialproductionofaminoacids.
Wiley,NewYorkYunJH,RohSW,WhonTW,JungMJ,KimMS,ParkDS,YoonC,NamYD,KimYJ,ChoiJH,KimJY,ShinNR,KimSH,LeeWJ,BaeJW(2014)Insectgutbacterialdiversitydeterminedbyenvironmentalhabitat,diet,developmentalstage,andphylogenyofhost.
ApplEnvironMicrobiol80(17):5254–5264Sevimetal.
EgyptianJournalofBiologicalPestControl(2018)28:30Page9of9
)(Lepidoptera:Tortricidae)andtheirvirulencetothehostElifSevim1,Mustafaocar2,FikriyeMilletliSezgin3andAliSevim1*AbstractThisstudyaimedtoisolateandcharacterizebacteriafromthecodlingmoth,Cydiapomonella(L.
)(Lepidoptera:Tortricidae),anddeterminetheirvirulencetoitslarvae.
Atotalof16bacteriawereisolatedfromlarvaebelongingtodifferentinstars.
Basedonmorphological,biochemical,physiological,andmolecularstudies,thebacterialisolateswereidentifiedasPseudomonassp.
(Cp1,3,5,and13),Corynebacteriumsp.
(Cp2),Bacillussp.
(Cp4,7,9,10,12,and15),Pectobacteriumcarotovorum(Cp6),Paenibacillussp.
(Cp8),Bacillusmegaterium(Cp11),Bacilluspumilus(Cp14),andTerribacillussaccharophilus(Cp16).
Itwasalsodeterminedthevirulenceoftheseisolates,wherethehighestpotentialactivitywasobtainedbyBacillussp.
Cp9,with(76%)mortality.
TheseresultscouldbebeneficialforfuturebiocontrolprogramsofC.
pomonella.
Keywords:Bacteria,Codlingmoth,Cydiapomonella,Virulence,MicrobialcontrolBackgroundThecodlingmoth,Cydiapomonella(L.
)(Lepidoptera:Tortricidae),isoneofthemostimportantpestsinmanyorchardsworldwide,mainlyapples,pears,quince,peach,plum,apricot,andwalnut.
Itcauseseconomiclossesinfruitproduction(Pajacetal.
2012andAlford2014).
ThelarvaeofthispestwhichoverwinterinthecrackedbarkoftreetrunksandincocoonsatpackagingandstorageplacesdeveloptopupaeinlateApriltoearlyMay.
Thematedfemalemothsemergingfrompupaelayeggsatappropriatetemperatures.
Thehatchedlarvaeburrowintothefruitwithin4–8handrenderthemunsalable(Beersetal.
2003).
Inordertocontrolthispestandtoobtainundamagedfruits,traditionalinsecticidessuchasorganophosphoruscompoundsandsyntheticpyrethroidshavebeenused.
However,theseinsecticidescauseun-favorableenvironmentalimpacts.
Inaddition,somestrainsofthispesthaveacquiredresistancetoseveralinsecticides(LaceyandUnruh2005).
Entomopathogenicmicroorganismssuchasbacteria,viruses,fungi,nematodes,andprotistsareabletoinfectdifferentinsectspecies,andtheycanbeusedasbio-logicalcontrolagentsagainstinsectpests(Khetan2001).
Amongentomopathogens,theentomopathogenicbac-teria(EPB)playakeyroleinthecommercialcontrolofinsectpestsandBacillusthuringiensis(Bt)isthespeciesonwhichmostofthescientificcommunityandindustryeffortshavebeenfocused(Owuama2001andRuiuetal.
2013).
ApartfromBt,manydifferentEPBbelongingtodifferentspeciesofBacillusandothergenera,suchasBacillussphaericus,Paenibacilluspapillae,andSerratiaentomophila,areavailableasinsecticides(Federici2007).
Manyinsectlifecyclesareassociatedwithsymbioticmicroorganisms,andthereisincreasingevidencethatsymbioticmicroorganismsinfluencemanyinsectfeaturessuchassexdetermination,nutrientexchange,nutrition,anddigestionprocesses(Rajagopal2009;Douglas2014;andBrune2014).
However,someinsectgroupsarenotobligatorydependentontheirmicrobiota(Douglas2014).
Symbioticmicroorganisms,especiallybacteria,canbeusedinthebiologicalcontrolofinsectpeststhroughtheuseofdifferentmethods,e.
g.
,theycanbeusedtoexpressinsecticidaltoxinsorproteinsbyusinggeneticengineeringtechniques(Beardetal.
1998).
Inaddition,changingthedynamicsamongbacterialmicrobesintheinsectgutcouldbeusedforcontrollinginsectpests.
Fordifferent*Correspondence:ali.
sevim@ahievran.
edu.
tr1FacultyofEngineeringandArchitecture,GeneticandBioengineering,AhiEvranUniversity,40100Krehir,TurkeyFulllistofauthorinformationisavailableattheendofthearticleEgyptianJournalofBiologicalPestControlTheAuthor(s).
