www.proteomics-journal.com
6kkbb.com 时间:2021-04-08 阅读:()
Page1ProteomicsReceived:28-Apr-2014;Revised:14-Aug-2014;Accepted:06-Oct-2014Thisarticlehasbeenacceptedforpublicationandundergonefullpeerreviewbuthasnotbeenthroughthecopyediting,typesetting,paginationandproofreadingprocess,whichmayleadtodifferencesbetweenthisversionandtheVersionofRecord.
Pleasecitethisarticleasdoi:10.
1002/pmic.
201400169.
Thisarticleisprotectedbycopyright.
Allrightsreserved.
InvestigationofPhosphoproteomeinRAGEsignalingKedarBBatkulwar,SnehaBBansode,GouriVPatil,RashmiKGodbole,RubinaSKazi,SubashchandraboseChinnathambi,DhanasekaranShanmugam,MaheshJKulkarni*ProteomicsFacility,DivisionofBiochemicalSciences,CSIR-NationalChemicalLaboratory,Pune-411008,India#Authors2to5haveequallycontributed*ForcorrespondenceMaheshJKulkarniProteomicsFacility,DivisionofBiochemicalSciences,CSIR-NationalChemicalLaboratory,Pune-411008,IndiaE-mail:mj.
kulkarni@ncl.
res.
inPhone:+912025902541www.
proteomics-journal.
comPage2ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
AbstractReceptorforAdvancedGlycationEndproducts(RAGE)isoneofthemostimportantfactorsimplicatedindiabetes,cardiovasculardiseases,neurodegenerativediseases,andcancer.
Itisapatternrecognitionreceptor,byvirtueofitsabilitytointeractwithmultipleligands;RAGEactivatesseveralsignaltransductionpathwaysthroughinvolvementofvariouskinases,whichphosphorylatetheirrespectivesubstrates.
OnlyfewsubstrateshavebeenknowntobephosphorylatedinresponsetoactivationbyRAGE(e.
g.
NF-кB),howeveritispossiblethatthesekinasescanphosphorylatemultiplesubstratesdependingupontheirexpressionandlocalization,leadingtoalteredcellularresponsesindifferentcelltypesandconditions.
Onesuchexample,Glycogensynthasekinase3beta(GSK3β)whichisknowntophosphorylateglycogensynthase,actsdownstreamtoRAGEandhyperphosphorylatesMicrotubuleAssociatedProteinTau(MAPT)causingneuronaldamage.
Thus,itisimportanttounderstandtheroleofvariousRAGEactivatedkinasesandtheirsubstrates.
Therefore,wehavereviewedherethedetailsofRAGEactivatedkinasesinresponsetodifferentligandsandtheirrespectivephosphoproteome.
Further,wediscusstheanalysisofthedataminedforknownsubstratesofthesekinasesfromPhosphoSitePlus(http://www.
phosphosite.
org)database,andtheroleofsomeoftheimportantsubstratesinvolvedincancer,diabetes,cardiovasculardiseasesandneurodegenerativediseases.
Insummary,thisreviewprovidesinformationonRAGEactivatedkinasesandtheirphosphoproteome,whichwillbehelpfulinunderstandingthepossibleroleofRAGEanditsligandsinprogressionofdiseases.
Keywords:disease,glycation,kinase,phosphoproteome,RAGEwww.
proteomics-journal.
comPage3ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Abbreviations1.
RAGE-Receptorforadvancedglycationendproducts2.
AGEs-Advancedglycationendproducts3.
NF-кB-NuclearfactorkappaB4.
GSK3β-Glycogensynthasekinase-3beta5.
HMGB1-Highmobilitygroupboxprotein16.
Aβ-Amyloidbeta7.
SP1-Specificityprotein18.
MAPK-Mitogenactivatedproteinkinase9.
PKC-ProteinkinaseC10.
JNK-c-JunN-terminalKinase11.
JAK-Janusactivatedkinase12.
ERK-Extracellularsignalregulatedkinase13.
iNOS-induciblenitricoxidesynthase14.
eNOS-Endothelialnitricoxidesynthase15.
mTOR-Mammaliantargetofrapamycin16.
MMP-Matrixmetalloproteinase17.
STAT-Signaltransducersandactivatorsoftranscription18.
TNFα-Tumornecrosisfactoralphawww.
proteomics-journal.
comPage4ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
IntroductionRAGE,amulti-ligandpatternrecognitionreceptor,belongstotheimmunoglobulinsuperfamilyofproteins.
Althoughitwasfirstdescribedasareceptorforadvancedglycationendproducts(AGEs),itinteractswithdiverseclassesofligandincludingS100B,highmobilitygroupboxprotein(HMGB)-1,Mac-1,amyloidbeta(Aβ)peptideandbetasheetfibrils[1].
TheRAGEmRNAandmatureproteincanundergoalternativesplicingandproteolysis,respectivelyresultinginmanyvariants.
FulllengthRAGE(flRAGE)consistsof5domains,threeextracellular,asingletransmembranedomainandasmallcytosolictail.
ExtracellulardomainsarelabeledasV,C1andC2domain[2].
WhilethesolubleRAGE(sRAGE)lackstransmembraneandcytosolicdomain,andthereforeitisneitheranchoredincellmembranenorabletoactivatesignalingpathwayslikeflRAGE(Fig1)[3].
ThesRAGEinteractswithRAGEligandsandhelpsintheclearanceofthesefromthesystem[4].
TheinitiationofRAGEsignalingcommonlyoccursthroughproductionofROSviaNADPHoxidaseandactivationofvariouskinasesincludingMAPKlikeERK1/2,SAPK/JNK,p38;PI3K/Akt;JAK/STAT;GSK3βetc[5].
ThesekinasestriggerseveraldownstreamsignalingcascadesoftenviaactivationoftranscriptionfactorsNF-кBandSP1,therebymodulatingspecificcellularprocesses[6].
Becauseofitsroleinoxidativestressandinflammation,RAGEisimplicatedinseveraldiseases.
Indiabetes,AGEsarethepredominantligandsthatinteractwithRAGEandcontributetothepathogenesisofdiabeticcomplicationslikeglomerulopathy,atherosclerosis,vasculopathyetc[7].
AGEsareagroupofheterogeneousmoleculesformedasaresultofaseriesofnon-enzymaticreactionsbetweenreducingsugarsandproteins[8,9].
SomeofthepredominantAGEsincludecarboxymethyllysine(CML),carboxyethyllysinewww.
proteomics-journal.
comPage5ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
(CEL),pentosidineetc[10-12].
AGEformationisacceleratedindiabetesduetohyperglycemiccondition[13].
AccumulationofAGEsleadsto(i)alteredproteinstructureandfunction[14],(ii)proteinaggregation,fibrilformation,andproteaseresistance[15,16],(iii)dysfunctionofextracellularmatrix[14]and(iv)RAGEactivationanddownstreamsignaling[17].
AGE/RAGEinteractioncontributessubstantiallytotheprogressionofdiabeticcomplications,cardiovasculardiseases,acceleratedaging,neurodegenerativediseases,andcancer[18].
WhereasinAlzheimer'sdisease,AβistheprincipalligandthatinteractswithRAGE[19].
Itconsistsof36-43aminoacidresiduesproducedasaresultofsequentialcleavageofAPPbybetaandgammasecretases[20].
RAGEmediatedAβinternalizationresultsinmitochondrialdysfunctioncausingcellulardamage[21].
Whileincancer,S100B,acalciumbindingproteinisknowntobeinvolvedinRAGEactivationbydimerization[22].
S100B/RAGEinteractionelicitsvariouscellulareffectsincludingcellsurvivalandcellproliferation[23,24].
HMGB1alsocalledasHMG1oramphoterinisasmallnon-histonechromatinproteinwhichinteractswithRAGEandactivatesproinflammatorypathways[25,26].
BlockadeofRAGEmediatedsignalingsuppressestumorgrowthandmetastasis,suggestingitspotentialinvolvementincancer[27].
Hence,RAGEplaysanimportantroleintheregulationofvitalcellprocessessuchasapoptosis,autophagy,inflammationandproliferation.
TheinvolvementofRAGEindifferentdiseaseshasbeenreviewedingreatdetailearlier[28-31].
However,thedetailedmolecularmechanismofRAGEsignaling,especiallytheinvolvementofvariouskinasesandtheirsubstratesisnotwellunderstood.
MostofthereportedstudiesonRAGEsignalingfocusonactivationofspecifickinaseslikeMAPKs,PI3K,PKCandGSK3β.
ThephosphorylationofonlyfewimportantsubstratessuchasNF-кB,FOXO,andSP1isknown;butglobalphosphoproteomeinRAGEactivatedsignalingispoorlystudied.
Itisimportanttoidentifywww.
proteomics-journal.
comPage6ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
thefactorsthatfacilitatetherecruitmentofspecifickinases,alongwiththeirphosphoproteome,whichcangreatlyenhanceourunderstandingofthecellularresponsestoRAGEsignaling.
Therefore,inthisreviewwehavediscussedindetailaboutRAGEactivatedkinasesandtheirphosphoproteomeinresponsetodifferentligands.
RAGEactivatedkinasesOnceactivatedbyitsligands,RAGEcanrecruitseveralkinasesandtriggeracascadeofsignalingevents(Fig2).
SomeoftheimportantkinasesthatareactivateddownstreamtoRAGEaredescribedbelow.
Mitogenactivatedproteinkinases(MAPK)MAPKsareserine/threonineproteinkinasesthattranslateexternalstimuliintoawiderangeofcellularresponses.
ConventionalMAPKsincludeextracellularsignal-regulatedkinases1/2(ERK1/2),c-JunN-terminalkinases1/2/3(JNK1/2/3),p38isoforms(α,β,γ,andδ),andERK5.
AtypicalMAPKsincludeERK3/4,ERK7,andNemo-likekinase(NLK).
EachgroupofconventionalMAPKsiscomposedofasetofthreeevolutionarilyconserved,sequentiallyactingkinases:aMAPK,aMAPKkinase(MAPKK),andaMAPKKkinase(MAPKKK)[32].
SeveralligandsinteractwithRAGEandactivatedifferentMAPKsdependingoncelltypeanddisease.
p38MAPKisoneofthekeydownstreameffectorofRAGEthatmediatesactivationofNF-кBdependentsecretionofcytokinesresultinginacceleratedinflammation.
AGEinducedNF-кBactivationresultsinincreasedsecretionofTumorNecrosisFactoralpha(TNF-α),interleukin-1beta(IL-1β),andMonocytesChemoattractantProtein(MCP)-1.
RAGEhasbeenimplicatedintheformationofproinflammatorymilieu,whichplaysanimportantroleintheadvancementofseveraldisorders[33].
www.
proteomics-journal.
comPage7ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Indiabetes,AGEinducedp38MAPKactivationresultsinincreasedalkalinephosphataseactivityviaRAGE,whichin-turninducesosteoblastlikedifferentiationofsmoothmusclecellsresultinginvascularcalcification[34].
Indiabeticnephropathy(DN),AGE/RAGEinteractionactivatesp38MAPKthatphosphorylatesATF2,atranscriptionfactorwhichinducesiNOSexpressioninmesangialcells[35].
Inpodocytes,thisinteractionresultsinactivationoftranscriptionfactorFOXO4viap38MAPK,leadingtoapoptosis[36].
AGEscaninduceexpressionofconnectivetissuegrowthfactor(CTGF),whichpromotestheadvancementofdiabeticnephropathy.
TheexpressionofCTGFdependsonRAGE-ERK/p38-Smad3cross-talkpathway[37].
AGEsalsoactivateERK1/2andEGFRviaRAGEmediatedROSinNRF-49Fkidneycells,whichareimplicatedindiabeticnephropathy[38].
AGEsactivateSmad2/3inhumanvasculopathyviaRAGEandERK/p38MAPKs[39].
AGEdepositionandRAGEexpressioncontributestotheprogressionofatherosclerosisviaERKactivationindiabeticandhypertensivemice[40].
OligomerizationofRAGEisbelievedtoplayanessentialroleinsignaltransduction.
HeparansulfateinduceshexamerizationoftheextracellularVandC1domainspresentinRAGE,resultinginactivationofERK1/2inendothelialcells[41].
S100A4demonstratedRAGEmediatedERKphoshorylationandMMP2activation,whichincreasescellmotilityinhumanpulmonaryarterysmoothmusclecells(hPASMCs)contributingtoprogressionofvasculardiseases[42].
AGEshavebeenalsoimplicatedindiabetickeratopathy,whereintheyinduceapoptosisincornealepithelialcellsbyROSgenerationthroughactivationofNADPHoxidase,JNKandp38MAPKinRAGEdependentmanner[43].
Incancer,bindingofamphoterintoRAGEenhancestheRAGEboundERK1/2activity,asdemonstratedbyphosphorylationofmyelinbasicprotein,anERKsubstrate[44].
Inwww.
proteomics-journal.
comPage8ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
myoblastsandrhabdomyosarcomacells,HMGB1/RAGEinteractionpreventstumorformationwhereasingliomasitfacilitatesmetastasisensuingininvasivephenotype.
RAGEactivationbyHMGB1leadstomyogenesisviaCdc42-Rac-MKK6-p38MAPKpathwayandisassociatedwithinhibitionofproliferation.
FunctionalinactivationofRAGEcausesdecreaseinactivityofp21Waf1,aproliferationinhibitorandincreaseinactivityofcyclinD1,retinoblastomaprotein,ERK1/2andJNK[45,46].
WhereasinC6gliomacells,RAGE/amphoterininteractionplaysanimportantroleintumorproliferation,invasionandmatrixmetalloproteinase(MMP)expression,whileblockingthisinteractiondecreasesgrowthandmetastasisbyinactivationofp44/p42,p38andJNKMAPKpathway[47].
S100A8/A9ligandhasadualroleintumorbiology;stimulatespseudopodiaformationandexertsgrowthpromotingactivity.
ItpromotescellgrowthinaRAGEdependentmannerbyphosphorylationofp38andp44/42MAPKsandactivationofNF-кB[48].
p38MAPKcausesactivationofMAPKAPkinase-2(MK2),Hsp27andSAPK/JNKpathways.
SimilarresponseswereobservedinHaCaTkeratinocytesandclearcellrenalcellcarcinoma(CCRCC)viaERK1/2activationinRAGEdependentmanner[49].
HMGB1/RAGEinteractionplaysanimportantroleintheadvancementofliverfibrosis.
RAGEmediatedHMGB1internalizationcausesSmad2andERK1/2phosphorylationresultingincollagentypeIandα-smoothmuscleactin(α-SMA)expression,andproliferationofhepaticstellatecellsleadingtothefibroticchangesofliver[50].
S100P,aRAGEligandthatisexpressedathighlevelsincoloncancer,interactswithRAGEandstimulatesgrowth,migration,ERKphosphorylation,andNF-кBactivationincancercells[51].
VEGFincardiacmyocytesrespondtoS100BandRAGEinteractioninducedERK1/2phosphorylationandmyofibroblastproliferationcontributingtomyocardialinfarction[52].
RAGE/S100BinteractionaftermyocardialinfarctionstimulatesmyocyteapoptosisbyactivatingERK1/2andp53signaling[53].
www.
proteomics-journal.
comPage9ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Incorticalneurons,RAGEmediatesAβuptakeacrossthecellmembraneviaactivationofp38MAPK.
p38MAPKactivationcausesinflammatoryresponseandisimplicatedinthepathogenesisofAlzheimer'sdisease[54].
Aβimpairslong-termpotentiation(LTP)intheentorhinalcortexthroughRAGE-mediatedp38MAPKactivation[55].
Bloodbrainbarrier(BBB)ismainlycomposedofcerebralendothelialcells(CEC)andastrocytes.
BBBdamageisassociatedwithseveraldisordersincludingAlzheimer'sdisease.
AβinducedERK1/2andcytosolicphosphorilaseA2phosphorylationinCECsandastrocytesoccursviaRAGEdependentpathwayscausingcellulardamage[56].
APP/PS1transgenicalzheimermousemodeldemonstratesstrongincreaseintheexpressionofmouseformylpeptidereceptor(mFPR1/2)andRAGEexpressioninthecortexandhippocampus.
FPRandRAGEplayanimportantroleinAβ,S100BandAGE-BSAinducedERK1/2phosphorylationandregulationofcAMPlevelinglialcells[57].
AβstimulatedtransientactivationofERK1/2,p38MAPKinmicrogliaandmonocytesresultsinphosphorylationofcAMPresponseelement-binding(CREB)protein,providingamechanismforAβinducedchangesingeneexpression[58].
Inaddition,RAGEviaMAPKisalsoinvolvedinasthma,chronicobstructivepulmonarydisease,cysticfibrosis,andrheumatoidarthritis.
S100A12/RAGEinteractioninducesoverexpressionofMUC5AC,amajormucinprotein,viaERKactivationinairwayepithelialcells[59].
RAGEinducesdamageinhumanpulmonaryarteryendothelialcellsviap38MAPKactivationandHsp27phosphorylationresultinginacutelunginjury[60].
HMGB1inpresenceofLPSupregulatesTLR4andRAGEexpressioninSynovialfibroblasts,whichinducesp38MAPKandNF-кBactivationresultingintheproductionofIL-6,MMP3,andMMP13inrheumatoidarthritis[61,62].
www.
proteomics-journal.
comPage10ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
c-JunN-terminalkinase(JNK)JNKisaprolinedirectedSerine/Threoninekinase,andbelongstotheMAPKfamilywhichintegrateavarietyofenvironmentalstimulitocauseproliferation,differentiation,development,inflammatoryresponseandapoptosis.
TheJNKarmofMAPKsisactivatedinconditionsofintracellularandenvironmentalstress;henceitisalternativelyreferredtoasStressActivatedProteinKinase(SAPK).
JNKisactivatedbydualphosphorylationataspecificthreonineandtyrosineresiduesbyMKK4andMKK7kinases(MAPKKs)[63].
JNKactivationcausesphosphorylationoftranscriptionfactorsaswellasotherproteinstoeitheractivateorinactivatethem.
Itpredominantlyphosphorylatesandactivatesthec-JunandATF-2transcriptionfactorsoftheAP1complex.
JNKisactivatedbyERstress,osmoticstress,redoxstress,UVradiation,thecytokinesTNF-αandIL-1β,microbialinvasion,aswellasbyRAGEactivationinmanypathologicalconditions[64].
Inosteoarthritis,accumulationofAGEsinthearticularcartilagechondrocyteshasbeenassociatedwithinflammation.
AGEinducesexpressionofCOX-2andmicrosomalprostaglandinEsynthase-1byRAGEmediatedactivationofJNK,whichleadstoincreasedproductionofProstaglandinE2,amediatorofinflammation[65].
AGEBSAinducessecretionofMMP1,3and13inhumanosteoarthriticchondrocytesbyinteractionwithRAGEandactivationofJNKandp38kinase[66].
Onthecontrary,AGE/RAGEsignalingintheinfiltratingmacrophagescausesphosphorylationofp44/42ERK,p38,JNK,andPI3KandinhibitionofJNKactivitysignificantlyenhancedthestimulatoryeffectofAGEonMMP9secretion[67].
InDiabeticcondition,osteoblastapoptosisanddeficientbonemarrowformationismediatedbyAGE/RAGEinteraction.
CMLmodifiedcollagenactivatesp38,JNK,andtranscriptionfactor,FOXO1,leadingtoincreasedcaspase-3activityinfibroblastsresultinginwww.
proteomics-journal.
comPage11ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
apoptosis,whileinhibitionofthesekinasesreducedCML-collagenstimulatedapoptosis[68].
InMyocardialinfarction,stroke,andvenousthromboembolism,AGE/RAGEinteractioncausestranslocationofPKC-β2thatphosphorylatesJNKleadingtoincreasedtranscriptionofearlygrowthresponse-1(EGR-1).
EGR-1playsanintegralroleinthebiologicalresponsetohypoxia,itsupregulationmediatesincreasedexpressionofinflammatoryandprothromboticgenesresultingincellulardamage[69].
RAGEactivationbyAGEs,S100andHMGB1hasbeenimplicatedinischemicrepurfusion(I/R)injuryofcardiomyocytes.
CML/RAGEinteractionduringI/RincardiomyocytesandmousemodelcausesactivationofJNKandSTAT3leadingtocellulardamage[70,71].
LigationoftheheterodimericS100A8/A9proteinwithRAGEinpostischemiccardiacfibroblastsincreasedNF-кBnucleartranslocationandexpressionofpro-inflammatorycytokinesbyactivationofallthethreearmsofMAPKsnamelyERK,JNKandp38[72].
WhereasinteractionofHMGB1withRAGEinI/RcausedtheactivationofERK1/2andJNKMAPKsonly,alongwithincreasednucleartranslocationofNF-кB[73].
SimilareffectsofHMGB1/RAGEinteractionwereobservedinadiabeticmousemodelexposedtoI/R,thereforeimplyingaroleofHMGB1inheartfailureasacomplicationofdiabetes[74].
InconditionsofhepaticI/R,apartfromactivationofthesekinases,RAGEsignalingalsoactivatesc-Jun,leadingtoincreasedserumalanineaminotransferaseactivity,TNF-αandIL-6levels,NF-кBactivityandultimatelyapoptosisinlivertissue[75].
HMGB1functionsasacytokineandactivatesmacrophagesbybindingtoRAGE.
ThisinteractioncausesincreasedexpressionofMHCclassII,TNF-αandNObyactivationofp38,p42/44,JNKMAPKsalongwithincreasednucleartranslocationofNF-кB[76].
JNKactivationviaRAGEisimplicatedinpathogenesisofatherosclerosis.
S100B,andoxidizedLDLs(oxLDL)containingAGEepitopesinteractwithRAGEinaorticendothelialcellstocauseactivationofJNKleadingtoincreasedMMP2activityandupregulationofVCAM-1www.
proteomics-journal.
comPage12ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
whichmarksthesitesdestinedfordevelopmentofatheroscleroticplaques[77].
AGE/RAGEinteractionisreportedtocauseactivationofJNK,whichfurtherphosphorylatesAP1transcriptionfactortocauseexpressionofE-selectin,whichmediatesatherosclerosis[78].
Inendothelialcells,HMGB1stimulatesproinflammatoryresponsebyupregulationofleukocyteadhesionmoleculesnamelyICAM-1,VCAM-1,TNFα,IL-8,MCP-1,plasminogenactivatorinhibitor1(PAI-1),andtissueplasminogenactivator(tPA).
ThiseffectwasobservedbytransientphosphorylationoftheERK,p38andJNKandincreasednucleartranslocationofNF-кB[79].
S100B/RAGEInteractioninBV-2microglialcellsshowedupregulationofCOX2,IL-1βandTNF-αexpressionandchemokinesecretionbyparallelactivationofCdc42-Rac1-JNK-AP1andRas-Rac1-NF-кBpathwayscausinginflammation[80-82].
S100B/RAGEinteractioninamixedpopulationofneuronalandglialcellscausedactivationofJNK,whichspecificallyphosphorylatedthec-Jun.
c-JuncausestheexpressionofDickopff-1whichdisruptsWntsignalingpathwayandactivatesGSK3βresultinginTauhyperphosphorylation,apathologicalhallmarkofAD[83].
AβcausesapoptosisthroughinteractionwithRAGEviaactivationofp38,JNKpathways,andNF-кB,thusimplyingtheroleofRAGEinneurodegenerationincybridsofsporadicAD[84].
Incaseofbraininjury,HMGB1releasedfromreactiveastrocytescausesproliferationofneuronalstemandprogenitorcellsbyinteractionwithRAGEanddownstreamJNKphosphorylation[85].
ProteinkinaseC(PKC)ProteinkinaseC,amultifunctionalprotein,belongstoafamilyofserinethreoninekinase.
Thereare11isoformsofPKCandthesecanbeclassifiedintothreesubgroups;1)conventionalPKCs(α,β1,β2,andγ)thatareCa2+dependentandactivatedbywww.
proteomics-journal.
comPage13ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
phosphatidylserineandthesecondmessengerdiacylglycerol(DAG);2)novelPKCs(δ,η,ε,β)thatareCa2+independentandstimulatedbyDAG;and3)atypicalPKCs(ζandι/λ),neitherofwhichareknowntorequirecalciumorDAGforactivation.
AllPKCsappeartobephosphorylatedbyphosphoinositide-dependentproteinkinase1(PDPK1)[86].
PKChasbeenassociatedwithvascularalterationssuchasincreasedpermeability,contractility,extracellularmatrixsynthesis,cellgrowth,apoptosis,angiogenesis,leukocyteadhesionandcytokineactivation[87].
PKCphosphorylatescytoplasmicdomainofRAGEuponbindingofligands.
TIRAPandMyD88functionasadaptorproteinsforRAGEwhichfurtherrecruitIRAK4,whichinturnactivatesdownstreamkinasessuchasAkt,p38,JNKandIKKs.
Thisultimatelyleadstoproductionofinflammatorycytokines,IL-6,IL-8,andTNFαthroughactivationofNF-кB,whichregulatesinflammation,immuneresponseandothercellularfunctions[88].
PKCactivationinRAGEsignalingplaysanimportantroleintheprogressionofdiseasessuchasdiabetesandAlzheimer's.
ImpairmentofinsulinactioninthemuscleismediatedthroughtheformationofamultimolecularcomplexincludingAGE/RAGE/IRS-1/SrcandPKCα.
ThiscomplexrequiresIRS-1,oneofthekeymoleculesininsulinsignaling.
PKCmediatestheformationofthiscomplex,whichinhibitinsulinactioninskeletalmusclecellsinvitroand,possibly,invivo[89].
AGE/RAGEinteractioncausesPKCmediatedNADPHoxidaseactivationthatleadstoincreasedretinalcapillarypermeability,whichisassociatedwithinitialstagesofmacularedemaanddiabeticretinopathy[90].
AGEassociatedRAGEexpressionisfoundtobesignificantlyincreasedincaseofdiabeticnephropathy.
NADPHoxidase,whichplaysanimportantroleinROSproduction,isactivatedbyAGE/RAGEinteraction.
ThesubunitsofNADPHoxidaseincludingp47phox,nox4andp22phoxarewww.
proteomics-journal.
comPage14ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
activatedbyPKCαmediatedphosphorylation,whichfurtherresultsinoxidativestress[91].
Conversely,AGE/RAGEmediatedROSproductionactivatesPKC,whichphosphorylatesp47phoxthatinturnactivatesNADPHoxidase,resultinginapositivefeedbackloopforROSproduction[92].
AGE/RAGEinteractionelevatesCx43expression,themajorconnexininventricularcardiomyocytesthroughactivationofPKCandERKpathway.
ItalsoenhancesCx43redistributioninvivo,whichmaycontributetothearrhythmiasindiabetes[93].
Indiabeticmononuclearphagocytes,RAGEactivatesPKCwhichphosphorylatespleckstrin,acriticalmoleculeforproinflammatorycytokinesecretion[94].
AGEscanaugmentinflammatoryresponsesbyup-regulatingCOX-2viaRAGEandPKC,ERKandp38MAPKsignalingpathways,therebyleadingtomonocyteactivationandvascularcelldysfunction[95].
Inneuronalcells,AGE/RAGEinteractioncausesapoptosisduetoactivationofNADPHoxidase,PKCdeltaandredoxsensitivetranscriptionfactorAP1[66].
RAGEplaysacrucialroleinactivationofepithelialsodiumchannel(ENaC)throughPKC-gp91phoxsignaling,whichregulateslungfluidbalance[96].
Phosphoinositide3kinases/Akt(PI3K/Akt)Phosphoinositide3kinases(PI3K)areaclassofenzymeswhichphosphorylatethethirdhydroxylgroupintheinositolringofphosphotidylinositols(PtdIns)presentontheinnerleafletofplasmamembrane.
ThereareeightdistinctPI3Kcatalyticsubunitsthatarecapableofphosphorylatinginositollipids.
PhosphorylationofPtdInsbyvariousPI3KsleadstoformationofPhosphatidylinositol3phosphate(PI(3)P),Phosphatidylinositol(3,4)-bisphosphate(PI(3,4)P2),andPhosphatidyl-inositol(3,4,5)-trisphosphate(PI(3,4,5)P3),whichareimportantsignalingmolecules.
Thesephosphoinositidesparticipateinsignaltransductionpathwaysbybindingtolipidbindingdomainslikethepleckstrindomainofwww.
proteomics-journal.
comPage15ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Akt/PKBandphosphoinositidedependentkinase-1(PDK1),zincfingerFYVEdomainandPXdomainsofproteins[97].
ActivationofPI3Kislinkedtoanincrediblydiversesetofkeycellularfunctions,includingcellgrowth,proliferation,motility,differentiation,survivalandintracellulartrafficking.
CellularPI3Kactivitiesarebalancedbyphosphoinositide3-phosphataseactivity,foundinthetumorsuppressorproteinPTENandinmembersofthemyotubularinfamily.
Alteredactivationofthesekinasesisassociatedwithvarioushumandiseasesincludingcancer.
PI3KsareactivateddownstreamtoGproteincoupledreceptors(GPCRs),receptortyrosinekinase(RTK),aswellasRAGE[98].
AGE/RAGEaxishasbeenreportedtotransactivateInsulinlikegrowthfactor1receptor(IGF-1R)causingactivationofPI3K/Aktin3T3adipocytesleadingtotheirdifferentiation.
UponbindingofAGEstoRAGE,itwasobservedthatRAGEcouplestoNADPHoxidasetostimulateSrckinase.
TheactivatedSrckinaseinturnphosphorylatesandactivatesIGF-1receptorandPI3K-Aktpathway.
ThisadipogenesiscausedduetoAGE/RAGEandactivationofPI3K/Aktcanleadtoobesity,orpredisposeapersontodiabetes,orcausediabeticcomplications[99].
Furthermore,activationofPI3KduetoAGE/RAGEhasbeenimplicatedindiabeticnephropathy,whichisassociatedwithdepositionofextracellularmatrixproteins(e.
g.
collagen)intheglomerularbasementmembrane.
AGE/RAGEinteractioncausedproductionofROSandactivationofSrckinasecausingPI3Kphosphorylation.
InterestinglyPI3KcauseddownstreamactivationofPKBandki-Ras,whichrecruitsERKarmoftheMAPK[100].
Diabeticnephropathyischaracterizedbyproteinuriawhichreflectsmalfunctioningofglomerularvisceralepithelialcells,namelypodocytes.
Clinicalandexperimentalevidenceshowsthatdiabeticnephropathyischaracterizedbypodocytehypertrophycausinganincreaseinslitlengthandwidth,andalsoreductioninthewww.
proteomics-journal.
comPage16ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
numberofpodocytesperglomerulus[101,102].
AGE/RAGEsignalinginpodocyteshasbeenshowntocausepodocyteapoptosis.
Inculturedmurinepodocytes,AGE/RAGEaxiscausesdephosphorylationofAktensuingintranscriptionalactivationofFOXO4andtherebyapoptosis[36].
TherenninangiotensinsystemresponsibleformaintainingbloodvolumeandpressureisalsoaffectedbyAGE/RAGEsignalingtocausepodocyteapoptosis.
ThiseffectwasdependentonPI3K/Aktmediatedphosphorylation[103].
Indiabeticuremia,neutrophilsexhibitedanincreasedadhesiontoAGE-collagenthroughinteractionwithRAGEbyactivationofPI3K/Aktpathway[104].
ThePI3K/Akt/mTORpathwayisanegativeregulatorofautophagy.
AGEsinduceproliferationofrataorticvascularsmoothmusclecells(VSMCs)byinducingautophagyviainactivationofPI3K/Aktpathwaytherebyacceleratingatherosclerosisindiabeticconditions[105].
Inhumanumbilicalvascularendothelialcells(HUVECs),AGEscauseROSmediatedactivationofPI3K/Akt/IкB/NF-кBandJNK/AP1leadingtoincreasedexpressionofE-selectin,whichisassociatedwithprogressionofatherosclerosis[78].
HyperlipidemiaisshowntoincreasetheexpressionofHMGB1inserum,heartandlungtissuesofatheroscleroticmice,whichstimulatesRAGEsignalingthroughactivationofAktandproductionofproinflammatorycytokines[106].
Lectinlikeoxidizedlowdensitylipoproteinreceptor1(LOX1)isascavengerreceptorofoxidizedlowdensitylipoproteins(oxLDL),knowntobeelevatedinatheroscleroticplaquesandimplicatedintheinitiationandprogressionofatherosclerosis.
AGE/RAGEinteractionisproposedtocausetheupregulationofLOX1inaorticendothelialcellsbydownstreamactivationofPI3KwhichphosphorylatesmTORC2andAkt,indirectlyplayingaroleinatherosclerosis[107].
Indiabeticcardiovascularwww.
proteomics-journal.
comPage17ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
disorders,AGE/RAGEinteractioninducestheexpressionofheparinaseandPI3K/Aktphosphorylationleadingtomigrationofmacrophages[108].
Indorsalrootgangliasensoryneurons,S100B/RAGEinteractionactivatesPI3K/AktandERKpathways,whichfurthercauseslipidperoxidationandcaspase-3activationresultinginneuronaldamage[109].
S100B/RAGEinteractionincreasestheexpressionandreleaseofchemokinesinmicrogliaresultinginactivationofSrc/Ras/PI3K/RhoA/ROCKpathwayalongwithp38,ERK,JNK-AP1andNF-кB[82].
Incarcinogenesis,hypoxiainducedexpressionofRAGEandP2X7RcausedincreasedphosphorylationofAktandERK,andNF-кBnucleartranslocation.
Further,NF-кBincreasedtheexpressionofMMP2and9contributingtocellularinvasivenessuponHMGB1treatment[110].
HMGB1/RAGEinteractionleadstogrowth,migrationandinvasionofcolorectalcarcinomacellsbythephosphorylationofERK1/2,Rac1,AktandproductionofMMP9.
WhereasAGEsledtotheproductionofiNOSandNF-кBp65toagreaterextentthanHMGB1highlightingthedifferentialeffectsofboththeligandsandsuggestingamorecloseassociationofHMGB1withcancer[111].
BindingofHMGB1toRAGEandTLR4inlungcancercellsisshowntocausetheactivationofPI3K/AktandERK1/2leadingtoincreasedproliferation.
Thiseffectwasobservedduetodecreasedexpressionofpro-apoptoticBaxproteinandincreasedlevelsofanti-apoptoticBcl-2protein[112].
TreatmentofCML-BSAtoHEK293causedphosphorylationofthep66adaptorproteintocausedownstreamphosphorylationofAkt.
ActivatedAktcausesthephosphorylationofforkheadtranscriptionfactor3referredtoasFKHRL1leadingtoitsinactivationandpreventingitsnucleartranslocationtherebysuppressingtheexpressionofmanganesesuperoxidedismutase(MnSOD).
DownregulationofMnSODfacilitatesROSproduction.
TheseeffectsweresuppressedinAGEReceptor-1(AGER1)overexpressingwww.
proteomics-journal.
comPage18ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
HEK293cellssuggestingtheprotectiveeffectofAGER-1againstoxidativestresscausedbyAGE/RAGEinteraction[113].
S100A8/A9causesgrowthofnormalaswellastransformedhumanepidermalkeratinocytesbyinteractionwithRAGE/DNAX-activatingprotein(DAP10)heterodimersanddownstreamactivationofAkt.
Thisinteractioninducedapoptosisbyactivationofcaspase8innormalhumanepidermalkeratinocytes,whereassustainedgrowthwasobservedinthetransformedcelllinesHaCaTandA431duetoenhancedactivationofAkt,suggestingvariableeffectsofAktdownstreamtoRAGE/DAP10[114].
JanusActivatedkinase(JAK)JAKisafamilyoftyrosinekinasesthatactivatesignaltransducersandactivatorsoftranscription(STAT)inresponsetocytokinesandgrowthfactorsaswellasbyRAGEactivation.
JAKphosphorylatesSTATresultinginitsdimerizationandnucleartranslocation.
STATregulatestheexpressionofgenesinvolvedinantiviraldefence,embryogenesis,cellgrowth,cellsurvival,andcellmotility[115-120].
ConstitutiveactivationofmembersofSTATfamilyisassociatedwithseveraltypesofcancers.
TyrosinephosphorylatedSTAT1,3,and5arepersistentlyfoundinmostofthemalignancies.
AGE/RAGEaxisactivatesJAKinmyeloidleukemiawhereitinducesSTAT3phosphorylation[121-124].
RAGEpromotespancreaticcancerbyregulatingconjugationofATG12-ATG5,autophagyrelatedproteins;andBECN1-PIK3C3(Beclin1-Phosphatidylinositol3-kinasecatalyticsubunittype3)complexformation,whichresultsinsecretionofIL6,andmitochondrialSTAT3activation[125,126].
AGE/RAGEinteractionincreasestyrosinephosphorylationofHsp70,cyclinE,andcyclinD1inJAKdependentmanner.
ItalsoincreasesHsp70proteininteractionwithSTAT1,STAT3,STAT5b,cyclinD1,andcyclinE.
ThesefindingsindicatethatAGEsinducemitogenesisbyactivatingtheJAK2,STAT1,STAT3,andSTAT5-www.
proteomics-journal.
comPage19ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
cyclinD1pathwayinNRK-49Fcells[127].
Whileinhepatocytes,RAGEshowsanti-tumoureffectsbydecreasingp65levels,NF-кBactivation,cyclinD1productionandIL6mediatedSTAT3phosphorylation,therebydownregulatingPim1andsuppressingthehyperplasticresponse.
BlockadeofRAGErescuesliverremnantsfromadaptiveproliferationtriggeredbyMyd88signaling[128].