2018OpenAccessThisarticleisdistributedunderthetermsoftheCreativeCommonsAttribution4.
0InternationalLicense(http://creativecommons.
org/licenses/by/4.
0/),whichpermitsunrestricteduse,distribution,andreproductioninanymedium,providedyougiveappropriatecredittotheoriginalauthor(s)andthesource,providealinktotheCreativeCommonslicense,andindicateifchangesweremade.
Sevimetal.
EgyptianJournalofBiologicalPestControl(2018)28:30https://doi.
org/10.
1186/s41938-018-0036-1purposes,themicrobiotaofmanyinsectspecieshasbeendetermined(Sevimetal.
2012;Demircietal.
2013;andRoopaetal.
2014).
ThisstudyaimedtoisolateandcharacterizebacterialspeciesfromC.
pomonella.
Additionally,thesebacterialisolatesweretestedagainstthelarvaeofthecodlingmoth.
MethodsCollectionoflarvaeDifferentlarvalinstarsofC.
pomonellawerecollectedfrominfestedwalnutfruitsatthevicinityofKrehir,Turkey,inthesummerof2015.
Theobtainedlarvaewereseparatedaccordingtothedevelopmentalstage(instars)andusedintheprocessofbacterialisolation.
IsolationofbacteriaThecollectedlarvaeweredividedintothreegroupsbasedontheirinstars.
Thefirstgroupconsistedofthefirstandsecondlarvalinstars,thesecondgroupcon-sistedofthethirdinstar,andthethirdgroupconsistedofthefourthandfifthlarvalinstars.
Thebacterialiso-lationwasseparatelyperformedfromthesegroups.
Atotaloftenlarvaewereusedforeachgroupforthebacterialisolation.
Thelarvaeweresurface-sterilizedwith70%ethanolfor2–3minandwashedthreetimeswithsteriledistilledwater(LipaandWiland1972).
Thereafter,thelarvaebelongingtothedifferentgroupswereseparatelyplacedintoglasstesttubes(10ml)includ-ing3mlnutrientbroth(Difco,NJ,USA)withsterilefor-cepsandcompletelyhomogenized,usingasterileglasstissuegrinder.
Thehomogenateswerefilteredthroughtwolayersofsterilecheeseclothtoremoveinsectdeb-ris.
Aseriesofdilutionsfrom101to108werepre-paredfromtheinsecthomogenates,and101,103,105,and108dilutionsfromeachhomogenatewereplatedonnutrientagarandthenincubatedat30°Cfor3days.
Inaddition,thesedilutionswereheatedat80°Cfor10mintoeliminatenon-spore-formingbacteria.
Then,theywereplatedonnutrientagarandincubatedat30°Cfor3days;then,thebacterialcolonieswerecounted,andthetotalnumberofbacteriaperlarvaewascalculatedas2*106cfu(colonyformingunits).
Moreover,thedif-ferentbacterialcolonieswerestreakedonnutrientagarandincubatedat30°Cfor18and48hforslow-growingisolatestoobtainpurecultures.
Theobtainedpurecultureswerestoredin20%glycerolat20°C.
Thebacterialisolateswereidentifiedbasedonvarioustests.
Allisolatesfromthisstudyarepubliclyaccess-ibleandweredepositedatMicrobiologyLaboratory,GeneticandBioengineering,AhiEvranUniversity,Krehir,Turkey.
MorphologicalcharacterizationofthebacterialisolatesThebacterialisolatesweremorphologicallycharacter-izedonthebasisoftheircolony,cell,andsporefea-tures.
Colonymorphologiesofthebacterialisolateswereevaluatedonnutrientagarplatesbyusingastereomicroscope(Demircietal.
2013).
Cellpropertiesoftheisolateswereevaluatedbythegramandendo-sporestaining.
Thecapsulelayerofthebacterialiso-lateswasdeterminedbynegativestaining.
ThemotilityoftheisolateswasdeterminedaccordingtothemethodofSoutourinaetal.
(2001).
PhysiologicalcharacterizationofthebacterialisolatesThebacterialisolateswerealsophysiologicallycharacter-izedonthebasisoftheirgrowthatdifferenttempera-tures,NaClconcentrations,andpH.
Allisolateswereinoculatedintonutrientbroth(3ml)andincubatedatdifferenttemperaturesrangingfrom4to55°C.
Also,allisolateswereincubatedintonutrientbroth(3ml)withdifferentconcentrationsofNaCl,rangingfrom3to15%.
Finally,allisolateswereincubatedintonutrientbroth(3ml)withdifferentpHvaluesrangingfrom3to12.
Evaluationswerevisuallymade.
VITEK2microbialidentificationsystemThebacterialisolateswerealsoidentifiedusingtheVITEK2microbialidentificationsystem.
Firstly,thebacterialiso-lateswerestreakedonnutrientagarplatestoobtainsinglecolonies.
Thebacterialsuspensionswerepreparedfromasinglecolony,using2mlof0.
45%sterilesalinesolutiontotheequivalentofa0.
5McFarlandturbiditystandard.