Indiabeticnephropathy,JAK2-STAT1/STAT3pathwayisinvolvedinAGE-inducedtype-1collagenproductioninkidneyfibroblastcells,whichisconsideredasamajorcauseofrenalfibrosis[129].
JAK/STATsignalingisalsoinvolvedinthedevelopmentofosteoarthritis.
IL-7activatesJAK/STATpathwayinchondrocytesleadingtoincreasedsecretionofS100A4whichinteractswithRAGEinautocrinemannerandstimulatesMMP13production[130].
AGEinducesactivationofMMP13,disintegrinandmetalloproteinasewiththrombospondinmotifs(ADAMTS)inaJAK/STAT3dependentmanner,whichisinvolvedincollagenIIandproteoglycanproductionandcontributestoosteoarthritis[131].
Duringembryogenesis,RAGEisinvolvedinneuronaldifferentiation.
LigandssuchasHumanglycatedalbumin,S100BandHMGB1arefoundtoincreasetheneuritegrowth.
TheseligandsactivateJAK/STAT,ERKandNF-кBwhichplayanimportantroleinaxonaloutgrowth[132].
GlycogenSynthaseKinase-3beta(GSK3β)GSK3βisaprolinedirectedserine/threoninekinase.
Itwasinitiallyidentifiedasakinasethatphosphorylatesglycogensynthase,akeyenzymeinglycogensynthesis[133,134].
Apartfromitsinvolvementinglycogensynthesis,itplaysanimportantroleinseveralbiologicalprocessessuchasdifferentiation,neurogenesis,cellfatedetermination,proliferation,andsurvival[135].
GSK3βactivityisdeterminedbyitsphosphorylationstatus;phosphorylationofser9decreasesitsactivity,whereasdephosphorylationactivatesit.
Itsactivityisalsowww.
proteomics-journal.
comPage20ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
enhancedbyphosphorylationoftyrosine216[136-138].
GSK3βphosphorylationisregulatedbyseveralkinasesincludingERKs,MAPKAPK-1,Akt,fewisoformsofPKC,proteinkinaseAandp70S6kinase.
Itisassociatedwithnumerousdisordersincludingdiabetes,cardiovasculardisorders,Parkinson's,Alzheimer'sdisease,amyotrophiclateralsclerosis,multiplesclerosisandcancer[139-141].
AGE/RAGEdependentGSK3βactivationinducesTauhyperphosphorylationinSK-N-SHcells,primaryhippocampalcellsandratbrain.
Inneuronalcells,AGE/RAGEinteractionleadstoactivationofGSK3β,ERK,p38MAPK,whichareassociatedwithmemorydeterioration,declineofsynapticproteins,andimpairmentoflong-termpotentiation.
BlockadeofRAGEattenuatedalltheAGEinducedimpairments,suggestingavitalroleofAGE/RAGEmediatedGSK3βstimulationinprogressionofAlzheimer'sdisease[142,143].
MethylglyoxalinducesGSK3βandp38MAPKactivationwhichleadstotauhyperphosphorylationinNeuro-2acells[144,145].
ThisisperhapsduetoincreasedRAGEsignalingasaresultofAGEformationpromotedbymethylglyoxal.
Apartfromtaupathology,GSK3βisalsoinvolvedinAβsynthesisandapoptoticneuronaldeath,whicharethemajorhallmarksofAD[146].
AGE/RAGEinteractionleadstoGSK3βactivationduringhypoxia/reoxygenationofcardiomyocyte[70].
AGEaccumulationandRAGEexpressionissignificantlyincreasedindiabeticratswithcardiomyopathyalongwithmarkedincreaseinGSK3βactivitywhichisreducedaftercurcumintreatmentbyAktphosphorylation[147].
However,therearenostudiesshowingRAGEdependentGSK3βactivationwithrespecttootherRAGEligandsincludingAβ,HMGB1andS100B,whicharefoundtobesignificantlyincreasedinAlzheimer'sdisease,cardiovasculardisordersandcancer[148-151].
www.
proteomics-journal.
comPage21ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
AnalysisofglobalphosphoproteomeinRAGEsignalingRAGEcanpotentiallyinteractwithmultipleligandsandactivateseveraldifferentsignaltransductionpathwaysinvolvingvariousproteinkinasesasdescribedabove.
Thesekinasesphosphorylatedownstreamsignalingmoleculesandcausespecificcellularresponse.
Askinaseshavemultiplesubstrates,thephosphorylationofasubstratedependsonitsexpression,localization,andcelltype.
OnesuchexampleisGSK3β,whichphosphorylatesglycogensynthaseinhepatocytesandregulatesglycogenmetabolism.
Whereas,inhumanembryonickidneycells(HEK293T),itphosphorylatesbetacateninrenderingittoproteasomaldegradation[152],andinneuronalcells,GSK3βactsdownstreamtoRAGEandhyperphosphorylatestaucausingneuronaldamage.
ThisobservationimpelledustoanalysephosphorylationsubstratesofkinasesinvolvedinRAGEsignaling.
Here,wehavefocusedonfewmajorkinases,namelyPKC,ERK,PI3K/Akt,p38,JAK,GSK3βandJNK,allofwhichareknowntoactdownstreamofRAGE.
AcomprehensivelistofproteinsubstratesforthesekinaseswasminedfromPhosPhoSitePlusdatabase[153].
Adetailedworkflowofdataminingandanalysisofphosphosubstratesisdepictedintheflowchart(Fig3).
Altogether,1045differentsubstrateswerefoundtobeassociatedwiththese7kinases(SupplementaryFig1).
Amongstthese7kinases,PKCandERKappeartophosphorylatemaximumnumberofsubstrates,andsignificantnumbersofthesephosphosubstratesareuniquetothesetwokinases(SupplementaryTable1and2),manyarealsosharedwithotherkinases.
Oneinterestingexampleisp21Cip1(p21Waf1),aproteininvolvedincellcycleregulation,wasfoundtobephosphorylatedbysixdifferentkinasesinthisanalysis.
Interestingly,itwasalsoshowntobephosphorylatedinresponsetoRAGEmediatedsignaling.
Overexpressionofwww.
proteomics-journal.
comPage22ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
RAGEcausedsignificantincreaseinp21Waf1induction,whichregulatescellproliferationinL6myoblasts[40].
Further,wehaveanalyzedtheassociationofthesephosphosubstrateswithdiseasessuchascancer,diabetes,cardiovascularandneurodegenerativediseases(Fig4).
Outofthe1045phosphosubstrates,thereare772,65,27and10phosphosubstratesfoundtobeassociatedwithcancer,neurodegenerativediseases,diabetesandcardiovasculardisordersrespectively(Fig3,SupplementaryTable3).
AninterestingobservationisthatthenumberofphosphosubstratesassociatedwithcancerisquitelargeandmanyofthesecanbepotentiallyphophorylatedbyrespectivekinasesinresponsetoRAGEactivation.
Insupportofthisnotion,involvementofRAGEincancerisbecomingincreasinglyevidentfromseveralstudies[44-53].
ERK2,akeyplayerinMAPKsignalingistheonlyphosphosubstratethatisassociatedwithallfourdiseases.
Inaddition,wehaveidentified11phosphosubstratesassociatedwiththreeofthediseases,whichincludeMARCKS,MAP4,Tau-8,Akt1,GSK3β,eNOS,ERK1,p70S6K,JNK1,AMPKA1andCREB.
ExceptMARCKSandMAP4,allotherproteinsareshowntobeinvolvedinRAGEsignaling,whichsupportsthisanalysis.
Tauiso8andMAP4arecytoskeletalproteinswhichplayanimportantroleinmicrotubuleassembly.
FunctionallossofTauiso8isassociatedwithseveralneurodegenerativedisordersincludingfrontotemporaldementia,Alzheimer'sdisease,pick'sdiseaseetc[154].
Endothelialnitricoxidesynthase(eNOS)producesnitricoxideinbloodvesselsandactsasaregulatorofenergymetabolisminthebody.
Itisassociatedwithseveraldisordersincludingdiabetes,hypertension,cancer,Alzheimer's,cardiovasculardisease,stroke,andatherosclerosis[155].
AGEshaveshowntoreduceeNOSphosphorylationanddown-regulationviaRAGEcausingendothelialdysfunction[156,157].
p70S6Kisaserinethreoninekinasethatactswww.
proteomics-journal.
comPage23ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
downstreamtoPI3K/Akt/mTORpathway.
Itphosphorylatesribosomalproteinandinducesproteinsynthesis.
ChronicintakeofAGEsmediatesp70S6KphosphorylationandRAGEup-regulationinrats.
Itisimplicatedinmultiplediseasesincludinginsulinresistance,skeletalmusclehypertrophyaswellasageing[158-160].
AMPKA1isacatalyticsubunitofAMPactivatedproteinkinaseknowntobeinvolvedinseveralbiologicalprocessessuchaslipidbiosynthesis,glucosehomeostasis,apoptosis,signaltransductionetc.
AβinducestheformationofautophagosomesinbrainsofAPP/PS1transgenicmiceaswellasinSH-SY5YcellsbyinteractionwithRAGEbyAMPKactivation[161,162].
CREBisatranscriptionfactorwhichbindswiththecAMPresponsiveelementandregulatestheexpressionofseveralgenes.
DeregulationofCREBfunctionisassociatedwithdisorderslikeAlzheimer'sdisease,huntington'sdiseaseandcancer.
AmphoterininducedCREBphosphorylationviaRAGEisinvolvedintheexpressionofchromograninsAandBinneuroblastomacellsleadingtotheirdifferentiation[163-165].
Myristoylatedalanine-richC-kinasesubstrate(MARCKS)isanacidicproteininvolvedintheregulationofcellproliferation,cellularsecretionandtransport,cellintegrityandmotility.
ThephosphorylationstatusofthisproteininRAGEsignalingneedstobeinvestigated[166,167].
ThisanalysisclearlysuggestedthatRAGEmediatedkinaseactivationcanpotentiallyresultinphosphorylationofseveraloftheirsubstrates.
Thepossibilityofaproteingettingphosphorylatedismorewhenitisasharedasasubstratebymultiplekinases.
Therefore,itisimportantnottounderminetheinvolvementofvariousknownsubstratesofthekinasesactivatedinRAGEsignaling.
Inconclusion,thisreviewemphasizesonthevitalroleofRAGEsignalinginvariouscellularprocessessuchasproliferation,apoptosis,migration,inflammation,immuneresponse,autophagyetc,andprovidesinsightintheplausibleroleofRAGEactivatedphosphoproteomeinseveraldiseases.
www.
proteomics-journal.
comPage24ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Acknowledgement:ThisworkissupportedbyCSIR-NetworkProjectBSC0111.
KBB,SBBandRSKacknowledgeCSIRandUGCforresearchfellowship.
GVPisgratefultoLadyTataMemorialTrustforaresearchfellowship.
Theauthorshavedeclarednoconflictofinterest.
References1.
Schmidt,A.
M.
,DuYan,S.
,Yan,S.
F.
,Stern,D.
M.
,ThemultiligandreceptorRAGEasaprogressionfactoramplifyingimmuneandinflammatoryresponses.
JClinInvest.
2001,108,949-955.
2.
Harashima,A.
,Yamamoto,Y.
,Cheng,C.
,Tsuneyama,K.
etal.
,Identificationofmouseorthologueofendogenoussecretoryreceptorforadvancedglycationend-products:structure,functionandexpression.
BiochemJ.
2006,396,109-115.
3.
Huttunen,H.
J.
,Fages,C.
,Rauvala,H.
,Receptorforadvancedglycationendproducts(RAGE)-mediatedneuriteoutgrowthandactivationofNF-кBrequirethecytoplasmicdomainofthereceptorbutdifferentdownstreamsignalingpathways.
JBiolChem.
1999,274,19919-19924.
4.
Kalea,A.
Z.
,Schmidt,A.
M.
,Hudson,B.
I.
,AlternativesplicingofRAGE:rolesinbiologyanddisease.
FrontBiosci.
2011,17,2756-2770.
5.
Kulkarni,M.
J.
,Korwar,A.
M.
,Mary,S.
,Bhonsle,H.
S.
,Giri,A.
P.
,Glycatedproteome:Fromreactiontointervention.
ProteomicsClinAppl.
2013,7,155-170.
6.
Kislinger,T.
,Fu,C.
,Huber,B.
,Qu,W.
etal.
,Nε-(carboxymethyl)lysineadductsofproteinsareligandsforreceptorforadvancedglycationendproductsthatactivatecellsignalingpathwaysandmodulategeneexpression.
JBiolChem.
1999,274,31740-31749.
www.
proteomics-journal.
comPage25ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
7.
Wang,Z.
,Jiang,Y.
,Liu,N.
,Ren,L.
etal.
,Advancedglycationend-productNε-carboxymethyl-Lysineacceleratesprogressionofatheroscleroticcalcificationindiabetes.
Atherosclerosis2012,221,387-396.
8.
Reynolds,T.
M.
,CHEMISTRYOFNONENZYMICBROWNING.
I.
THEREACTIONBETWEENALDOSESANDAMINES.
AdvFoodRes.
1963,12,1-52.
9.
Bunn,H.
F.
,Nonenzymaticglycosylationofprotein:relevancetodiabetes.
AmJMed.
1981,70,325-330.
10.
Ahmed,M.
U.
,Dunn,J.
A.
,Walla,M.
D.
,Thorpe,S.
R.
,Baynes,J.
W.
,Oxidativedegradationofglucoseadductstoprotein.
Formationof3-(Nepsilon-lysino)-lacticacidfrommodelcompoundsandglycatedproteins.
JBiolChem.
1988,263,8816-8821.
11.
Ahmed,M.
U.
,BrinkmannFrye,E.
,Degenhardt,T.
P.
,Thorpe,S.
R.
,Baynes,J.
W.
,N-epsilon-(carboxyethyl)lysine,aproductofthechemicalmodificationofproteinsbymethylglyoxal,increaseswithageinhumanlensproteins.
BiochemJ.
1997,324,565-570.
12.
Dyer,D.
G.
,Blackledge,J.
A.
,Thorpe,S.
R.
,Baynes,J.
W.
,Formationofpentosidineduringnonenzymaticbrowningofproteinsbyglucose.
Identificationofglucoseandothercarbohydratesaspossibleprecursorsofpentosidineinvivo.
JBiolChem.
1991,266,11654-11660.
13.
Beisswenger,P.
J.
,Szwergold,B.
S.
,Yeo,K.
T.
,Glycatedproteinsindiabetes.
ClinLabMed.
2001,21,53-78.
14.
Charonis,A.
S.
,Tsilibary,E.
C.
,StructuralandfunctionalchangesoflamininandtypeIVcollagenafternonenzymaticglycation.
Diabetes1992,41,49-51.
www.
proteomics-journal.
comPage26ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
15.
Chen,L.
,Wei,Y.
,Wang,X.
,He,R.
,Ribosylationrapidlyinducesalpha-synucleintoformhighlycytotoxicmoltenglobulesofadvancedglycationendproducts.
PLoSOne2010,5,e9052.
16.
Bansode,S.
B.
,Chougale,A.
D.
,Joshi,R.
S.
,Giri,A.
P.
etal.
,Proteomicanalysisofproteaseresistantproteinsinthediabeticratkidney.
MolCellProteomics2013,12,228-236.
17.
Neeper,M.
,Schmidt,A.
M.
,Brett,J.
,Yan,S.
D.
etal.
,Cloningandexpressionofacellsurfacereceptorforadvancedglycosylationendproductsofproteins.
JBiolChem.
1992,267,14998-15004.
18.
Alexiou,P.
,Chatzopoulou,M.
,Pegklidou,K.
,Demopoulos,V.
J.
,RAGE:amulti-ligandreceptorunveilingnovelinsightsinhealthanddisease.
CurrMedChem.
2010,17,2232-2252.
19.
DuYan,S.
,Chen,X.
,Fu,J.
,Chen,M.
etal.
,RAGEandamyloid-βpeptideneurotoxicityinAlzheimer'sdisease.
Nature1996,382,685-691.
20.
Hartmann,T.
,Bieger,S.
C.
,Brühl,B.
,Tienari,P.
J.
etal.
,DistinctsitesofintracellularproductionforAlzheimer'sdiseaseAβ40/42amyloidpeptides.
NatMed.
1997,3,1016-1020.
21.
Takuma,K.
,Fang,F.
,Zhang,W.
,Yan,S.
etal.
,RAGE-mediatedsignalingcontributestointraneuronaltransportofamyloid-βandneuronaldysfunction.
ProcNatlAcadSciUSA2009,106,20021-20026.
22.
Ostendorp,T.
,Leclerc,E.
,Galichet,A.
,Koch,M.
etal.
,StructuralandfunctionalinsightsintoRAGEactivationbymultimericS100B.
EMBOJ.
2007,26,3868-3878.
23.
Huttunen,H.
J.
,Kuja-Panula,J.
,Sorci,G.
,Agneletti,A.
L.
etal.
,CoregulationofneuriteoutgrowthandcellsurvivalbyamphoterinandS100proteinsthroughwww.
proteomics-journal.
comPage27ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
receptorforadvancedglycationendproducts(RAGE)activation.
JBiolChem.
2000,275,40096-40105.
24.
Sims,G.
P.
,Rowe,D.
C.
,Rietdijk,S.
T.
,Herbst,R.
,Coyle,A.
J.
,HMGB1andRAGEininflammationandcancer.
AnnuRevImmunol.
2009,28,367-388.
25.
Shaw,S.
S.
,Schmidt,A.
M.
,Banes,A.
K.
,Wang,X.
etal.
,S100B/RAGE-mediatedaugmentationofangiotensinII-inducedactivationofJAK2invascularsmoothmusclecellsisdependentonPLD2.
Diabetes2003,52,2381-2388.
26.
Klune,J.
R.
,Dhupar,R.
,Cardinal,J.
,Billiar,T.
R.
,Tsung,A.
,HMGB1:endogenousdangersignaling.
MolMed.
2008,14,476-484.
27.
Taguchi,A.
,Blood,D.
C.
,delToro,G.
,Canet,A.
etal.
,BlockadeofRAGE-amphoterinsignalingsuppressestumourgrowthandmetastases.
Nature2000,405,354-360.
28.
Mulder,D.
J.
,Water,T.
V.
D.
,Lutgers,H.
L.
,Graaff,R.
etal.
,Skinautofluorescence,anovelmarkerforglycemicandoxidativestress-derivedadvancedglycationendproducts:anoverviewofcurrentclinicalstudies,evidence,andlimitations.
DiabetesTechnolTher.
2006,8,523-535.
29.
Meerwaldt,R.
,Links,T.
,Graaff,R.
,Thorpe,S.
R.
etal.
,Simplenoninvasivemeasurementofskinautofluorescence.
AnnNYAcadSci.
2005,1043,290-298.
30.
Lutgers,H.
L.
,Graaff,R.
,Links,T.
P.