ConcentrationswerecheckedwiththeVITEKcolorimeterforeachisolate.
Additionally,theoxidaseandcatalasepro-ductionoftheisolatesweremanuallydetermined.
Atotaloftwocardswereusedtoidentifytheisolates.
TheGNIDcardwasusedforgram-negativebacterialidentificationandtheGPIDcardwasusedforgram-positivebacterialidentification.
Thebacterialsuspensionspreparedasabovewereinoculatedontothesecardsandincubatedat30°Cfor18h.
Thetimebetweenpreparationofthesuspen-sionandcardfillingwaslessthanhalfanhour.
TheresultswereautomaticallyevaluatedwiththeVITEK2device(Ligozzietal.
2002).
16SrRNAgenesequencingThebacterialisolateswerefurthercharacterized,usingthepartialsequencingof16SrRNAgene.
GenomicDNAswereextractedbytheGenomicDNAisolationkit(ThermoFisherScientific,Waltham,MA,USA).
TheextractedDNAswerestoredat20°CuntilPCRwasdone.
Approximately1.
450bpofthe16SrRNAgenere-gionwastargetedandamplified.
Theprimerpairsof27F(5′-AGAGTTTGATCMTGGCTCAG-3′asforward)and1492R(5′-GGYTACCTTGTTACGACTT-3′asreverse)Sevimetal.
EgyptianJournalofBiologicalPestControl(2018)28:30Page2of9werepurchasedfromMACROGENandusedforamplifi-cation.
ThetotalvolumeofPCRreactionswas50μltowhich50–100nggenomicDNAwasadded.
ThePCRmixof50μlpersamplecontained25pmolofeachprimer,200mMeachofthedNTPs,1*PCRbuffer,3mMMgCl2and1.
5UTaqDNApolymerase.
Afteraddingallthecomponents,thefinalvolumewasadjustedto50μlwithsteriledistilledwater.
ThePCRprogramconsistedof95°C(60s)fortheinitialdenaturation,followedby35cyclesof94°C(45s)fordenaturation,55°C(30s)forannealing,72°C(1.
5min)forextension,andafinalextensionof72°C(5min).
AfterperformingPCR,5mloftheproductswasanalyzedbyelectrophoresison1.
0%agarosegelcontainingethidiumbromidetochecktheTable1ThemorphologicalpropertiesofthebacterialisolatesIsolateColonyshapeColonycolorGramstainingCellshapeSporestainingMotilityCapsuleGrowthinNBaInstarCp1SmoothCream–Bacil––+Turbid1–2Cp2SmoothCream+Coccus–––Turbid1–2Cp3SmoothYellow–Coccus–––Turbid1–2Cp4SmoothPink+Bacil–––Turbid1–2Cp5SmoothYellow–Bacil–––Turbid1–2Cp6SmoothCream–Bacil–+–Turbid3Cp7SmoothCream+Bacil++–Turbid3Cp8SmoothCream+Bacil++–Turbid3Cp9RoughCream+Bacil++–Turbid3Cp10SmoothCream+Bacil++–Turbid3Cp11RoughCream+Bacil–––Turbid3Cp12MucoidCream+Bacil–+–Precipitated3Cp13SmoothYellow–Bacil–++Turbid3Cp14SmoothCream+Bacil+––Turbid4–5Cp15SmoothCream+Bacil–+–Turbid4–5Cp16SmoothCream+Bacil++–Turbid4–5aNutrientbrothTable2Thephysiologicalpropertiesofthebacterialisolates.
Luria-BertanibrothwasusedasgrowthmediumIsolateGrowthNaCl(%)pHTemperature(°C)3571012153456789101243037455055Cp1Cp2Cp3Cp4Cp5Cp6Cp7Cp8Cp9Cp10Cp11Cp12Cp13Cp14Cp15Cp16Sevimetal.
EgyptianJournalofBiologicalPestControl(2018)28:30Page3of9sizesandamountsoftheamplicons.
AftercheckingPCRproducts,theaccurateproductsweresenttoMACRO-GEN(theNetherlands)forsequencing.
ThePCRproductsweresequencedwiththeprimerpairs518F(5′-CCAGCAGCCGCGGTAATACG-3′)and800R(5′-TACCAGGGTATCTAATCC-3′).
Theobtainedsequencesweresub-jectedtothenucleotideBLASTsearchesintheNCBIGenBankdatabasetogetthepercentagesimilarityofthebacterialisolatestothemostrelatedbacterialspecies(Altschuletal.
1990).
Table3PercentsimilarityofthebacterialisolateswiththeircloselyrelatedspeciesbasedontheBLASTsearchesinNCBIGanBankdatabase(Altschuletal.
1990)IsolateBacterialspeciesGenBankaccessionnumberQuerycover(%)Similarity(%)VITEK2(%)Cp1Pseudomonassp.
PDD-59b-7Pseudomonassp.
R3ScM3P1C11PseudomonassyringaestrainPDD-48b-5KR922145KF147001KR922059969696999999Lysinibacillussphaericus/Lysinibacillusfusiformis(91)Cp2CorynebacteriumvariabilestrainC3-13Corynebacteriumsp.