,Ubink-Veltmaat,L.
J.
etal.
,Skinautofluorescenceasanoninvasivemarkerofvasculardamageinpatientswithtype2diabetes.
DiabetesCare2006,29,2654-2659.
31.
Ramasamy,R.
,Yan,S.
F.
,Schmidt,A.
M.
,RAGE:therapeutictargetandbiomarkeroftheinflammatoryresponse-theevidencemounts.
JLeukocBiol.
2009,86,505-512.
www.
proteomics-journal.
comPage28ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
32.
Cargnello,M.
,Roux,P.
P.
,ActivationandfunctionoftheMAPKsandtheirsubstrates,theMAPK-activatedproteinkinases.
MicrobiolMolBiolRev.
2011,75,50-83.
33.
Yeh,C.
H.
,Sturgis,L.
,Haidacher,J.
,Zhang,X.
N.
etal.
,Requirementforp38andp44/p42mitogen-activatedproteinkinasesinRAGE-mediatednuclearfactor-kappaBtranscriptionalactivationandcytokinesecretion.
Diabetes2001,50,1495-1504.
34.
Tanikawa,T.
,Okada,Y.
,Tanikawa,R.
,Tanaka,Y.
,AdvancedglycationendproductsinducecalcificationofvascularsmoothmusclecellsthroughRAGE/p38MAPK.
JVascRes.
2009,46,572-580.
35.
Chang,P.
C.
,Chen,T.
H.
,Chang,C.
J.
,Hou,C.
C.
etal.
,Advancedglycosylationendproductsinduceinduciblenitricoxidesynthase(iNOS)expressionviaap38MAPK-dependentpathway.
KidneyInt.
2004,65,1664-1675.
36.
Chuang,P.
Y.
,Yu,Q.
,Fang,W.
,Uribarri,J.
,He,J.
C.
,AdvancedglycationendproductsinducepodocyteapoptosisbyactivationoftheFOXO4transcriptionfactor.
KidneyInt.
2007,72,965-976.
37.
Chung,A.
C.
,Zhang,H.
,Kong,Y.
Z.
,Tan,J.
J.
etal.
,Advancedglycationend-productsinducetubularCTGFviaTGFbeta-independentSmad3signaling.
JAmSocNephrol.
2010,21,249-260.
38.
Chen,S.
C.
,Guh,J.
Y.
,Hwang,C.
C.
,Chiou,S.
J.
etal.
,Advancedglycationend-productsactivateextracellularsignal-regulatedkinaseviatheoxidativestress-EGFreceptorpathwayinrenalfibroblasts.
JCellBiochem.
2010,109,38-48.
39.
Li,J.
H.
,Huang,X.
R.
,Zhu,H.
J.
,Oldfield,M.
etal.
,AdvancedglycationendproductsactivateSmadsignalingviaTGF-β-dependentand-independentwww.
proteomics-journal.
comPage29ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
mechanisms:implicationsfordiabeticrenalandvasculardisease.
FASEBJ.
2004,18,176-178.
40.
Li,Y.
,Liu,S.
,Zhang,Z.
,Xu,Q.
etal.
,RAGEmediatesaccelerateddiabeticveingraftatherosclerosisinducedbycombinedmechanicalstressandAGEsviasynergisticERKactivation.
PLoSOne2012,7,e35016.
41.
Xu,D.
,Young,J.
H.
,Krahn,J.
M.
,Song,D.
etal.
,StableRAGE–heparansulfatecomplexesareessentialforsignaltransduction.
ACSChemBiol.
2013,8,1611-1620.
42.
Spiekerkoetter,E.
,Guignabert,C.
,deJesusPerez,V.
,Alastalo,T.
P.
etal.
,S100A4andbonemorphogeneticprotein-2codependentlyinducevascularsmoothmusclecellmigrationviaphospho-extracellularsignal-regulatedkinaseandchlorideintracellularchannel4.
CircRes.
2009,105,639-647.
43.
Shi,L.
,Yu,X.
,Yang,H.
,Wu,X.
,AdvancedglycationendproductsinducehumancornealepithelialcellsapoptosisthroughgenerationofreactiveoxygenspeciesandactivationofJNKandp38MAPKpathways.
PLoSONE2013,8,ArticleIDe66781.
44.
Ishihara,K.
,Tsutsumi,K.
,Kawane,S.
,Nakajima,M.
,Kasaoka,T.
,Thereceptorforadvancedglycationend-products(RAGE)directlybindstoERKbyaD-domain-likedockingsite.
FEBSLett.
2003,550,107-113.
45.
Riuzzi,F.
,Sorci,G.
,Donato,R.
,TheAmphoterin(HMGB1)/receptorforadvancedglycationendproducts(RAGE)pairmodulatesmyoblastproliferation,apoptosis,adhesiveness,migration,andInvasiveness:FunctionalinactivationofRAGEinL6myoblastsresultsintumorformationinvivo.
JBiolChem.
2006,281,8242-8253.
www.
proteomics-journal.
comPage30ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
46.
Riuzzi,F.
,Sorci,G.
,Donato,R.
,RAGEexpressioninrhabdomyosarcomacellsresultsinmyogenicdifferentiationandreducedproliferation,migration,invasiveness,andtumorgrowth.
AmJPathol.
2007,171,947-961.
47.
Taguchi,A.
,Blood,D.
C.
,delToro,G.
,Canet,A.
etal.
,BlockadeofRAGE–amphoterinsignallingsuppressestumourgrowthandmetastases.
Nature2000,405,354-360.
48.
Ghavami,S.
,Rashedi,I.
,Dattilo,B.
M.
,Eshraghi,M.
etal.
,S100A8/A9atlowconcentrationpromotestumorcellgrowthviaRAGEligationandMAPkinase-dependentpathway.
JLeukocBiol.
2008,83,1484-1492.
49.
Lin,L.
,Zhong,K.
,Sun,Z.
,Wu,G.
,Ding,G.
,Receptorforadvancedglycationendproducts(RAGE)partiallymediatesHMGB1-ERKsactivationinclearcellrenalcellcarcinoma.
JCancerResClinOncol.
2012,13,11-22.
50.
Kao,Y.
H.
,Lin,Y.
C.
,Tsai,M.
S.
,Sun,C.
K.
etal.
,InvolvementofthenuclearhighmobilitygroupB1peptidesreleasedfrominjuredhepatocytesinmurinehepaticfibrogenesis.
BiochimBiophysActa2014,1842,1720-1732.
51.
Fuentes,M.
,Nigavekar,S.
,Arumugam,T.
,Logsdon,C.
etal.
,RAGEActivationbyS100PinColonCancerStimulatesGrowth,Migration,andCellSignalingPathways.
DisColonRectum2007,50,1230-1240.
52.
Tsoporis,J.
N.
,Izhar,S.
,Proteau,G.
,Slaughter,G.
,Parker,T.
G.
,S100B-RAGEdependentVEGFsecretionbycardiacmyocytesinducesmyofibroblastproliferation.
JMolCellCardiol.
2012,52,464–473.
53.
Tsoporis,J.
N.
,Izhar,S.
,Leong-Poi,H.
,Desjardins,J.
F.
etal.
,S100Binteractionwiththereceptorforadvancedglycationendproducts(RAGE):anovelreceptor-www.
proteomics-journal.
comPage31ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
mediatedmechanismformyocyteapoptosispostinfarction.
CircRes.
2010,106,93-101.
54.
Liu,R.
,Wu,C.
X.
,Zhou,D.
,Yang,F.
etal.
,Pinocembrinprotectsagainstβ-amyloid-inducedtoxicityinneuronsthroughinhibitingreceptorforadvancedglycationendproducts(RAGE)-independentsignalingpathwaysandregulatingmitochondrion-mediatedapoptosis.
BMCMed.
2012,10,105.
55.
Origlia,N.
,Righi,M.
,Capsoni,S.
,Cattaneo,A.
etal.
,ReceptorforAdvancedGlycationEndProduct-DependentActivationofp38Mitogen-ActivatedProteinKinaseContributestoAmyloid-β-MediatedCorticalSynapticDysfunction.
JNeurosci.
2008,28,3521-3530.
56.
Askarova,S.
,Yang,X.
,Sheng,W.
,Sun,G.
Y.
,Lee,J.
C.
,RoleofAbeta-receptorforadvancedglycationendproductsinteractioninoxidativestressandcytosolicphospholipaseA(2)activationinastrocytesandcerebralendothelialcells.
Neuroscience2011,199,375-385.
57.
Slowik,A.
,Merres,J.
,Elfgen,A.
,Jansen,S.
etal.
,Involvementofformylpeptidereceptorsinreceptorforadvancedglycationendproducts(RAGE)andamyloidbeta1-42-inducedsignaltransductioninglialcells.
Mol.
Neurodeg.
2012,7,55.
58.
McDonald,D.
,Bamberger,M.
,Combs,C.
,Landreth,G.
,b-amyloidfibrilactivateparallelmitogen-activatedproteinkinasepathwaysinmicrogliaandTHP-1monocytes.
JNeurosci.
1998,18,4451-4460.
59.
Kang,J.
H.
,Hwang,S.
M.
,Chung,I.
Y.
,S100A8,S100A9,andS100A12activateairwayepithelialcellstoproduceMUC5ACviaERKandNF-κBpathways.
2014www.
proteomics-journal.
comPage32ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
60.
Wolfson,R.
K.
,Chiang,E.
T.
,Garcia,J.
G.
,HMGB1induceshumanlungendothelialcellcytoskeletalrearrangementandbarrierdisruption.
Microvasc.
Res.
2011,81,189-197.
61.
He,Z.
W.
,Qin,Y.
H.
,Wang,Z.
W.
,Chen,Y.
etal.
,HMGB1actsinsynergywithlipopolysaccharideinactivatingrheumatoidsynovialfibroblastsviap38MAPKandNF-kappaBsignalingpathways.
MediatorsInflamm.
2013,596716.
62.
Qin,Y.
H.
,Dai,S.
M.
,Tang,G.
S.
,Zhang,J.
etal.
,HMGB1enhancestheproinflammatoryactivityoflipopolysaccharidebypromotingthephosphorylationofMAPKp38throughreceptorforadvancedglycationendproducts.
JImmunol.
2009,183,6244-6250.
63.
Weston,C.
R.
,Davis,R.
J.
,TheJNKsignaltransductionpathway.
CurrOpinGenetDev.
2002,12,14-21.
64.
Bogoyevitch,M.
A.
,Kobe,B.
,UsesforJNK:themanyandvariedsubstratesofthec-JunN-terminalkinases.
MicrobiolMolBiolRev.
2006,70,1061-1095.
65.
Nah,S.
S.
,Choi,I.
Y.
,Lee,C.
K.
,Oh,J.
S.
etal.
,EffectsofadvancedglycationendproductsontheexpressionofCOX-2,PGE2andNOinhumanosteoarthriticchondrocytes.
Rheumatology2008,47,425-431.
66.
Nah,S.
S.
,Choi,I.
Y.
,Yoo,B.
,Kim,Y.
G.
etal.
,Advancedglycationendproductsincreasesmatrixmetalloproteinase-1,-3,and-13,andTNF-αinhumanosteoarthriticchondrocytes.
FEBSlett.
2007,581,1928-1932.
67.
Zhang,F.
,Banker,G.
,Liu,X.
,Suwanabol,P.
A.
etal.
,ThenovelfunctionofadvancedglycationendproductsinregulationofMMP9production.
JSurgRes.
2011,171,871-876.
www.
proteomics-journal.
comPage33ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
68.
Alikhani,M.
,MacLellan,C.
M.
,Raptis,M.
,Vora,S.
etal.
,AdvancedglycationendproductsinduceapoptosisinfibroblaststhroughactivationofROS,MAPkinases,andtheFOXO1transcriptionfactor.
AmJPhysiolCellPhysiol.
2007,292,C850-C856.
69.
Chang,J.
S.
,Wendt,T.
,Qu,W.
,Kong,L.
etal.
,Oxygendeprivationtriggersupregulationofearlygrowthresponse-1bythereceptorforadvancedglycationendproducts.
CircRes.
2008,102,905-913.
70.
Shang,L.
,Ananthakrishnan,R.
,Li,Q.
,Quadri,N.
etal.
,RAGEmodulateshypoxia/reoxygenationinjuryinadultmurinecardiomyocytesviaJNKandGSK-3βsignalingpathways.
PLoSOne2010,5,e10092.
71.
Aleshin,A.
,Ananthakrishnan,R.
,Li,Q.
,Rosario,R.
etal.
,RAGEmodulatesmyocardialinjuryconsequenttoLADinfarctionviaimpactonJNKandSTATsignalinginamurinemodel.
AmJPhysiolHeartCircPhysiol.
2008,294,H1823-H1832.
72.
Volz,H.
C.
,Laohachewin,D.
,Seidel,C.
,Lasitschka,F.
etal.
,S100A8/A9aggravatespost-ischemicheartfailurethroughactivationofRAGE-dependentNF-κBsignaling.
BasicResCardiol.
2012,107,1-16.
73.
Andrassy,M.
,Volz,H.
C.
,Igwe,J.
C.
,Funke,B.
etal.
,High-mobilitygroupbox-1inischemia-reperfusioninjuryoftheheart.
Circulation2008,117,3216-3226.
74.
Volz,H.
C.
,Seidel,C.
,Laohachewin,D.
,Kaya,Z.
etal.
,HMGB1:themissinglinkbetweendiabetesmellitusandheartfailure.
BasicResCardiol.
2010,105,805-820.
75.
Koh,E.
J.
,Yoon,S.
J.
,Lee,S.
M.
,Losartanprotectsliveragainstischaemia/reperfusioninjurythroughPPAR‐γactivationandreceptorforwww.
proteomics-journal.
comPage34ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
advancedglycationend‐productsdown‐regulation.
BrJPharmacol.
2013,169,1404-1416.
76.
Kokkola,R.
,Andersson,A.
,Mullins,G.
,Ostberg,T.
etal.
,RAGEisthemajorreceptorfortheproinflammatoryactivityofHMGB1inrodentmacrophages.
ScandJImmunol.
2005,61,1-9.
77.
Harja,E.
,Bu,D.
X.
,Hudson,B.
I.
,Chang,J.
S.
etal.
,VascularandinflammatorystressesmediateatherosclerosisviaRAGEanditsligandsinapoE–/–mice.
JClinInvest.
2008,118,183-194.
78.
Higai,K.
,Shimamura,A.
,Matsumoto,K.
,Amadori-modifiedglycatedalbuminpredominantlyinducesE-selectinexpressiononhumanumbilicalveinendothelialcellsthroughNADPHoxidaseactivation.
Clinchimacta.
2006,367,137-143.
79.
Fiuza,C.
,Bustin,M.
,Talwar,S.
,Tropea,M.
etal.
,Inflammation-promotingactivityofHMGB1onhumanmicrovascularendothelialcells.
Blood.
2003,101,2652-2660.
80.
Bianchi,R.
,Adami,C.
,Giambanco,I.
,Donato,R.
,S100BbindingtoRAGEinmicrogliastimulatesCOX-2expression.
JLeukocbiol.
2007,81,108-118.
81.
Bianchi,R.
,Giambanco,I.
,Donato,R.
,S100B/RAGE-dependentactivationofmicrogliaviaNF-κBandAP1:Co-regulationofCOX-2expressionbyS100B,IL-1βandTNF-α.
NeurobiolAging2010,31,665-677.
82.
Bianchi,R.
,Kastrisianaki,E.
,Giambanco,I.
,Donato,R.
,S100BproteinstimulatesmicrogliamigrationviaRAGE-dependentup-regulationofchemokineexpressionandrelease.
JBiolChem.
2011,286,7214-7226.
www.
proteomics-journal.
comPage35ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
83.
Esposito,G.
,Scuderi,C.
,Lu,J.
,Savani,C.
etal.
,S100BinducestauproteinhyperphosphorylationviaDickopff‐1up‐regulationanddisruptstheWntpathwayinhumanneuralstemcells.
JCellMolMed.
2008,12,914-927.
84.
Onyango,I.
G.
,Tuttle,J.
B.
,BennettJr,J.
P.
,AlteredintracellularsignalingandreducedviabilityofAlzheimer'sdiseaseneuronalcybridsisreproducedbyβ-amyloidpeptideactingthroughreceptorforadvancedglycationendproducts(RAGE).
MolCellNeurosci.
2005,29,333-343.
85.
Li,M.
,Sun,L.
,Luo,Y.
,Xie,C.
etal.
,High-mobilitygroupbox1releasedfromastrocytespromotestheproliferationofculturedneuralstem/progenitorcells.
IntJMolMed.
2014,34,705-714.
86.
Ohshiro,Y.
,Lee,Y.
,King,G.
L.
,Mechanismofdiabeticnephropathy:roleofproteinkinaseCactivation.
AdvStudMed.
2005,5,s11-s19.
87.
Geraldes,P.
,King,G.
L.
,ActivationofproteinkinaseCisoformsanditsimpactondiabeticcomplications.
CircRes.
2010,106,1319-1331.
88.
Sakaguchi,M.
,Murata,H.
,Yamamoto,K.
,Ono,T.
etal.
,TIRAP,anadaptorproteinforTLR2/4,transducesasignalfromRAGEphosphorylateduponligandbinding.
PLoSOne2011,6,e23132.
89.
Cassese,A.
,Esposito,I.
,Fiory,F.
,Barbagallo,A.
P.
etal.
,Inskeletalmuscleadvancedglycationendproducts(AGEs)inhibitinsulinactionandinducetheformationofmultimolecularcomplexesincludingthereceptorforAGEs.
JBiolChem.
2008,283,36088-36099.
90.
Warboys,C.
M.
,Toh,H.
B.
,Fraser,P.
A.
,RoleofNADPHoxidaseinretinalmicrovascularpermeabilityincreasebyRAGEactivation.
InvestOphthalmol.
Vis.
Sci.
2009,50,1319-1328.
www.
proteomics-journal.
comPage36ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
91.
Thallas-Bonke,V.
,Thorpe,S.
R.
,Coughlan,M.
T.
,Fukami,K.
etal.
,InhibitionofNADPHoxidasepreventsadvancedglycationendproduct-mediateddamageindiabeticnephropathythroughaproteinkinaseC-alphadependentpathway.
Diabetes2008,57,460-469.
92.
Yamagishi,S.
I.
,Commenton:Thallas-Bonkeetal.
(2008)InhibitionofNADPHOxidasePreventsAdvancedGlycationEndProduct–MediatedDamageinDiabeticNephropathyThroughaProteinKinaseC-α–DependentPathway:Diabetes57:460–469,2008.
Diabetes2008,57,e13-e13.
93.
Yu,L.
,Zhao,Y.
,Xu,S.
,Ding,F.
etal.
,AdvancedGlycationEndProduct(AGE)-AGEReceptor(RAGE)SystemUpregulatedConnexin43ExpressioninRatCardiomyocytesviaPKCandErkMAPKPathways.
IntJMolSci.
2013,14,2242-2257.
94.
Ding,Y.
,Kantarci,A.
,Badwey,J.
A.
,Hasturk,H.
,Malabanan,A.
,VanDyke,T.
E.
,Phosphorylationofpleckstrinincreasesproinflammatorycytokinesecretionbymononuclearphagocytesindiabetesmellitus.
JImmunol.
2007,179,647-654.
95.
Shanmugam,N.
,Kim,Y.
S.
,Lanting,L.
,Natarajan,R.
,Regulationofcyclooxygenase-2expressionbyligationofthereceptorofadvancedglycationendproducts.
J.
Biol.
Chem.
2003,278,34834-3484.
96.
Downs,C.
A.
,Kreiner,L.
H.
,Johnson,N.
M.
,Brown,L.
A.
,Helms,M.
N.
,RAGERegulatesLungFluidBalanceviaPKC-gp91phoxSignalingtoEpithelialSodiumChannels(ENaC).
AmJRespirCellMolBiol.
2014.
97.
Foster,F.
M.
,Traer,C.
J.
,Abraham,S.
M.
,Fry,M.
J.
,Thephosphoinositide(PI)3-kinasefamily.
JCellSci.
2003,116,3037-3040.
www.
proteomics-journal.
comPage37ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
98.
Cantley,L.
C.
,Thephosphoinositide3-kinasepathway.
Science2002,296,1655-1657.
99.
Yang,S.
J.
,Chen,C.
Y.
,Chang,G.
D.
,Wen,H.
C.
etal.
,ActivationofAktbyadvancedglycationendproducts(AGEs):involvementofIGF-1receptorandcaveolin-1.
PloSone2013,8,e58100.
100.
Xu,D.
,Kyriakis,J.
M.
,Phosphatidylinositol3′-kinase-dependentactivationofrenalmesangialcellKi-RasandERKbyadvancedglycationendproducts.
JBiolChem.
2003,278,39349-39355.
101.
Li,J.
J.
,Kwak,S.
J.
,Jung,D.
S.
,Kim,J.
J.
etal.
,Podocytebiologyindiabeticnephropathy.
KidneyIntSuppl.
2007,72,S36-S42.
102.
DallaVestra,M.
,Masiero,A.
,Roiter,A.
M.
,Saller,A.
etal.
,IspodocyteinjuryrelevantindiabeticnephropathyStudiesinpatientswithtype2diabetes.
Diabetes2003,52,1031-1035.
103.
Cheng,C.
L.
,Tang,Y.
,Zheng,Z.
,Liu,X.
etal.
,Advancedglycationend-productsactivatetherenin-angiotensinsystemthroughtheRAGE/PI3-Ksignalingpathwayinpodocytes.
ClinInvestMed.
2012,35,E282-E293.
104.
Touré,F.
,Zahm,J.
,Garnotel,R.
,Lambert,E.
etal.
,Receptorforadvancedglycationend-products(RAGE)modulatesneutrophiladhesionandmigrationonglycoxidatedextracellularmatrix.
Biochem.
J.
2008,416,255-261.
105.
Hu,P.
,Lai,D.
,Lu,P.
,Gao,J.
,He,H.
,ERKandAktsignalingpathwaysareinvolvedinadvancedglycationendproduct-inducedautophagyinratvascularsmoothmusclecells.
IntJMolMed.
2012,29,613-618.
www.
proteomics-journal.
comPage38ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
106.
Haraba,R.
,Suica,V.
I.
,Uyy,E.
,Ivan,L.
,Antohe,F.
,Hyperlipidemiastimulatestheextracellularreleaseofthenuclearhighmobilitygroupbox1protein.
CellTissueRes.
2011,346,361-368.
107.
Shiu,S.
W.
,Wong,Y.
,Tan,K.
C.
,Effectofadvancedglycationendproductsonlectin-likeoxidizedlowdensitylipoproteinreceptor-1expressioninendothelialcells.
JAtherosclerThromb.
2011,19,1083-1092.
108.
Qin,Q.
,Niu,J.
,Wang,Z.
,Xu,W.
etal.
,Heparanaseinducedbyadvancedglycationendproducts(AGEs)promotesmacrophagemigrationinvolvingRAGEandPI3K/AKTpathway.
CardiovascDiabetol.
2013,12,37.
109.
Vincent,A.
M.
,Perrone,L.
,Sullivan,K.
A.
,Backus,C.
etal.
,Receptorforadvancedglycationendproductsactivationinjuresprimarysensoryneuronsviaoxidativestress.
Endocrinology2007,148,548-558.
110.
Tafani,M.
,Schito,L.
,Pellegrini,L.
,Villanova,L.
etal.
,Hypoxia-increasedRAGEandP2X7RexpressionregulatestumorcellinvasionthroughphosphorylationofErk1/2andAktandnucleartranslocationofNF-κB.
Carcinogenesis2011,32,1167-1175.
111.
Kuniyasu,H.
,Chihara,Y.
,Kondo,H.
,Differentialeffectsbetweenamphoterinandadvancedglycationendproductsoncoloncancercells.
IntJCancer2003,104,722-727.
112.
Xu,X.
,Zhu,H.
,Wang,T.
,Sun,Y.
etal.
,ExogenousHigh‐MobilityGroupBox1InhibitsApoptosisandPromotestheProliferationofLewisCellsviaRAGE/TLR4‐DependentSignalPathways.
ScandJImmunol.
2014,79,386-394.
www.
proteomics-journal.
comPage39ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
113.
Cai,W.
,He,J.
C.
,Zhu,L.
,Chen,X.
etal.
,AGE-receptor-1counteractscellularoxidantstressinducedbyAGEsvianegativeregulationofp66shc-dependentFKHRL1phosphorylation.
AmJPhysiolCellPhysiol.
2008,294,C145-C152.
114.
Sakaguchi,M.
,Murata,H.
,Aoyama,Y.
,Hibino,T.
etal.
,DNAX-ActivatingProtein10(DAP10)MembraneAdaptorAssociateswithReceptorforAdvancedGlycationEndProducts(RAGE)andModulatestheRAGE-triggeredSignalingPathwayinHumanKeratinocytes.
JBiolChem.
2014,jbc-M114.
115.
Rawlings,J.
S.
,Rosler,K.
M.
,Harrison,D.
A.
,TheJAK/STATsignalingpathway.
JCellSci.
2004,117,1281-1283.
116.
Bierhaus,A.
,Humpert,P.
M.
,Morcos,M.
,Wendt,T.
etal.
,UnderstandingRAGE,thereceptorforadvancedglycationendproducts.
JMolMed.
2005,83,876-886.
117.
Rojas,A.
,Figueroa,H.
,Morales,E.
,Fuelinginflammationattumormicroenvironment:theroleofmultiligand/RAGEaxis.
Carcinogenesis2010,31,334-341.
118.
Schindler,C.
W.
,SeriesIntroduction.
JAK-STATsignalinginhumandisease.
JClinInvest.
2002,109,1133-1137.
119.
Constantinescu,S.
N.
,Girardot,M.
,Pecquet,C.
,MiningforJAK-STATmutationsincancer.
TrendsBiochemSci.
2008,33,122-131.
120.
Tang,J.
Z.
,Zuo,Z.
H.
,Kong,X.
J.
,Steiner,M.
etal.
,Signaltransducerandactivatoroftranscription(STAT)-5AandSTAT5Bdifferentiallyregulatehumanmammarycarcinomacellbehaviour.
Endocrinology2010,151,43-55.
121.
Kim,J.
Y.
,Park,H.
K.
,Yoon,J.
S.
,Kim,S.
J.
etal.
,Advancedglycationendproduct(AGE)-inducedproliferationofHELcellsviareceptorforAGE-relatedsignalpathways.
IntJOncol.
2008,33,493-501.
www.
proteomics-journal.
comPage40ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
122.
Sansone,P.
,Bromberg,J.
,Targetingtheinterleukin-6/Jak/statPathwayinhumanmalignancies.
JClinOncol.
2012,30,1005-1014.
123.
Bromberg,J.
,Statproteinsandoncogenesis.
JClinInvest.
2002,109,1139-1142.
124.
O'Shea,J.
J.
,Gadina,M.
,Schreiber,R.
D.
,CytokineSignalingin2002:NewSurprisesintheJak/StatPathway.
Cell2002,109,S121-S131.
125.
Kang,R.
,Tang,D.
,Lotze,M.
T.
,Zeh,H.
J.
,AGER/RAGE-mediatedautophagypromotespancreatictumorigenesisandbioenergeticsthroughtheIL6-pSTAT3pathway.
Autophagy2012,8,989-91.
126.
Kang,R.
,Loux,T.
,Tang,D.
,Schapiro,N.
E.
etal.
,Theexpressionofthereceptorforadvancedglycationendproducts(RAGE)ispermissiveforearlypancreaticneoplasia.
ProcNatlAcadSciUSA.
2012,109,7031-7036.
127.
Chen,S.
C.
,Guh,J.
Y.
,Chen,H.
C.
,Yang,Y.
L.
etal.
,Advancedglycationend-product-inducedmitogenesisisdependentonJanuskinase2-inducedheatshockprotein70innormalratkidneyinterstitialfibroblastcells.
TranslRes.
2007,149,274-281.
128.
Zeng,S.
,Zhang,Q.
Y.
,Huang,J.
,Vedantham,S.
etal.
,OpposingrolesofRAGEandMyd88signalinginextensiveliverresection.
FASEBJ.
2012,26,882-893.
129.
Huang,J.
S.
,Guh,J.
Y.
,Chen,H.
C.
,Hung,W.
C.
etal.
,Roleofreceptorforadvancedglycationend-product(RAGE)andtheJAK/STAT-signalingpathwayinAGE-inducedcollagenproductioninNRK-49Fcells.
JCellBiochem.
2001,81,102-113.
130.
Yammani,R.
R.
,Long,D.
,Loeser,R.
F.
,Interleukin-7stimulatessecretionofS100A4byactivatingtheJAK/STATsignalingpathwayinhumanarticularchondrocytes.
ArthritisRheum.
2009,60,792-800.
www.
proteomics-journal.
comPage41ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
131.
Huang,C.
Y.
,Lai,K.
Y.
,Hung,L.
F.
,Wu,W.
L.
etal.
,AdvancedglycationendproductscausecollagenIIreductionbyactivatingJanuskinase/signaltransducerandactivatoroftranscription3pathwayinporcinechondrocytes.
Rheumatology2011,50,1379-1389.
132.
Saleh,A.
,Smith,D.
R.
,Tessler,L.
,Mateo,A.
R.
etal.
,Receptorforadvancedglycationend-products(RAGE)activatesdivergentsignalingpathwaystoaugmentneuriteoutgrowthofadultsensoryneurons.
ExpNeurol.
2013,249,149-159.
133.
Fiol,C.
J.
,Wang,A.
,Roeske,R.
W.
,Roach,P.
J.
,Orderedmultisiteproteinphosphorylation.
Analysisofglycogensynthasekinase3actionusingmodelpeptidesubstrates.
JBiolChem.
1990,265,6061-6065.
134.
Embi,N.
,RYLATT,D.
B.
,COHEN,P.
,GlycogenSynthaseKinase‐3fromRabbitSkeletalMuscle.
EurJBiochem.
1980,107,519-527.
135.
Ali,A.
,Hoeflich,K.
P.
,Woodgett,J.
R.
,Glycogensynthasekinase-3:properties,functions,andregulation.
ChemRev.
2001,101,2527-2540.
136.
Yao,H.
B.
,Shaw,P.
C.
,Wong,C.
C.
,Wan,D.
C.
C.
,Expressionofglycogensynthasekinase-3isoformsinmousetissuesandtheirtranscriptioninthebrain.
JChemNeuroanat.
2002,23,291-297.
137.
Hughes,K.
,Nikolakaki,E.
,Plyte,S.
E.
,Totty,N.
F.
,Woodgett,J.
R.
,Modulationoftheglycogensynthasekinase-3familybytyrosinephosphorylation.
EMBOJ.
1993,12,803-808.
138.
Sutherland,C.
,Leighton,I.
A.
,Cohen,P.
,Inactivationofglycogensynthasekinase-3betabyphosphorylation:newkinaseconnectionsininsulinandgrowth-factorsignaling.
BiochemJ.
1993,296,15-19.
www.
proteomics-journal.
comPage42ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
139.
Tanji,C.
,Yamamoto,H.
,Yorioka,N.
,Kohno,N.
etal.
,A-kinaseanchoringproteinAKAP220bindstoglycogensynthasekinase-3β(GSK-3β)andmediatesproteinkinaseA-dependentinhibitionofGSK-3β.
JBiolChem.
2002,277,36955-36961.
140.
Grimes,C.
A.
,Jope,R.
S.
,Themultifacetedrolesofglycogensynthasekinase3βincellularsignaling.
ProgNeurobiol.
2001,65,391-426.
141.
Doble,B.
W.
,Woodgett,J.
R.
,GSK-3:tricksofthetradeforamulti-taskingkinase.
JCellSci.
2003,116,1175-1186.
142.
Li,X.
H.
,Lv,B.
L.
,Xie,J.
Z.
,Liu,J.
etal.
,AGEsinduceAlzheimer-liketaupathologyandmemorydeficitviaRAGE-mediatedGSK-3activation.
NeurobiolAging2012,33,1400-1410.
143.
Chen,S.
,An,F.
M.
,Yin,L.
,Liu,A.
R.
etal.
,Glucagon-likepeptide-1protectshippocampalneuronsagainstadvancedglycationendproduct-inducedtauhyperphosphorylation.
Neuroscience2014,256,137-146.
144.
Li,X.
H.
,Du,L.
L.
,Cheng,X.
S.
,Jiang,X.
etal.
,Glycationexacerbatestheneuronaltoxicityofβ-amyloid.
CellDeathDis.
2013,4,e673.
145.
Li,X.
H.
,Xie,J.
Z.
,Jiang,X.
,Lv,B.
L.
etal.
,MethylglyoxalinducestauhyperphosphorylationviapromotingAGEsformation.
NeuromolecularMed.
2012,14,338-348.
146.
Qu,Z.
S.
,Li,L.
,Sun,X.
J.
,Zhao,Y.
W.
etal.
,GlycogenSynthaseKinase-3RegulatesProductionofAmyloid-βPeptidesandTauPhosphorylationinDiabeticRatBrain.
ScientificWorldJournal2014.
147.
Yu,W.
,Wu,J.
,Cai,F.
,Xiang,J.
etal.
,Curcuminalleviatesdiabeticcardiomyopathyinexperimentaldiabeticrats.
PloSone2012,7,e52013.
www.
proteomics-journal.
comPage43ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
148.
Chaves,M.
L.
,Camozzato,A.
L.
,Ferreira,E.
D.
,Piazenski,I.
etal.
,SerumlevelsofS100BandNSEproteinsinAlzheimer'sdiseasepatients.
JNeuroinflammation2010,7.
149.
Li,J.
P.
,Lu,L.
,Wang,L.
J.
,Zhang,F.
R.
,Shen,W.
F.
,IncreasedserumlevelsofS100Barerelatedtotheseverityofcardiacdysfunction,renalinsufficiencyandmajorcardiaceventsinpatientswithchronicheartfailure.
ClinBiochem.
2011,44,984-988.
150.
Sun,K.
K.
,Ji,C.
,Li,X.
,Zhang,L.
etal.
,OverexpressionofhighmobilitygroupproteinB1correlateswiththeproliferationandmetastasisoflungadenocarcinomacells.
MolMedRep.
2013,7,1678-1682.
151.
Arancio,O.
,Zhang,H.
P.
,Chen,X.
,Lin,C.
etal.
,RAGEpotentiatesAβ‐inducedperturbationofneuronalfunctionintransgenicmice.
EMBOJ.
2004,23,4096-4105.
152.
Aberle,H.
,Bauer,A.
,Stappert,J.
,Kispert,A.
,Kemler,R.
β‐cateninisatargetfortheubiquitin–proteasomepathway.
EMBOJ.
1997,16,3797-3804.
153.
Hornbeck,P.
V.
,Kornhauser,J.
M.
,Tkachev,S.
,Zhang,B.
etal.
,PhosphoSitePlus:acomprehensiveresourceforinvestigatingthestructureandfunctionofexperimentallydeterminedpost-translationalmodificationsinmanandmouse.
NucleicAcidsRes.
2012,40,D261-D270.
154.
Ghidoni,R.
,Paterlini,A.
,Benussi,L.
,TranslationalproteomicsinAlzheimer'sdiseaseandrelateddisorders.
ClinBiochem.
2013,46,480-486.
155.
Atochin,D.
N.
,Wang,A.
,Liu,V.
W.
,Critchlow,J.
D.
etal.
,ThephosphorylationstateofeNOSmodulatesvascularreactivityandoutcomeofcerebralischemiainvivo.
JClinInvest.
2007,117,1961-1967.
www.
proteomics-journal.
comPage44ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
156.
Liang,C.
,Ren,Y.
,Tan,H.
,He,Z.
etal.
,RosiglitazoneviaupregulationofAkt/eNOSpathwaysattenuatesdysfunctionofendothelialprogenitorcells,inducedbyadvancedglycationendproducts.
BrJPharmacol.
2009,158,1865-1873.
157.
Chen,J.
,Jin,J.
,Song,M.
,Dong,H.
etal.
,C-reactiveproteindown-regulatesendothelialnitricoxidesynthaseexpressionandpromotesapoptosisinendothelialprogenitorcellsthroughreceptorforadvancedglycationend-products.
Gene2012,496,128-135.
158.
Chung,J.
,Grammar,T.
C.
,Lemon,K.
P.
,Kazlauskas,A.
,Blenis,J.
,PDGF-andinsulin-dependentpp70S6kactivationmediatedbyphosphatidylinositol-3-OHkinase.
Nature1994,370,71-75.
159.
Chung,J.
,Kuo,C.
J.
,Crabtree,G.
R.
,lenis,J.
,Rapamycin-FKBPspecificallyblocksgrowth-dependentactivationofandsignalingbythe70kdS6proteinkinases.
Cell1992,69,1227-1236.
160.
Murakami,H.
,Henry,G.
,Leonil,J.
,Galmier,M.
J.
etal.
,Effectofachronicintakeofhighdietaryglycatedproteinsonskeletalmuscleproteinmetabolismduringaginginnormalrats.
FASEBJ.
2011,25,233-2.
161.
Shaw,R.
J.
,LKB1andAMP‐activatedproteinkinasecontrolofmTORsignallingandgrowth.
ActaPhysiol.
2009,196,65-80.
162.
Son,S.
M.
,Jung,E.
S.
,Shin,H.
J.
,Byun,J.
,Mook-Jung,I.
,Aβ-inducedformationofautophagosomesismediatedbyRAGE-CaMKKβ-AMPKsignaling.
NeurobiolAging2012,33,1006-e11.
163.
Catalano,S.
,Giordano,C.