ZT10-3CorynebacteriumvariabileDSM44702KP114214KT597082NR102874969696969696Gardnerellavaginalis(93)Cp3Pseudomonassp.
MN11-3PseudomonasmatsuisolistrainCC-MHH0089PseudomonasmatsuisolistrainCC-MHH0089JQ396614NR134793KJ720680979797969696Dermacoccusnishinomiyaensis/Kytococcussedentarius(96)Cp4BacteriumBELC12Bacillussp.
13K7a2Bacillussp.
7Kp1aBacilluspumilusstrainOU101KT382407KT825840KT825839KR1403779696969699999999Lysinibacillussphaericus/Lysinibacillusfusiformis(87)Cp5PseudomonasmatsuisolistrainCC-MHH0089PseudomonasmatsuisolistrainCC-MHH0089Pseudomonassp.
RBSB9_C3NR134793KJ720680KT390731979796969697Aeromonassalmonicida(98)Cp6Pectobacteriumcarotovorumsubsp.
brasiliensestrainY4516SPectobacteriumcarotovorumsubsp.
brasiliensestrainY3416SPectobacteriumcarotovorumsubsp.
brasiliensestrainY3316SKP187510KP187504KP187503989898979797UnidentifiedorganismCp7Bacillussp.
BG2-9BacilluspumilusstrainT246BacilluspumilusstrainML353KP992115KC764989KC692160989999979797Bacilluspumilus(85)Cp8Paenibacillussp.
MOLA507BacteriumUKRA17Paenibacillussp.
S8AM990732KT382376KR051059969696979797Paenibacilluspolymyxa(90)Cp9Bacillussp.
210_50Bacillussp.
strainRHH15Bacillussp.
L11(2010)GQ199752HQ143613HQ222333999999969696Bacilluspumilus(86)Cp10BacilluspumilusstrainX22BacilluspumilusstrainHN-30BacilluspumilusstrainHN-10FJ763645KT003271KT003256999999989898Bacilluspumilus(88)Cp11BacillusmegateriumstrainD5BacillusmegateriumstrainBCRh8BacillusmegateriumstrainBS9KC441754KT153604KR063189999999989898Bacillusmegaterium(87)Cp12Bacillussp.
C26(2014)BacillussubtilisstrainL-13BacillustequilensisstrainYJ-S4KM117217HQ232422KF876849999999999999UnidentifiedorganismCp13Pseudomonassp.
BE07Unculturedbacteriumisolate1112863845131Pseudomonassp.
DR11(2011)AY456700HQ121073JN210571979797969696Aeromonassalmonicida(97)Cp14BacilluspumilusstrainZA13BacilluspumilusstrainLX11BacilluspumilusstrainY13FJ263042KP192031KF641806989898999999UnidentifiedorganismCp15BacillussubtilisstrainL-13BacillussubtilisstrainYA4BZBacillussp.
RKZ11262HQ232422JQ346075EU835569989898999999UnidentifiedorganismCp16TerribacillussaccharophilusstrainMER_108TerribacillussaccharophilusstrainJP44SK46TerribacillussaccharophilusstrainWA2-4KT719683JX155763JF496471969696969696UnidentifiedorganismSevimetal.
EgyptianJournalofBiologicalPestControl(2018)28:30Page4of9PhylogeneticanalysisPhylogeneticanalysisofthebacterialisolatesandtheircloselyrelatedspecieswasperformedformolecularcharacterizationofthebacterialisolates.
ThesequenceswereeditedusingBioedit,andmultiplesequencealign-mentswerecreatedbyusing16SrRNAsequencesbe-longingtoourstrainsanddifferentbacterialspeciesfromtheNCBIGenBankdatabaseforthepurposeofdevelopingaphylogenetictree.
ThemultiplesequencealignmentwasperformedwithClustalWinBioedit(Hall1999).
Finally,thesequencesweresubjectedtoneighbor-joininganalysiswithp-distancecorrection,gapomission,and1.
000bootstrappseudoreplicatesusingMEGA6.
0(Tamuraetal.
2013).
NucleotidesequenceaccessionnumbersTheGenBankaccessionnumbersofthe16SrRNAgenesequencesbelongingtothebacterialisolatesfromthisstudyarelistedinTable4.
BioassayEachbacteriumisolatedfromC.
pomonellainthestockculturewasstreakedonnutrientagartoobtainasinglecolonyandtocheckthepurityofthecultures.
Afterthat,3mlofnutrientbrothwasinoculatedfromeachsinglecolonyof16bacteriaandincubatedat30°Cover-night.
Attheendoftheincubationperiod,thebacterialdensitywasmeasuredat600nmabsorbanceandadjustedto1.
8*109cfu/mlbycentrifugation(4.
000rpmfor15min)andusingsterilephosphatebuffersolution(PBS)(Moaretal.
1995).
Thebacterialsolutionswerefreshlypreparedandusedforbioassay.