,Rizza,P.
,Gu,G.
,Barone,I.
,EvidencethatleptinthroughSTATandCREBsignalingenhancescyclinD1expressionandpromoteshumanendometrialcancerproliferation.
JCellPhysiol.
2009,218,490-500.
www.
proteomics-journal.
comPage45ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
164.
Sugars,K.
L.
,Rubinsztein,D.
C.
,TranscriptionalabnormalitiesinHuntingtondisease.
TrendsGenet.
200319,233-238.
165.
Huttunen,H.
J.
,Kuja-Panula,J.
,Rauvala,H.
,Receptorforadvancedglycationendproducts(RAGE)signalinginducesCREB-dependentchromograninexpressionduringneuronaldifferentiation.
JCellBiochem.
2002,277,38635-38646.
166.
Aderem,A.
,Signaltransductionandtheactincytoskeleton:therolesofMARCKSandprofilin.
TrendsBiochemSci.
1992,17,438-443.
167.
Green,T.
D.
,Crews,A.
L.
,Park,J.
,Fang,S.
,Adler,K.
B.
,Regulationofmucinsecretionandinflammationinasthma:aroleforMARCKSproteinBiochimBiophysActa.
2011,1810,1110-1113.
www.
proteomics-journal.
comPage46ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Figure1.
SchematicrepresentationofRAGEanditsisoforms(esRAGE-endogenoussecretoryRAGE,sRAGE-solubleRAGE,flRAGE-fulllengthRAGE)www.
proteomics-journal.
comPage47ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Figure2.
DepictionofRAGEactivatedsignalingpathwayindifferentdiseases.
www.
proteomics-journal.
comPage48ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Figure3.
WorkflowusedtoanalyzethesubstratesofRAGEactivatedkinasesfromPhosphositeplusdatabasewww.
proteomics-journal.
comPage49ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Figure4.
TheplotdepictsassociationofphosphosubstrateswithdifferentdiseasesandRAGEactivatedkinases,andcolourcodeindicatesthediseaseassociation.
ForexampleERK2isasubstrateforERKandJNK,andisassociatedwithallfourdiseases.
Pleasecitethisarticleasdoi:10.
1002/pmic.
201400169.
Thisarticleisprotectedbycopyright.
Allrightsreserved.
InvestigationofPhosphoproteomeinRAGEsignalingKedarBBatkulwar,SnehaBBansode,GouriVPatil,RashmiKGodbole,RubinaSKazi,SubashchandraboseChinnathambi,DhanasekaranShanmugam,MaheshJKulkarni*ProteomicsFacility,DivisionofBiochemicalSciences,CSIR-NationalChemicalLaboratory,Pune-411008,India#Authors2to5haveequallycontributed*ForcorrespondenceMaheshJKulkarniProteomicsFacility,DivisionofBiochemicalSciences,CSIR-NationalChemicalLaboratory,Pune-411008,IndiaE-mail:mj.
kulkarni@ncl.
res.
inPhone:+912025902541www.
proteomics-journal.
comPage2ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
AbstractReceptorforAdvancedGlycationEndproducts(RAGE)isoneofthemostimportantfactorsimplicatedindiabetes,cardiovasculardiseases,neurodegenerativediseases,andcancer.
Itisapatternrecognitionreceptor,byvirtueofitsabilitytointeractwithmultipleligands;RAGEactivatesseveralsignaltransductionpathwaysthroughinvolvementofvariouskinases,whichphosphorylatetheirrespectivesubstrates.
OnlyfewsubstrateshavebeenknowntobephosphorylatedinresponsetoactivationbyRAGE(e.
g.
NF-кB),howeveritispossiblethatthesekinasescanphosphorylatemultiplesubstratesdependingupontheirexpressionandlocalization,leadingtoalteredcellularresponsesindifferentcelltypesandconditions.
Onesuchexample,Glycogensynthasekinase3beta(GSK3β)whichisknowntophosphorylateglycogensynthase,actsdownstreamtoRAGEandhyperphosphorylatesMicrotubuleAssociatedProteinTau(MAPT)causingneuronaldamage.
Thus,itisimportanttounderstandtheroleofvariousRAGEactivatedkinasesandtheirsubstrates.
Therefore,wehavereviewedherethedetailsofRAGEactivatedkinasesinresponsetodifferentligandsandtheirrespectivephosphoproteome.
Further,wediscusstheanalysisofthedataminedforknownsubstratesofthesekinasesfromPhosphoSitePlus(http://www.
phosphosite.
org)database,andtheroleofsomeoftheimportantsubstratesinvolvedincancer,diabetes,cardiovasculardiseasesandneurodegenerativediseases.
Insummary,thisreviewprovidesinformationonRAGEactivatedkinasesandtheirphosphoproteome,whichwillbehelpfulinunderstandingthepossibleroleofRAGEanditsligandsinprogressionofdiseases.
Keywords:disease,glycation,kinase,phosphoproteome,RAGEwww.
proteomics-journal.
comPage3ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Abbreviations1.
RAGE-Receptorforadvancedglycationendproducts2.
AGEs-Advancedglycationendproducts3.
NF-кB-NuclearfactorkappaB4.
GSK3β-Glycogensynthasekinase-3beta5.
HMGB1-Highmobilitygroupboxprotein16.
Aβ-Amyloidbeta7.
SP1-Specificityprotein18.
MAPK-Mitogenactivatedproteinkinase9.
PKC-ProteinkinaseC10.
JNK-c-JunN-terminalKinase11.
JAK-Janusactivatedkinase12.
ERK-Extracellularsignalregulatedkinase13.
iNOS-induciblenitricoxidesynthase14.
eNOS-Endothelialnitricoxidesynthase15.
mTOR-Mammaliantargetofrapamycin16.
MMP-Matrixmetalloproteinase17.
STAT-Signaltransducersandactivatorsoftranscription18.
TNFα-Tumornecrosisfactoralphawww.
proteomics-journal.
comPage4ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
IntroductionRAGE,amulti-ligandpatternrecognitionreceptor,belongstotheimmunoglobulinsuperfamilyofproteins.
Althoughitwasfirstdescribedasareceptorforadvancedglycationendproducts(AGEs),itinteractswithdiverseclassesofligandincludingS100B,highmobilitygroupboxprotein(HMGB)-1,Mac-1,amyloidbeta(Aβ)peptideandbetasheetfibrils[1].
TheRAGEmRNAandmatureproteincanundergoalternativesplicingandproteolysis,respectivelyresultinginmanyvariants.
FulllengthRAGE(flRAGE)consistsof5domains,threeextracellular,asingletransmembranedomainandasmallcytosolictail.
ExtracellulardomainsarelabeledasV,C1andC2domain[2].
WhilethesolubleRAGE(sRAGE)lackstransmembraneandcytosolicdomain,andthereforeitisneitheranchoredincellmembranenorabletoactivatesignalingpathwayslikeflRAGE(Fig1)[3].
ThesRAGEinteractswithRAGEligandsandhelpsintheclearanceofthesefromthesystem[4].
TheinitiationofRAGEsignalingcommonlyoccursthroughproductionofROSviaNADPHoxidaseandactivationofvariouskinasesincludingMAPKlikeERK1/2,SAPK/JNK,p38;PI3K/Akt;JAK/STAT;GSK3βetc[5].
ThesekinasestriggerseveraldownstreamsignalingcascadesoftenviaactivationoftranscriptionfactorsNF-кBandSP1,therebymodulatingspecificcellularprocesses[6].
Becauseofitsroleinoxidativestressandinflammation,RAGEisimplicatedinseveraldiseases.
Indiabetes,AGEsarethepredominantligandsthatinteractwithRAGEandcontributetothepathogenesisofdiabeticcomplicationslikeglomerulopathy,atherosclerosis,vasculopathyetc[7].
AGEsareagroupofheterogeneousmoleculesformedasaresultofaseriesofnon-enzymaticreactionsbetweenreducingsugarsandproteins[8,9].
SomeofthepredominantAGEsincludecarboxymethyllysine(CML),carboxyethyllysinewww.
proteomics-journal.
comPage5ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
(CEL),pentosidineetc[10-12].
AGEformationisacceleratedindiabetesduetohyperglycemiccondition[13].
AccumulationofAGEsleadsto(i)alteredproteinstructureandfunction[14],(ii)proteinaggregation,fibrilformation,andproteaseresistance[15,16],(iii)dysfunctionofextracellularmatrix[14]and(iv)RAGEactivationanddownstreamsignaling[17].
AGE/RAGEinteractioncontributessubstantiallytotheprogressionofdiabeticcomplications,cardiovasculardiseases,acceleratedaging,neurodegenerativediseases,andcancer[18].
WhereasinAlzheimer'sdisease,AβistheprincipalligandthatinteractswithRAGE[19].
Itconsistsof36-43aminoacidresiduesproducedasaresultofsequentialcleavageofAPPbybetaandgammasecretases[20].
RAGEmediatedAβinternalizationresultsinmitochondrialdysfunctioncausingcellulardamage[21].
Whileincancer,S100B,acalciumbindingproteinisknowntobeinvolvedinRAGEactivationbydimerization[22].
S100B/RAGEinteractionelicitsvariouscellulareffectsincludingcellsurvivalandcellproliferation[23,24].
HMGB1alsocalledasHMG1oramphoterinisasmallnon-histonechromatinproteinwhichinteractswithRAGEandactivatesproinflammatorypathways[25,26].
BlockadeofRAGEmediatedsignalingsuppressestumorgrowthandmetastasis,suggestingitspotentialinvolvementincancer[27].
Hence,RAGEplaysanimportantroleintheregulationofvitalcellprocessessuchasapoptosis,autophagy,inflammationandproliferation.
TheinvolvementofRAGEindifferentdiseaseshasbeenreviewedingreatdetailearlier[28-31].
However,thedetailedmolecularmechanismofRAGEsignaling,especiallytheinvolvementofvariouskinasesandtheirsubstratesisnotwellunderstood.
MostofthereportedstudiesonRAGEsignalingfocusonactivationofspecifickinaseslikeMAPKs,PI3K,PKCandGSK3β.
ThephosphorylationofonlyfewimportantsubstratessuchasNF-кB,FOXO,andSP1isknown;butglobalphosphoproteomeinRAGEactivatedsignalingispoorlystudied.
Itisimportanttoidentifywww.
proteomics-journal.
comPage6ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
thefactorsthatfacilitatetherecruitmentofspecifickinases,alongwiththeirphosphoproteome,whichcangreatlyenhanceourunderstandingofthecellularresponsestoRAGEsignaling.
Therefore,inthisreviewwehavediscussedindetailaboutRAGEactivatedkinasesandtheirphosphoproteomeinresponsetodifferentligands.
RAGEactivatedkinasesOnceactivatedbyitsligands,RAGEcanrecruitseveralkinasesandtriggeracascadeofsignalingevents(Fig2).
SomeoftheimportantkinasesthatareactivateddownstreamtoRAGEaredescribedbelow.
Mitogenactivatedproteinkinases(MAPK)MAPKsareserine/threonineproteinkinasesthattranslateexternalstimuliintoawiderangeofcellularresponses.
ConventionalMAPKsincludeextracellularsignal-regulatedkinases1/2(ERK1/2),c-JunN-terminalkinases1/2/3(JNK1/2/3),p38isoforms(α,β,γ,andδ),andERK5.
AtypicalMAPKsincludeERK3/4,ERK7,andNemo-likekinase(NLK).
EachgroupofconventionalMAPKsiscomposedofasetofthreeevolutionarilyconserved,sequentiallyactingkinases:aMAPK,aMAPKkinase(MAPKK),andaMAPKKkinase(MAPKKK)[32].
SeveralligandsinteractwithRAGEandactivatedifferentMAPKsdependingoncelltypeanddisease.
p38MAPKisoneofthekeydownstreameffectorofRAGEthatmediatesactivationofNF-кBdependentsecretionofcytokinesresultinginacceleratedinflammation.
AGEinducedNF-кBactivationresultsinincreasedsecretionofTumorNecrosisFactoralpha(TNF-α),interleukin-1beta(IL-1β),andMonocytesChemoattractantProtein(MCP)-1.
RAGEhasbeenimplicatedintheformationofproinflammatorymilieu,whichplaysanimportantroleintheadvancementofseveraldisorders[33].
www.
proteomics-journal.
comPage7ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Indiabetes,AGEinducedp38MAPKactivationresultsinincreasedalkalinephosphataseactivityviaRAGE,whichin-turninducesosteoblastlikedifferentiationofsmoothmusclecellsresultinginvascularcalcification[34].
Indiabeticnephropathy(DN),AGE/RAGEinteractionactivatesp38MAPKthatphosphorylatesATF2,atranscriptionfactorwhichinducesiNOSexpressioninmesangialcells[35].
Inpodocytes,thisinteractionresultsinactivationoftranscriptionfactorFOXO4viap38MAPK,leadingtoapoptosis[36].
AGEscaninduceexpressionofconnectivetissuegrowthfactor(CTGF),whichpromotestheadvancementofdiabeticnephropathy.
TheexpressionofCTGFdependsonRAGE-ERK/p38-Smad3cross-talkpathway[37].
AGEsalsoactivateERK1/2andEGFRviaRAGEmediatedROSinNRF-49Fkidneycells,whichareimplicatedindiabeticnephropathy[38].
AGEsactivateSmad2/3inhumanvasculopathyviaRAGEandERK/p38MAPKs[39].
AGEdepositionandRAGEexpressioncontributestotheprogressionofatherosclerosisviaERKactivationindiabeticandhypertensivemice[40].
OligomerizationofRAGEisbelievedtoplayanessentialroleinsignaltransduction.
HeparansulfateinduceshexamerizationoftheextracellularVandC1domainspresentinRAGE,resultinginactivationofERK1/2inendothelialcells[41].
S100A4demonstratedRAGEmediatedERKphoshorylationandMMP2activation,whichincreasescellmotilityinhumanpulmonaryarterysmoothmusclecells(hPASMCs)contributingtoprogressionofvasculardiseases[42].
AGEshavebeenalsoimplicatedindiabetickeratopathy,whereintheyinduceapoptosisincornealepithelialcellsbyROSgenerationthroughactivationofNADPHoxidase,JNKandp38MAPKinRAGEdependentmanner[43].
Incancer,bindingofamphoterintoRAGEenhancestheRAGEboundERK1/2activity,asdemonstratedbyphosphorylationofmyelinbasicprotein,anERKsubstrate[44].
Inwww.
proteomics-journal.
comPage8ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
myoblastsandrhabdomyosarcomacells,HMGB1/RAGEinteractionpreventstumorformationwhereasingliomasitfacilitatesmetastasisensuingininvasivephenotype.
RAGEactivationbyHMGB1leadstomyogenesisviaCdc42-Rac-MKK6-p38MAPKpathwayandisassociatedwithinhibitionofproliferation.
FunctionalinactivationofRAGEcausesdecreaseinactivityofp21Waf1,aproliferationinhibitorandincreaseinactivityofcyclinD1,retinoblastomaprotein,ERK1/2andJNK[45,46].
WhereasinC6gliomacells,RAGE/amphoterininteractionplaysanimportantroleintumorproliferation,invasionandmatrixmetalloproteinase(MMP)expression,whileblockingthisinteractiondecreasesgrowthandmetastasisbyinactivationofp44/p42,p38andJNKMAPKpathway[47].
S100A8/A9ligandhasadualroleintumorbiology;stimulatespseudopodiaformationandexertsgrowthpromotingactivity.
ItpromotescellgrowthinaRAGEdependentmannerbyphosphorylationofp38andp44/42MAPKsandactivationofNF-кB[48].
p38MAPKcausesactivationofMAPKAPkinase-2(MK2),Hsp27andSAPK/JNKpathways.
SimilarresponseswereobservedinHaCaTkeratinocytesandclearcellrenalcellcarcinoma(CCRCC)viaERK1/2activationinRAGEdependentmanner[49].
HMGB1/RAGEinteractionplaysanimportantroleintheadvancementofliverfibrosis.
RAGEmediatedHMGB1internalizationcausesSmad2andERK1/2phosphorylationresultingincollagentypeIandα-smoothmuscleactin(α-SMA)expression,andproliferationofhepaticstellatecellsleadingtothefibroticchangesofliver[50].
S100P,aRAGEligandthatisexpressedathighlevelsincoloncancer,interactswithRAGEandstimulatesgrowth,migration,ERKphosphorylation,andNF-кBactivationincancercells[51].
VEGFincardiacmyocytesrespondtoS100BandRAGEinteractioninducedERK1/2phosphorylationandmyofibroblastproliferationcontributingtomyocardialinfarction[52].
RAGE/S100BinteractionaftermyocardialinfarctionstimulatesmyocyteapoptosisbyactivatingERK1/2andp53signaling[53].
www.
proteomics-journal.
comPage9ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Incorticalneurons,RAGEmediatesAβuptakeacrossthecellmembraneviaactivationofp38MAPK.
p38MAPKactivationcausesinflammatoryresponseandisimplicatedinthepathogenesisofAlzheimer'sdisease[54].
Aβimpairslong-termpotentiation(LTP)intheentorhinalcortexthroughRAGE-mediatedp38MAPKactivation[55].
Bloodbrainbarrier(BBB)ismainlycomposedofcerebralendothelialcells(CEC)andastrocytes.
BBBdamageisassociatedwithseveraldisordersincludingAlzheimer'sdisease.
AβinducedERK1/2andcytosolicphosphorilaseA2phosphorylationinCECsandastrocytesoccursviaRAGEdependentpathwayscausingcellulardamage[56].
APP/PS1transgenicalzheimermousemodeldemonstratesstrongincreaseintheexpressionofmouseformylpeptidereceptor(mFPR1/2)andRAGEexpressioninthecortexandhippocampus.
FPRandRAGEplayanimportantroleinAβ,S100BandAGE-BSAinducedERK1/2phosphorylationandregulationofcAMPlevelinglialcells[57].
AβstimulatedtransientactivationofERK1/2,p38MAPKinmicrogliaandmonocytesresultsinphosphorylationofcAMPresponseelement-binding(CREB)protein,providingamechanismforAβinducedchangesingeneexpression[58].
Inaddition,RAGEviaMAPKisalsoinvolvedinasthma,chronicobstructivepulmonarydisease,cysticfibrosis,andrheumatoidarthritis.
S100A12/RAGEinteractioninducesoverexpressionofMUC5AC,amajormucinprotein,viaERKactivationinairwayepithelialcells[59].
RAGEinducesdamageinhumanpulmonaryarteryendothelialcellsviap38MAPKactivationandHsp27phosphorylationresultinginacutelunginjury[60].
HMGB1inpresenceofLPSupregulatesTLR4andRAGEexpressioninSynovialfibroblasts,whichinducesp38MAPKandNF-кBactivationresultingintheproductionofIL-6,MMP3,andMMP13inrheumatoidarthritis[61,62].
www.
proteomics-journal.
comPage10ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
c-JunN-terminalkinase(JNK)JNKisaprolinedirectedSerine/Threoninekinase,andbelongstotheMAPKfamilywhichintegrateavarietyofenvironmentalstimulitocauseproliferation,differentiation,development,inflammatoryresponseandapoptosis.