Forthebioassayexperiments,healthyC.
pomonellalarvaewereobtainedfromthelaboratorycultureatAhiEvranUniversity,GeneticBioengineeringandMicrobiologyLaboratory.
Healthylarvaewererandomlyselectedandusedforthebioassay.
Twenty-fivegramsoffreshlypre-paredartificialdiet(fordietingredientsandrearingcondi-tions(Fukovaetal.
2005))wasinoculatedwith1mlofthebacterialsuspensionpreparedasdescribedaboveforeachisolate.
Forthecontrolgroup,25goftheartificialdiethad1mlofthesterilePBSadded.
Thecontami-natedartificialdietswereseparatelyplacedintoplasticboxes(20*10*8mm)withventilatedlidstopermitairflow.
Afterthat,tenthirdinstarC.
pomonellalarvaewereplacedintotheboxforeachreplicateandallowedtofeedonthecontaminateddiets.
Atotaloftenlarvaewereusedforeachreplicate,andallexperimentswererepeatedthreetimes.
Finally,theplasticboxeswerein-cubatedat25°Cunder16:8(day:night)lightregime.
After10days,theboxeswerecheckedforlarvalmor-tality,andthenumberofdeadlarvaewasrecorded.
MortalitydatawerecorrectedbasedonAbbott'sfor-mula(Abbott1925).
Todeterminethedifferencesamongtheisolatesandthecontrolgroup,thedataweresubjectedtoANOVAandsubsequentlytotheLSDmultiplecomparisontest.
BeforeperformingANOVA,alldataweretestedforhomogeneityofvarianceusingLevene'sstatistic.
AlltestswereperformedwithSPSS16.
0statisticalsoftware.
ResultsanddiscussionAtotalof16isolatesofbacteriawereobtainedfromthetreatedC.
pomonellalarvae.
Amongtheseisolates,fivewerefromthefirstandsecondlarvalinstars,eightfromthethirdlarvalinstar,andthreefromthefourthandfifthlarvalinstars.
Theisolateswerecharacterizedonthebasisoftheirmorphological,physiological,andmolecularcharacteristics.
Coloniesofallisolatesweresmooth,exceptforCp9andCp11,whichwererough.
Onlyoneisolate(Cp12)hadamucoidcolony.
Threeisolates(Cp3,Cp5,andCp13)hadyellowcolonies,andoneisolate(Cp4)hadapinkcolony.
Theotherisolatesproducedcreamy-coloredcolonies.
Fiveisolates(Cp1,Cp3,Cp5,Cp6,andCp13)weregram-negative,andtheothersweregram-positive.
Allisolateswerebacilli-shaped,exceptforCp2andCp3whichwerecoccusshaped.
Itwasfoundthatsixisolates(Cp7,Cp8,Cp9,Cp10,Cp14,andCp16)formedspores.
Onlytwoisolates(Cp1andCp13)hadcapsules.
Allmorphologicalcharac-teristicsofthebacterialisolatesaregiveninTable1.
Table4TheproposedidentificationresultsofthebacterialisolatesandtheirGenBankaccessionnumbersfor16SrRNAgenesequencesIsolateSpeciesGenBankaccessionnumberCp1Pseudomonassp.
KX094470Cp2Corynebacteriumsp.
KX094471Cp3Pseudomonassp.
KX094472Cp4Bacillussp.
KX094473Cp5Pseudomonassp.
KX094474Cp6PectobacteriumcarotovorumKX094475Cp7Bacillussp.
KX094476Cp8Paenibacillussp.
KX094477Cp9Bacillussp.
KX094478Cp10Bacillussp.
KX094479Cp11B.
megateriumKX094480Cp12Bacillussp.
KX094481Cp13Pseudomonassp.
KX094482Cp14B.
pumilusKX094483Cp15Bacillussp.
KX094484Cp16TerribacillussaccharophilusKX094485Sevimetal.
EgyptianJournalofBiologicalPestControl(2018)28:30Page5of9Allisolateswereabletogrowin3%NaCl,andonlyoneisolate(Cp3)couldnotgrowin4%NaCl.
Growthcharac-teristicsoftheisolatesinotherNaClconcentrationswerevariable,dependingontheisolate.
AllisolateswereabletogrowinthepHrangeof3,4,and5,exceptforCp12.
AllisolateswereabletogrowatpH6and7.
GrowthcharacteristicsoftheisolatesatotherpHswerevariable,dependingontheisolate.
NoneoftheisolatesgrewatpH4and55°C,andgrowthpropertiesoftheisolatesat30,37,45,and50°Cwerevariable,Fig.
1Phylogenetictreederivedfromneighbor-joininganalysisof16SrRNAsequences(1400bp)fromthefloramembersofC.
pomonellaandtheircloselyrelatedspecies.
Bootstrapvaluesbasedon1000replicateswereindicatedabovenodes.
BootstrapvaluesC≥70arelabeled.
C.
pomonellaisolateswereindicatedwithblackcircle.
ThescaleonthebottomofthedendrogramindicatesthedegreeofdissimilaritySevimetal.
EgyptianJournalofBiologicalPestControl(2018)28:30Page6of9dependingontheisolate.
PhysiologicalpropertiesoftheisolatesaregiveninTable2.
TheVITEK2microbialcharacterizationsystemforbiochemicalcharacterizationoftheisolatesandgram-negativeandgram-positivecardswasusedtoidentifythem.
Fiveisolates(Cp6,Cp12,Cp14,Cp15,andCp16)werenotabletobecharacterizedbyVITEK2.
OtheridentificationsaregiveninTable3.
Thebacterialisolateswerealsocharacterizedonthebasisof16SrRNAgenesequencingtoverifythere-cordedconventionalcharacterizationsoftheisolates.
Basedonmolecularcharacterization,thebacterialiso-lateswereidentifiedasPseudomonassp.
(Cp1,3,5,and13),Corynebacteriumsp.
(Cp2),Bacillussp.
(Cp4,7,9,10,12,and15),Pectobacteriumcarotovorum(Cp6),Paenibacillussp.
(Cp8),Bacillusmegaterium(Cp11),Ba-cilluspumilus(Cp14),andTerribacillussaccharophilus(Cp16)(Table4).
Thisidentificationwasalsosupportedbyphylogeneticanalysis(Fig.
1).
Allisolatescauseddifferentmortalityvaluesincom-parisontoeachother(F=15.
43,df=16,pCydiapomonellaL.
(Lepidoptera:Tortricidae).
AfrJBiotechnol5(22):2081–2085EuzebyJP(1997)Listofbacterialnameswithstandinginnomenclature:afolderavailableontheinternet.
IntJSystBacteriol47(2):590–592FedericiBA(2007)Bacteriaasbiologicalcontrolagentsforinsects:economics,engineering,andenvironmentalsafety.
In:VurroM,GresselJ(eds)Novelbiotechnologiesforbiocontrolagentenhancementandmanagement.
Springer,Dordrecht,pp25–51FukovaI,NguyenP,MarecF(2005)Codlingmothcytogenetics:karyotype,chromosomallocationofrDNA,andmoleculardifferentiationofsexchromosomes.
Genome48:1083–1092GlasnerJD,Marquez-VillavicencioM,KimHS,JahnCE,MaB,BiehlBS,RissmanAI,MoleB,YiX,YangCH,DanglJL,GrantSR,PernaNT,CharkowskiAO(2008)Niche-specificityandthevariablefractionofthePectobacteriumpan-genome.
MolPlant-MicrobeInteract21:1549–1560GnanamanickamSS(2006)Plant-associatedbacteria.
Springer,DordrechtHallTA(1999)BioEdit:auser-friendlybiologicalsequencealignmenteditorandanalysisprogramforWindows95/98/NT.
NucleicAcidsSymp41:95–98HoevenRVD,BetrabetG,ForstS(2008)CharacterizationofthegutbacterialcommunityinManducasextaandeffectofantibioticsonbacterialdiversityandnematodereproduction.
FEMSMicrobiolLett286:249–256IanKT,KennethSB,HolevaCM,RJPB(2003)Softroterwiniae:fromgenestogenomes.
MolPlantPathol4(1):17–30KhetanSK(2001)MicrobialPestControl.
MarcelDekker,NewYorkLaceyLA,UnruhTR(2005)Biologicalcontrolofcodlingmoth(Cydiapomonella,Lepidoptera:Tortricidae)anditsroleinintegratedpestmanagement,withemphasisonentomopathogens.
Vedalia12(1):33–60LalS,TabacchioniS(2009)EcologyandbiotechnologicalpotentialofPaenibacilluspolymyxa:aminireview.
IndianJMicrobiol49:2–10LeeCW,LucasS,DesomazeaudMJ(1985)Phenylalanineandtyrosinecatabolisminsomecheesecoryneformbacteria.
FEMSMicrobiolLett26(2):201–205LigozziM,BerniniC,BonoraMG,deFatimaM,ZulianiJ,FontanaR(2002)EvaluationoftheVITEK2systemforidentificationandantimicrobialsusceptibilitytestingofmedicallyrelevantgram-positivecocci.
JClinMicrobiol40(5):1681–1686LipaJJ,WilandE(1972)BacteriaisolatedfromcutwormsandtheirinfectivitytoAgrotissp.
ActaMicrobiolPol4:127–140LiuW,JiangL,GuoC,YangSS(2010)Terribacillusaidingensissp.
nov.
,amoderatelyhalophilicbacterium.
IntJSystEvolMicrobiol60:2940–2945MaB,HibbingME,KimHS,ReedyRM,YedidiaI,BreuerJ,BreuerJ,GlasnerJD,PernaT,KelmanA,CharkowskiAO(2007)HostrangeandmolecularphylogeniesofthesoftrotenterobacterialgeneraPectobacteriumandDickeya.
Phytopathology97(9):1150–1163MaharAN,DarbanDA,GowenSR,HagueNGM,JanND,MunirM,MaharAQ(2005)UseofentomopathogenicbacteriumPseudomonasputida(Enterobacteriaceae)anditssecretionagainstgreaterwaxmoth,Galleriamellonellapupae.