TheJNKarmofMAPKsisactivatedinconditionsofintracellularandenvironmentalstress;henceitisalternativelyreferredtoasStressActivatedProteinKinase(SAPK).
JNKisactivatedbydualphosphorylationataspecificthreonineandtyrosineresiduesbyMKK4andMKK7kinases(MAPKKs)[63].
JNKactivationcausesphosphorylationoftranscriptionfactorsaswellasotherproteinstoeitheractivateorinactivatethem.
Itpredominantlyphosphorylatesandactivatesthec-JunandATF-2transcriptionfactorsoftheAP1complex.
JNKisactivatedbyERstress,osmoticstress,redoxstress,UVradiation,thecytokinesTNF-αandIL-1β,microbialinvasion,aswellasbyRAGEactivationinmanypathologicalconditions[64].
Inosteoarthritis,accumulationofAGEsinthearticularcartilagechondrocyteshasbeenassociatedwithinflammation.
AGEinducesexpressionofCOX-2andmicrosomalprostaglandinEsynthase-1byRAGEmediatedactivationofJNK,whichleadstoincreasedproductionofProstaglandinE2,amediatorofinflammation[65].
AGEBSAinducessecretionofMMP1,3and13inhumanosteoarthriticchondrocytesbyinteractionwithRAGEandactivationofJNKandp38kinase[66].
Onthecontrary,AGE/RAGEsignalingintheinfiltratingmacrophagescausesphosphorylationofp44/42ERK,p38,JNK,andPI3KandinhibitionofJNKactivitysignificantlyenhancedthestimulatoryeffectofAGEonMMP9secretion[67].
InDiabeticcondition,osteoblastapoptosisanddeficientbonemarrowformationismediatedbyAGE/RAGEinteraction.
CMLmodifiedcollagenactivatesp38,JNK,andtranscriptionfactor,FOXO1,leadingtoincreasedcaspase-3activityinfibroblastsresultinginwww.
proteomics-journal.
comPage11ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
apoptosis,whileinhibitionofthesekinasesreducedCML-collagenstimulatedapoptosis[68].
InMyocardialinfarction,stroke,andvenousthromboembolism,AGE/RAGEinteractioncausestranslocationofPKC-β2thatphosphorylatesJNKleadingtoincreasedtranscriptionofearlygrowthresponse-1(EGR-1).
EGR-1playsanintegralroleinthebiologicalresponsetohypoxia,itsupregulationmediatesincreasedexpressionofinflammatoryandprothromboticgenesresultingincellulardamage[69].
RAGEactivationbyAGEs,S100andHMGB1hasbeenimplicatedinischemicrepurfusion(I/R)injuryofcardiomyocytes.
CML/RAGEinteractionduringI/RincardiomyocytesandmousemodelcausesactivationofJNKandSTAT3leadingtocellulardamage[70,71].
LigationoftheheterodimericS100A8/A9proteinwithRAGEinpostischemiccardiacfibroblastsincreasedNF-кBnucleartranslocationandexpressionofpro-inflammatorycytokinesbyactivationofallthethreearmsofMAPKsnamelyERK,JNKandp38[72].
WhereasinteractionofHMGB1withRAGEinI/RcausedtheactivationofERK1/2andJNKMAPKsonly,alongwithincreasednucleartranslocationofNF-кB[73].
SimilareffectsofHMGB1/RAGEinteractionwereobservedinadiabeticmousemodelexposedtoI/R,thereforeimplyingaroleofHMGB1inheartfailureasacomplicationofdiabetes[74].
InconditionsofhepaticI/R,apartfromactivationofthesekinases,RAGEsignalingalsoactivatesc-Jun,leadingtoincreasedserumalanineaminotransferaseactivity,TNF-αandIL-6levels,NF-кBactivityandultimatelyapoptosisinlivertissue[75].
HMGB1functionsasacytokineandactivatesmacrophagesbybindingtoRAGE.
ThisinteractioncausesincreasedexpressionofMHCclassII,TNF-αandNObyactivationofp38,p42/44,JNKMAPKsalongwithincreasednucleartranslocationofNF-кB[76].
JNKactivationviaRAGEisimplicatedinpathogenesisofatherosclerosis.
S100B,andoxidizedLDLs(oxLDL)containingAGEepitopesinteractwithRAGEinaorticendothelialcellstocauseactivationofJNKleadingtoincreasedMMP2activityandupregulationofVCAM-1www.
proteomics-journal.
comPage12ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
whichmarksthesitesdestinedfordevelopmentofatheroscleroticplaques[77].
AGE/RAGEinteractionisreportedtocauseactivationofJNK,whichfurtherphosphorylatesAP1transcriptionfactortocauseexpressionofE-selectin,whichmediatesatherosclerosis[78].
Inendothelialcells,HMGB1stimulatesproinflammatoryresponsebyupregulationofleukocyteadhesionmoleculesnamelyICAM-1,VCAM-1,TNFα,IL-8,MCP-1,plasminogenactivatorinhibitor1(PAI-1),andtissueplasminogenactivator(tPA).
ThiseffectwasobservedbytransientphosphorylationoftheERK,p38andJNKandincreasednucleartranslocationofNF-кB[79].
S100B/RAGEInteractioninBV-2microglialcellsshowedupregulationofCOX2,IL-1βandTNF-αexpressionandchemokinesecretionbyparallelactivationofCdc42-Rac1-JNK-AP1andRas-Rac1-NF-кBpathwayscausinginflammation[80-82].
S100B/RAGEinteractioninamixedpopulationofneuronalandglialcellscausedactivationofJNK,whichspecificallyphosphorylatedthec-Jun.
c-JuncausestheexpressionofDickopff-1whichdisruptsWntsignalingpathwayandactivatesGSK3βresultinginTauhyperphosphorylation,apathologicalhallmarkofAD[83].
AβcausesapoptosisthroughinteractionwithRAGEviaactivationofp38,JNKpathways,andNF-кB,thusimplyingtheroleofRAGEinneurodegenerationincybridsofsporadicAD[84].
Incaseofbraininjury,HMGB1releasedfromreactiveastrocytescausesproliferationofneuronalstemandprogenitorcellsbyinteractionwithRAGEanddownstreamJNKphosphorylation[85].
ProteinkinaseC(PKC)ProteinkinaseC,amultifunctionalprotein,belongstoafamilyofserinethreoninekinase.
Thereare11isoformsofPKCandthesecanbeclassifiedintothreesubgroups;1)conventionalPKCs(α,β1,β2,andγ)thatareCa2+dependentandactivatedbywww.
proteomics-journal.
comPage13ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
phosphatidylserineandthesecondmessengerdiacylglycerol(DAG);2)novelPKCs(δ,η,ε,β)thatareCa2+independentandstimulatedbyDAG;and3)atypicalPKCs(ζandι/λ),neitherofwhichareknowntorequirecalciumorDAGforactivation.
AllPKCsappeartobephosphorylatedbyphosphoinositide-dependentproteinkinase1(PDPK1)[86].
PKChasbeenassociatedwithvascularalterationssuchasincreasedpermeability,contractility,extracellularmatrixsynthesis,cellgrowth,apoptosis,angiogenesis,leukocyteadhesionandcytokineactivation[87].
PKCphosphorylatescytoplasmicdomainofRAGEuponbindingofligands.
TIRAPandMyD88functionasadaptorproteinsforRAGEwhichfurtherrecruitIRAK4,whichinturnactivatesdownstreamkinasessuchasAkt,p38,JNKandIKKs.
Thisultimatelyleadstoproductionofinflammatorycytokines,IL-6,IL-8,andTNFαthroughactivationofNF-кB,whichregulatesinflammation,immuneresponseandothercellularfunctions[88].
PKCactivationinRAGEsignalingplaysanimportantroleintheprogressionofdiseasessuchasdiabetesandAlzheimer's.
ImpairmentofinsulinactioninthemuscleismediatedthroughtheformationofamultimolecularcomplexincludingAGE/RAGE/IRS-1/SrcandPKCα.
ThiscomplexrequiresIRS-1,oneofthekeymoleculesininsulinsignaling.
PKCmediatestheformationofthiscomplex,whichinhibitinsulinactioninskeletalmusclecellsinvitroand,possibly,invivo[89].
AGE/RAGEinteractioncausesPKCmediatedNADPHoxidaseactivationthatleadstoincreasedretinalcapillarypermeability,whichisassociatedwithinitialstagesofmacularedemaanddiabeticretinopathy[90].
AGEassociatedRAGEexpressionisfoundtobesignificantlyincreasedincaseofdiabeticnephropathy.
NADPHoxidase,whichplaysanimportantroleinROSproduction,isactivatedbyAGE/RAGEinteraction.
ThesubunitsofNADPHoxidaseincludingp47phox,nox4andp22phoxarewww.
proteomics-journal.
comPage14ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
activatedbyPKCαmediatedphosphorylation,whichfurtherresultsinoxidativestress[91].
Conversely,AGE/RAGEmediatedROSproductionactivatesPKC,whichphosphorylatesp47phoxthatinturnactivatesNADPHoxidase,resultinginapositivefeedbackloopforROSproduction[92].
AGE/RAGEinteractionelevatesCx43expression,themajorconnexininventricularcardiomyocytesthroughactivationofPKCandERKpathway.
ItalsoenhancesCx43redistributioninvivo,whichmaycontributetothearrhythmiasindiabetes[93].
Indiabeticmononuclearphagocytes,RAGEactivatesPKCwhichphosphorylatespleckstrin,acriticalmoleculeforproinflammatorycytokinesecretion[94].
AGEscanaugmentinflammatoryresponsesbyup-regulatingCOX-2viaRAGEandPKC,ERKandp38MAPKsignalingpathways,therebyleadingtomonocyteactivationandvascularcelldysfunction[95].
Inneuronalcells,AGE/RAGEinteractioncausesapoptosisduetoactivationofNADPHoxidase,PKCdeltaandredoxsensitivetranscriptionfactorAP1[66].
RAGEplaysacrucialroleinactivationofepithelialsodiumchannel(ENaC)throughPKC-gp91phoxsignaling,whichregulateslungfluidbalance[96].
Phosphoinositide3kinases/Akt(PI3K/Akt)Phosphoinositide3kinases(PI3K)areaclassofenzymeswhichphosphorylatethethirdhydroxylgroupintheinositolringofphosphotidylinositols(PtdIns)presentontheinnerleafletofplasmamembrane.
ThereareeightdistinctPI3Kcatalyticsubunitsthatarecapableofphosphorylatinginositollipids.
PhosphorylationofPtdInsbyvariousPI3KsleadstoformationofPhosphatidylinositol3phosphate(PI(3)P),Phosphatidylinositol(3,4)-bisphosphate(PI(3,4)P2),andPhosphatidyl-inositol(3,4,5)-trisphosphate(PI(3,4,5)P3),whichareimportantsignalingmolecules.
Thesephosphoinositidesparticipateinsignaltransductionpathwaysbybindingtolipidbindingdomainslikethepleckstrindomainofwww.
proteomics-journal.
comPage15ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Akt/PKBandphosphoinositidedependentkinase-1(PDK1),zincfingerFYVEdomainandPXdomainsofproteins[97].
ActivationofPI3Kislinkedtoanincrediblydiversesetofkeycellularfunctions,includingcellgrowth,proliferation,motility,differentiation,survivalandintracellulartrafficking.
CellularPI3Kactivitiesarebalancedbyphosphoinositide3-phosphataseactivity,foundinthetumorsuppressorproteinPTENandinmembersofthemyotubularinfamily.
Alteredactivationofthesekinasesisassociatedwithvarioushumandiseasesincludingcancer.
PI3KsareactivateddownstreamtoGproteincoupledreceptors(GPCRs),receptortyrosinekinase(RTK),aswellasRAGE[98].
AGE/RAGEaxishasbeenreportedtotransactivateInsulinlikegrowthfactor1receptor(IGF-1R)causingactivationofPI3K/Aktin3T3adipocytesleadingtotheirdifferentiation.
UponbindingofAGEstoRAGE,itwasobservedthatRAGEcouplestoNADPHoxidasetostimulateSrckinase.
TheactivatedSrckinaseinturnphosphorylatesandactivatesIGF-1receptorandPI3K-Aktpathway.
ThisadipogenesiscausedduetoAGE/RAGEandactivationofPI3K/Aktcanleadtoobesity,orpredisposeapersontodiabetes,orcausediabeticcomplications[99].
Furthermore,activationofPI3KduetoAGE/RAGEhasbeenimplicatedindiabeticnephropathy,whichisassociatedwithdepositionofextracellularmatrixproteins(e.
g.
collagen)intheglomerularbasementmembrane.
AGE/RAGEinteractioncausedproductionofROSandactivationofSrckinasecausingPI3Kphosphorylation.
InterestinglyPI3KcauseddownstreamactivationofPKBandki-Ras,whichrecruitsERKarmoftheMAPK[100].
Diabeticnephropathyischaracterizedbyproteinuriawhichreflectsmalfunctioningofglomerularvisceralepithelialcells,namelypodocytes.
Clinicalandexperimentalevidenceshowsthatdiabeticnephropathyischaracterizedbypodocytehypertrophycausinganincreaseinslitlengthandwidth,andalsoreductioninthewww.
proteomics-journal.
comPage16ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
numberofpodocytesperglomerulus[101,102].
AGE/RAGEsignalinginpodocyteshasbeenshowntocausepodocyteapoptosis.
Inculturedmurinepodocytes,AGE/RAGEaxiscausesdephosphorylationofAktensuingintranscriptionalactivationofFOXO4andtherebyapoptosis[36].
TherenninangiotensinsystemresponsibleformaintainingbloodvolumeandpressureisalsoaffectedbyAGE/RAGEsignalingtocausepodocyteapoptosis.
ThiseffectwasdependentonPI3K/Aktmediatedphosphorylation[103].
Indiabeticuremia,neutrophilsexhibitedanincreasedadhesiontoAGE-collagenthroughinteractionwithRAGEbyactivationofPI3K/Aktpathway[104].
ThePI3K/Akt/mTORpathwayisanegativeregulatorofautophagy.
AGEsinduceproliferationofrataorticvascularsmoothmusclecells(VSMCs)byinducingautophagyviainactivationofPI3K/Aktpathwaytherebyacceleratingatherosclerosisindiabeticconditions[105].
Inhumanumbilicalvascularendothelialcells(HUVECs),AGEscauseROSmediatedactivationofPI3K/Akt/IкB/NF-кBandJNK/AP1leadingtoincreasedexpressionofE-selectin,whichisassociatedwithprogressionofatherosclerosis[78].
HyperlipidemiaisshowntoincreasetheexpressionofHMGB1inserum,heartandlungtissuesofatheroscleroticmice,whichstimulatesRAGEsignalingthroughactivationofAktandproductionofproinflammatorycytokines[106].
Lectinlikeoxidizedlowdensitylipoproteinreceptor1(LOX1)isascavengerreceptorofoxidizedlowdensitylipoproteins(oxLDL),knowntobeelevatedinatheroscleroticplaquesandimplicatedintheinitiationandprogressionofatherosclerosis.
AGE/RAGEinteractionisproposedtocausetheupregulationofLOX1inaorticendothelialcellsbydownstreamactivationofPI3KwhichphosphorylatesmTORC2andAkt,indirectlyplayingaroleinatherosclerosis[107].
Indiabeticcardiovascularwww.
proteomics-journal.
comPage17ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
disorders,AGE/RAGEinteractioninducestheexpressionofheparinaseandPI3K/Aktphosphorylationleadingtomigrationofmacrophages[108].
Indorsalrootgangliasensoryneurons,S100B/RAGEinteractionactivatesPI3K/AktandERKpathways,whichfurthercauseslipidperoxidationandcaspase-3activationresultinginneuronaldamage[109].
S100B/RAGEinteractionincreasestheexpressionandreleaseofchemokinesinmicrogliaresultinginactivationofSrc/Ras/PI3K/RhoA/ROCKpathwayalongwithp38,ERK,JNK-AP1andNF-кB[82].
Incarcinogenesis,hypoxiainducedexpressionofRAGEandP2X7RcausedincreasedphosphorylationofAktandERK,andNF-кBnucleartranslocation.
Further,NF-кBincreasedtheexpressionofMMP2and9contributingtocellularinvasivenessuponHMGB1treatment[110].
HMGB1/RAGEinteractionleadstogrowth,migrationandinvasionofcolorectalcarcinomacellsbythephosphorylationofERK1/2,Rac1,AktandproductionofMMP9.
WhereasAGEsledtotheproductionofiNOSandNF-кBp65toagreaterextentthanHMGB1highlightingthedifferentialeffectsofboththeligandsandsuggestingamorecloseassociationofHMGB1withcancer[111].
BindingofHMGB1toRAGEandTLR4inlungcancercellsisshowntocausetheactivationofPI3K/AktandERK1/2leadingtoincreasedproliferation.
Thiseffectwasobservedduetodecreasedexpressionofpro-apoptoticBaxproteinandincreasedlevelsofanti-apoptoticBcl-2protein[112].
TreatmentofCML-BSAtoHEK293causedphosphorylationofthep66adaptorproteintocausedownstreamphosphorylationofAkt.
ActivatedAktcausesthephosphorylationofforkheadtranscriptionfactor3referredtoasFKHRL1leadingtoitsinactivationandpreventingitsnucleartranslocationtherebysuppressingtheexpressionofmanganesesuperoxidedismutase(MnSOD).
DownregulationofMnSODfacilitatesROSproduction.
TheseeffectsweresuppressedinAGEReceptor-1(AGER1)overexpressingwww.
proteomics-journal.
comPage18ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
HEK293cellssuggestingtheprotectiveeffectofAGER-1againstoxidativestresscausedbyAGE/RAGEinteraction[113].
S100A8/A9causesgrowthofnormalaswellastransformedhumanepidermalkeratinocytesbyinteractionwithRAGE/DNAX-activatingprotein(DAP10)heterodimersanddownstreamactivationofAkt.
Thisinteractioninducedapoptosisbyactivationofcaspase8innormalhumanepidermalkeratinocytes,whereassustainedgrowthwasobservedinthetransformedcelllinesHaCaTandA431duetoenhancedactivationofAkt,suggestingvariableeffectsofAktdownstreamtoRAGE/DAP10[114].
JanusActivatedkinase(JAK)JAKisafamilyoftyrosinekinasesthatactivatesignaltransducersandactivatorsoftranscription(STAT)inresponsetocytokinesandgrowthfactorsaswellasbyRAGEactivation.
JAKphosphorylatesSTATresultinginitsdimerizationandnucleartranslocation.
STATregulatestheexpressionofgenesinvolvedinantiviraldefence,embryogenesis,cellgrowth,cellsurvival,andcellmotility[115-120].