JEntomol2(1):77–85McSpaddenGardenerBB(2004)EcologyofBacillusandPaenibacillusspp.
inagriculturalsystems.
Phytopathology94:1252–1258MenezesCB,Bonugli-SantosRC,MiquelettoPB,PassariniMR,SilvaCH,JustoMR,LealRR,Fantinatti-GarbogginiF,OliveiraVM,BerlinckRG,SetteLD(2010)Microbialdiversityassociatedwithalgae,ascidiansandspongesfromthenorthcoastofSoPaulostate,Brazil.
MicrobiolRes165:466–482MoarWJ,Pusztzai-CareyM,MackTP(1995)ToxicityofpurifiedproteinsandtheHD-1strainfromBacillusthuringiensisagainstlessercornstalkborer(Lepidoptera:Pyralidae).
JEconEntomol88:606–609OwuamaCI(2001)Entomopathogenicsymbioticbacteria,XenorhabdusandPhororhabdusofnematodes.
WorldJMicrobiolBiotechnol17:505–515PajacI,BaricB,MikacK,PejicI(2012)NewinsightsintothebiologyandecologyofCydiapomonellafromappleorchardsinCroatia.
BullInsectol65(2):185–193RajagopalR(2009)Beneficialinteractionsbetweeninsectsandgutbacteria.
IndianJMicrobiol49:114–119RoopaHK,RebijithKB,AsokanR,MahmoodR,KumarK(2014)Isolationandidentificationofculturablebacteriafromhoneydewofwhitefly,Bemisiatabaci(G.
)(Hemiptera:Aleyrodidae).
MetaGene2:114–122RuiuL(2015)Insectpathogenicbacteriainintegratedpestmanagement.
Insects6:352–367RuiuL,SattaA,FlorisI(2013)Emergingentomopathogenicbacteriaforinsectpestmanagement.
BullInsectol66(2):181–186SevimE,elebi,SevimA(2012)DeterminationofthebacterialfloraasamicrobialcontrolagentofToxopteraaurantii(Homoptera:Aphididae).
Biologia67(2):397–404SoutourinaOA,SemenovaEA,ParfenovaVV,DanchinA,BertinP(2001)ControlofbacterialmotilitybyenvironmentalfactorsinpolarlyflagellatedandperitrichousbacteriaisolatedfromLakeBaikal.
ApplEnvironMicrobiol67:3852–3859TamuraK,StecherG,PetersonD,FilipskiA,KumarS(2013)MEGA6:molecularevolutionarygeneticsanalysisversion6.
0.
MolBiolEvol30:2725–2729VicenteCSL,NascimentoFX,EspadaM,BarbosaP,HasegawaK,MotaM,OliveiraS(2013)CharacterizationofbacterialcommunitiesassociatedwiththepinesawyerbeetleMonochamusgalloprovincialis,theinsectvectorofthepinewoodnematodeBursaphelenchusxylophilus.
FEMSMicrobiolLett347:130–139YamadaK,KinoshitaS,TsunodaT,AidaK(1972)Themicrobialproductionofaminoacids.
Wiley,NewYorkYunJH,RohSW,WhonTW,JungMJ,KimMS,ParkDS,YoonC,NamYD,KimYJ,ChoiJH,KimJY,ShinNR,KimSH,LeeWJ,BaeJW(2014)Insectgutbacterialdiversitydeterminedbyenvironmentalhabitat,diet,developmentalstage,andphylogenyofhost.
ApplEnvironMicrobiol80(17):5254–5264Sevimetal.
EgyptianJournalofBiologicalPestControl(2018)28:30Page9of9
- microbialcydia相关文档
- 值得解决ios 5.0.1越狱后cydia添加源和搜索软件闪退的问题
- 选择ios 5.1.1 cydia闪退解决方法
- 点击5.01版cydia闪退补丁解决办法
- 越狱Cydia闪退修复教程
- 点击解决CYDIA闪退
- 文件系统Cydia闪退用如下方法解决
Vultr再次发布充值多少送多少活动
昨天我们很多小伙伴们应该都有看到,包括有隔壁的一些博主们都有发布Vultr商家新的新用户注册福利活动。以前是有赠送100美元有效期30天的,这次改成有效期14天。早年才开始的时候有效期是60天的,这个是商家行为,主要还是吸引到我们后续的充值使用,毕竟他们的体验金赠送,在同类商家中算是比较大方的。昨天活动内容:重新调整Vultr新注册用户赠送100美元奖励金有效期14天今天早上群里的朋友告诉我,两年...
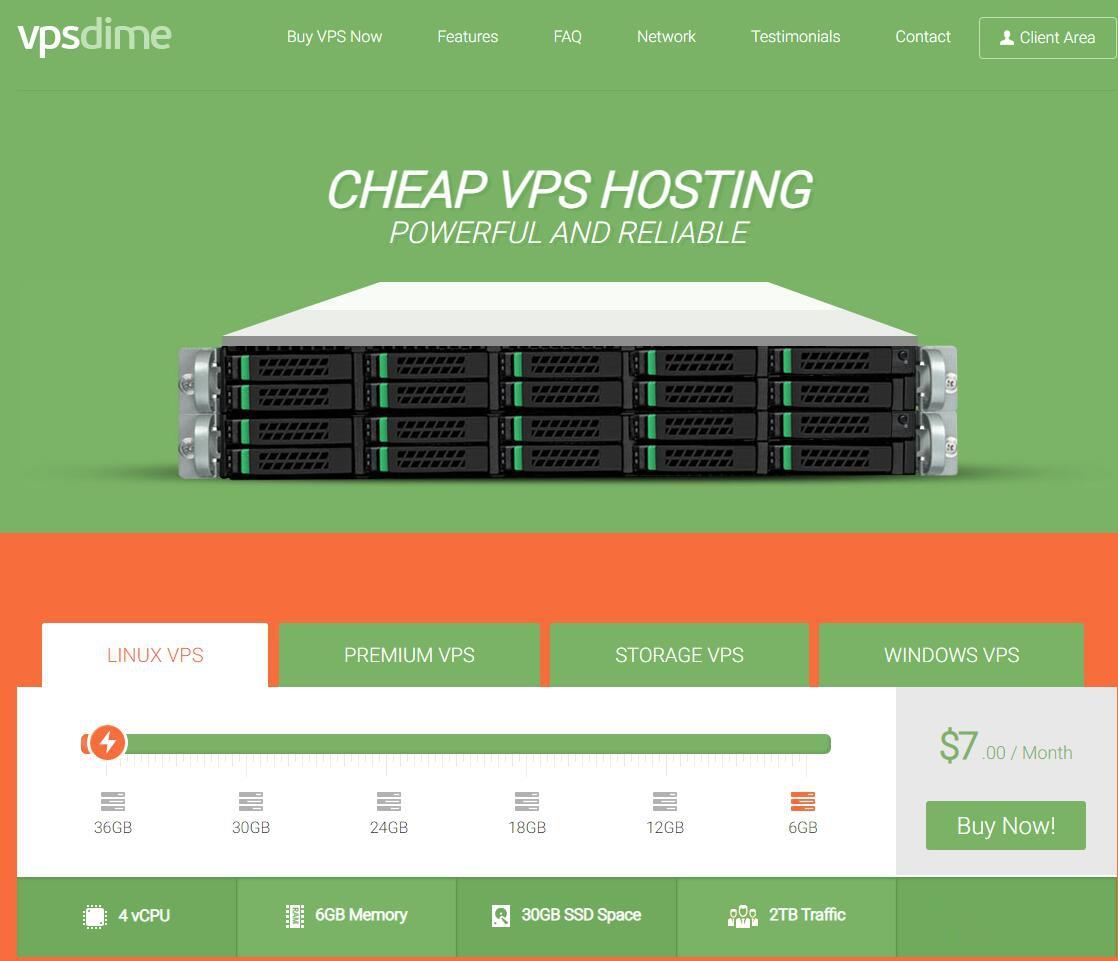
VPSDime7美元/月,美国达拉斯Windows VPS,2核4G/50GB SSD/2TB流量/Hyper-V虚拟化
VPSDime是2013年成立的国外VPS主机商,以大内存闻名业界,主营基于OpenVZ和KVM虚拟化的Linux套餐,大内存、10Gbps大带宽、大硬盘,有美国西雅图、达拉斯、新泽西、英国、荷兰机房可选。在上个月搞了一款达拉斯Linux系统VPS促销,详情查看:VPSDime夏季促销:美国达拉斯VPS/2G内存/2核/20gSSD/1T流量/$20/年,此次推出一款Windows VPS,依然是...
hostkvm:7折优惠-香港VPS韩国VPS,8折优惠-日本软银、美国CN2 GIA、新加坡直连VPS
hostkvm本月对香港国际线路的VPS、韩国CN2+bgp线路的VPS正在做7折终身优惠,对日本软银线路、美国CN2 GIA线路、新加坡直连线路的VPS进行8折终身优惠促销。所有VPS从4G内存开始支持Windows系统,当然主流Linux发行版是绝对不会缺席的!官方网站:https://hostkvm.com香港国际线路、韩国,7折优惠码:2021summer日本、美国、新加坡,8折优惠码:2...

cydia 闪退为你推荐
-
邮箱群发163的邮箱如何群发万维读者网读者投稿邮箱万维读者网《读者》要订购有网站吗?办公协同软件免费的多人协同办公软件哪些,我了解的有钉钉、企业微信,其他的还有么?网易公开课怎么下载哪位高手指导一下,如何下载网易公开课啊?srv记录SRV记录的定义服务器连接异常手机服务器连接异常微信电话本怎么用微信电话本在哪里 微信电话本怎么打开网站排名靠前如何使网站排名靠前?去鼠标加速度win7怎么去鼠标加速度