ConstitutiveactivationofmembersofSTATfamilyisassociatedwithseveraltypesofcancers.
TyrosinephosphorylatedSTAT1,3,and5arepersistentlyfoundinmostofthemalignancies.
AGE/RAGEaxisactivatesJAKinmyeloidleukemiawhereitinducesSTAT3phosphorylation[121-124].
RAGEpromotespancreaticcancerbyregulatingconjugationofATG12-ATG5,autophagyrelatedproteins;andBECN1-PIK3C3(Beclin1-Phosphatidylinositol3-kinasecatalyticsubunittype3)complexformation,whichresultsinsecretionofIL6,andmitochondrialSTAT3activation[125,126].
AGE/RAGEinteractionincreasestyrosinephosphorylationofHsp70,cyclinE,andcyclinD1inJAKdependentmanner.
ItalsoincreasesHsp70proteininteractionwithSTAT1,STAT3,STAT5b,cyclinD1,andcyclinE.
ThesefindingsindicatethatAGEsinducemitogenesisbyactivatingtheJAK2,STAT1,STAT3,andSTAT5-www.
proteomics-journal.
comPage19ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
cyclinD1pathwayinNRK-49Fcells[127].
Whileinhepatocytes,RAGEshowsanti-tumoureffectsbydecreasingp65levels,NF-кBactivation,cyclinD1productionandIL6mediatedSTAT3phosphorylation,therebydownregulatingPim1andsuppressingthehyperplasticresponse.
BlockadeofRAGErescuesliverremnantsfromadaptiveproliferationtriggeredbyMyd88signaling[128].
Indiabeticnephropathy,JAK2-STAT1/STAT3pathwayisinvolvedinAGE-inducedtype-1collagenproductioninkidneyfibroblastcells,whichisconsideredasamajorcauseofrenalfibrosis[129].
JAK/STATsignalingisalsoinvolvedinthedevelopmentofosteoarthritis.
IL-7activatesJAK/STATpathwayinchondrocytesleadingtoincreasedsecretionofS100A4whichinteractswithRAGEinautocrinemannerandstimulatesMMP13production[130].
AGEinducesactivationofMMP13,disintegrinandmetalloproteinasewiththrombospondinmotifs(ADAMTS)inaJAK/STAT3dependentmanner,whichisinvolvedincollagenIIandproteoglycanproductionandcontributestoosteoarthritis[131].
Duringembryogenesis,RAGEisinvolvedinneuronaldifferentiation.
LigandssuchasHumanglycatedalbumin,S100BandHMGB1arefoundtoincreasetheneuritegrowth.
TheseligandsactivateJAK/STAT,ERKandNF-кBwhichplayanimportantroleinaxonaloutgrowth[132].
GlycogenSynthaseKinase-3beta(GSK3β)GSK3βisaprolinedirectedserine/threoninekinase.
Itwasinitiallyidentifiedasakinasethatphosphorylatesglycogensynthase,akeyenzymeinglycogensynthesis[133,134].
Apartfromitsinvolvementinglycogensynthesis,itplaysanimportantroleinseveralbiologicalprocessessuchasdifferentiation,neurogenesis,cellfatedetermination,proliferation,andsurvival[135].
GSK3βactivityisdeterminedbyitsphosphorylationstatus;phosphorylationofser9decreasesitsactivity,whereasdephosphorylationactivatesit.
Itsactivityisalsowww.
proteomics-journal.
comPage20ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
enhancedbyphosphorylationoftyrosine216[136-138].
GSK3βphosphorylationisregulatedbyseveralkinasesincludingERKs,MAPKAPK-1,Akt,fewisoformsofPKC,proteinkinaseAandp70S6kinase.
Itisassociatedwithnumerousdisordersincludingdiabetes,cardiovasculardisorders,Parkinson's,Alzheimer'sdisease,amyotrophiclateralsclerosis,multiplesclerosisandcancer[139-141].
AGE/RAGEdependentGSK3βactivationinducesTauhyperphosphorylationinSK-N-SHcells,primaryhippocampalcellsandratbrain.
Inneuronalcells,AGE/RAGEinteractionleadstoactivationofGSK3β,ERK,p38MAPK,whichareassociatedwithmemorydeterioration,declineofsynapticproteins,andimpairmentoflong-termpotentiation.
BlockadeofRAGEattenuatedalltheAGEinducedimpairments,suggestingavitalroleofAGE/RAGEmediatedGSK3βstimulationinprogressionofAlzheimer'sdisease[142,143].
MethylglyoxalinducesGSK3βandp38MAPKactivationwhichleadstotauhyperphosphorylationinNeuro-2acells[144,145].
ThisisperhapsduetoincreasedRAGEsignalingasaresultofAGEformationpromotedbymethylglyoxal.
Apartfromtaupathology,GSK3βisalsoinvolvedinAβsynthesisandapoptoticneuronaldeath,whicharethemajorhallmarksofAD[146].
AGE/RAGEinteractionleadstoGSK3βactivationduringhypoxia/reoxygenationofcardiomyocyte[70].
AGEaccumulationandRAGEexpressionissignificantlyincreasedindiabeticratswithcardiomyopathyalongwithmarkedincreaseinGSK3βactivitywhichisreducedaftercurcumintreatmentbyAktphosphorylation[147].
However,therearenostudiesshowingRAGEdependentGSK3βactivationwithrespecttootherRAGEligandsincludingAβ,HMGB1andS100B,whicharefoundtobesignificantlyincreasedinAlzheimer'sdisease,cardiovasculardisordersandcancer[148-151].
www.
proteomics-journal.
comPage21ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
AnalysisofglobalphosphoproteomeinRAGEsignalingRAGEcanpotentiallyinteractwithmultipleligandsandactivateseveraldifferentsignaltransductionpathwaysinvolvingvariousproteinkinasesasdescribedabove.
Thesekinasesphosphorylatedownstreamsignalingmoleculesandcausespecificcellularresponse.
Askinaseshavemultiplesubstrates,thephosphorylationofasubstratedependsonitsexpression,localization,andcelltype.
OnesuchexampleisGSK3β,whichphosphorylatesglycogensynthaseinhepatocytesandregulatesglycogenmetabolism.
Whereas,inhumanembryonickidneycells(HEK293T),itphosphorylatesbetacateninrenderingittoproteasomaldegradation[152],andinneuronalcells,GSK3βactsdownstreamtoRAGEandhyperphosphorylatestaucausingneuronaldamage.
ThisobservationimpelledustoanalysephosphorylationsubstratesofkinasesinvolvedinRAGEsignaling.
Here,wehavefocusedonfewmajorkinases,namelyPKC,ERK,PI3K/Akt,p38,JAK,GSK3βandJNK,allofwhichareknowntoactdownstreamofRAGE.
AcomprehensivelistofproteinsubstratesforthesekinaseswasminedfromPhosPhoSitePlusdatabase[153].
Adetailedworkflowofdataminingandanalysisofphosphosubstratesisdepictedintheflowchart(Fig3).
Altogether,1045differentsubstrateswerefoundtobeassociatedwiththese7kinases(SupplementaryFig1).
Amongstthese7kinases,PKCandERKappeartophosphorylatemaximumnumberofsubstrates,andsignificantnumbersofthesephosphosubstratesareuniquetothesetwokinases(SupplementaryTable1and2),manyarealsosharedwithotherkinases.
Oneinterestingexampleisp21Cip1(p21Waf1),aproteininvolvedincellcycleregulation,wasfoundtobephosphorylatedbysixdifferentkinasesinthisanalysis.
Interestingly,itwasalsoshowntobephosphorylatedinresponsetoRAGEmediatedsignaling.
Overexpressionofwww.
proteomics-journal.
comPage22ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
RAGEcausedsignificantincreaseinp21Waf1induction,whichregulatescellproliferationinL6myoblasts[40].
Further,wehaveanalyzedtheassociationofthesephosphosubstrateswithdiseasessuchascancer,diabetes,cardiovascularandneurodegenerativediseases(Fig4).
Outofthe1045phosphosubstrates,thereare772,65,27and10phosphosubstratesfoundtobeassociatedwithcancer,neurodegenerativediseases,diabetesandcardiovasculardisordersrespectively(Fig3,SupplementaryTable3).
AninterestingobservationisthatthenumberofphosphosubstratesassociatedwithcancerisquitelargeandmanyofthesecanbepotentiallyphophorylatedbyrespectivekinasesinresponsetoRAGEactivation.
Insupportofthisnotion,involvementofRAGEincancerisbecomingincreasinglyevidentfromseveralstudies[44-53].
ERK2,akeyplayerinMAPKsignalingistheonlyphosphosubstratethatisassociatedwithallfourdiseases.
Inaddition,wehaveidentified11phosphosubstratesassociatedwiththreeofthediseases,whichincludeMARCKS,MAP4,Tau-8,Akt1,GSK3β,eNOS,ERK1,p70S6K,JNK1,AMPKA1andCREB.
ExceptMARCKSandMAP4,allotherproteinsareshowntobeinvolvedinRAGEsignaling,whichsupportsthisanalysis.
Tauiso8andMAP4arecytoskeletalproteinswhichplayanimportantroleinmicrotubuleassembly.
FunctionallossofTauiso8isassociatedwithseveralneurodegenerativedisordersincludingfrontotemporaldementia,Alzheimer'sdisease,pick'sdiseaseetc[154].
Endothelialnitricoxidesynthase(eNOS)producesnitricoxideinbloodvesselsandactsasaregulatorofenergymetabolisminthebody.
Itisassociatedwithseveraldisordersincludingdiabetes,hypertension,cancer,Alzheimer's,cardiovasculardisease,stroke,andatherosclerosis[155].
AGEshaveshowntoreduceeNOSphosphorylationanddown-regulationviaRAGEcausingendothelialdysfunction[156,157].
p70S6Kisaserinethreoninekinasethatactswww.
proteomics-journal.
comPage23ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
downstreamtoPI3K/Akt/mTORpathway.
Itphosphorylatesribosomalproteinandinducesproteinsynthesis.
ChronicintakeofAGEsmediatesp70S6KphosphorylationandRAGEup-regulationinrats.
Itisimplicatedinmultiplediseasesincludinginsulinresistance,skeletalmusclehypertrophyaswellasageing[158-160].
AMPKA1isacatalyticsubunitofAMPactivatedproteinkinaseknowntobeinvolvedinseveralbiologicalprocessessuchaslipidbiosynthesis,glucosehomeostasis,apoptosis,signaltransductionetc.
AβinducestheformationofautophagosomesinbrainsofAPP/PS1transgenicmiceaswellasinSH-SY5YcellsbyinteractionwithRAGEbyAMPKactivation[161,162].
CREBisatranscriptionfactorwhichbindswiththecAMPresponsiveelementandregulatestheexpressionofseveralgenes.
DeregulationofCREBfunctionisassociatedwithdisorderslikeAlzheimer'sdisease,huntington'sdiseaseandcancer.
AmphoterininducedCREBphosphorylationviaRAGEisinvolvedintheexpressionofchromograninsAandBinneuroblastomacellsleadingtotheirdifferentiation[163-165].
Myristoylatedalanine-richC-kinasesubstrate(MARCKS)isanacidicproteininvolvedintheregulationofcellproliferation,cellularsecretionandtransport,cellintegrityandmotility.
ThephosphorylationstatusofthisproteininRAGEsignalingneedstobeinvestigated[166,167].
ThisanalysisclearlysuggestedthatRAGEmediatedkinaseactivationcanpotentiallyresultinphosphorylationofseveraloftheirsubstrates.
Thepossibilityofaproteingettingphosphorylatedismorewhenitisasharedasasubstratebymultiplekinases.
Therefore,itisimportantnottounderminetheinvolvementofvariousknownsubstratesofthekinasesactivatedinRAGEsignaling.
Inconclusion,thisreviewemphasizesonthevitalroleofRAGEsignalinginvariouscellularprocessessuchasproliferation,apoptosis,migration,inflammation,immuneresponse,autophagyetc,andprovidesinsightintheplausibleroleofRAGEactivatedphosphoproteomeinseveraldiseases.
www.
proteomics-journal.
comPage24ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Acknowledgement:ThisworkissupportedbyCSIR-NetworkProjectBSC0111.
KBB,SBBandRSKacknowledgeCSIRandUGCforresearchfellowship.
GVPisgratefultoLadyTataMemorialTrustforaresearchfellowship.
Theauthorshavedeclarednoconflictofinterest.
References1.
Schmidt,A.
M.
,DuYan,S.
,Yan,S.
F.
,Stern,D.
M.
,ThemultiligandreceptorRAGEasaprogressionfactoramplifyingimmuneandinflammatoryresponses.
JClinInvest.
2001,108,949-955.
2.
Harashima,A.
,Yamamoto,Y.
,Cheng,C.
,Tsuneyama,K.
etal.
,Identificationofmouseorthologueofendogenoussecretoryreceptorforadvancedglycationend-products:structure,functionandexpression.
BiochemJ.
2006,396,109-115.
3.
Huttunen,H.
J.
,Fages,C.
,Rauvala,H.
,Receptorforadvancedglycationendproducts(RAGE)-mediatedneuriteoutgrowthandactivationofNF-кBrequirethecytoplasmicdomainofthereceptorbutdifferentdownstreamsignalingpathways.
JBiolChem.
1999,274,19919-19924.
4.
Kalea,A.
Z.
,Schmidt,A.
M.
,Hudson,B.
I.
,AlternativesplicingofRAGE:rolesinbiologyanddisease.
FrontBiosci.
2011,17,2756-2770.
5.
Kulkarni,M.
J.
,Korwar,A.
M.
,Mary,S.
,Bhonsle,H.
S.
,Giri,A.
P.
,Glycatedproteome:Fromreactiontointervention.
ProteomicsClinAppl.
2013,7,155-170.
6.
Kislinger,T.
,Fu,C.
,Huber,B.
,Qu,W.
etal.
,Nε-(carboxymethyl)lysineadductsofproteinsareligandsforreceptorforadvancedglycationendproductsthatactivatecellsignalingpathwaysandmodulategeneexpression.
JBiolChem.
1999,274,31740-31749.
www.
proteomics-journal.
comPage25ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
7.
Wang,Z.
,Jiang,Y.
,Liu,N.
,Ren,L.
etal.
,Advancedglycationend-productNε-carboxymethyl-Lysineacceleratesprogressionofatheroscleroticcalcificationindiabetes.
Atherosclerosis2012,221,387-396.
8.
Reynolds,T.
M.
,CHEMISTRYOFNONENZYMICBROWNING.
I.
THEREACTIONBETWEENALDOSESANDAMINES.
AdvFoodRes.
1963,12,1-52.
9.
Bunn,H.
F.
,Nonenzymaticglycosylationofprotein:relevancetodiabetes.
AmJMed.
1981,70,325-330.
10.
Ahmed,M.
U.
,Dunn,J.
A.
,Walla,M.
D.
,Thorpe,S.
R.
,Baynes,J.
W.
,Oxidativedegradationofglucoseadductstoprotein.
Formationof3-(Nepsilon-lysino)-lacticacidfrommodelcompoundsandglycatedproteins.
JBiolChem.
1988,263,8816-8821.
11.
Ahmed,M.
U.
,BrinkmannFrye,E.
,Degenhardt,T.
P.
,Thorpe,S.
R.
,Baynes,J.
W.
,N-epsilon-(carboxyethyl)lysine,aproductofthechemicalmodificationofproteinsbymethylglyoxal,increaseswithageinhumanlensproteins.
BiochemJ.
1997,324,565-570.
12.
Dyer,D.
G.
,Blackledge,J.
A.
,Thorpe,S.
R.
,Baynes,J.
W.
,Formationofpentosidineduringnonenzymaticbrowningofproteinsbyglucose.
Identificationofglucoseandothercarbohydratesaspossibleprecursorsofpentosidineinvivo.
JBiolChem.
1991,266,11654-11660.
13.
Beisswenger,P.
J.
,Szwergold,B.
S.
,Yeo,K.
T.
,Glycatedproteinsindiabetes.
ClinLabMed.
2001,21,53-78.
14.
Charonis,A.
S.
,Tsilibary,E.
C.
,StructuralandfunctionalchangesoflamininandtypeIVcollagenafternonenzymaticglycation.
Diabetes1992,41,49-51.
www.
proteomics-journal.
comPage26ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
15.
Chen,L.
,Wei,Y.
,Wang,X.
,He,R.
,Ribosylationrapidlyinducesalpha-synucleintoformhighlycytotoxicmoltenglobulesofadvancedglycationendproducts.
PLoSOne2010,5,e9052.
16.
Bansode,S.
B.
,Chougale,A.
D.
,Joshi,R.
S.
,Giri,A.
P.
etal.
,Proteomicanalysisofproteaseresistantproteinsinthediabeticratkidney.
MolCellProteomics2013,12,228-236.
17.
Neeper,M.
,Schmidt,A.
M.
,Brett,J.
,Yan,S.
D.
etal.
,Cloningandexpressionofacellsurfacereceptorforadvancedglycosylationendproductsofproteins.
JBiolChem.
1992,267,14998-15004.
18.
Alexiou,P.
,Chatzopoulou,M.
,Pegklidou,K.
,Demopoulos,V.
J.
,RAGE:amulti-ligandreceptorunveilingnovelinsightsinhealthanddisease.
CurrMedChem.
2010,17,2232-2252.
19.
DuYan,S.
,Chen,X.
,Fu,J.
,Chen,M.
etal.
,RAGEandamyloid-βpeptideneurotoxicityinAlzheimer'sdisease.
Nature1996,382,685-691.
20.
Hartmann,T.
,Bieger,S.
C.
,Brühl,B.
,Tienari,P.
J.
etal.
,DistinctsitesofintracellularproductionforAlzheimer'sdiseaseAβ40/42amyloidpeptides.
NatMed.
1997,3,1016-1020.
21.
Takuma,K.
,Fang,F.
,Zhang,W.
,Yan,S.
etal.
,RAGE-mediatedsignalingcontributestointraneuronaltransportofamyloid-βandneuronaldysfunction.
ProcNatlAcadSciUSA2009,106,20021-20026.
22.
Ostendorp,T.
,Leclerc,E.
,Galichet,A.
,Koch,M.
etal.
,StructuralandfunctionalinsightsintoRAGEactivationbymultimericS100B.
EMBOJ.
2007,26,3868-3878.
23.
Huttunen,H.
J.
,Kuja-Panula,J.
,Sorci,G.
,Agneletti,A.
L.
etal.
,CoregulationofneuriteoutgrowthandcellsurvivalbyamphoterinandS100proteinsthroughwww.
proteomics-journal.
comPage27ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
receptorforadvancedglycationendproducts(RAGE)activation.
JBiolChem.
2000,275,40096-40105.
24.
Sims,G.
P.
,Rowe,D.
C.
,Rietdijk,S.
T.
,Herbst,R.
,Coyle,A.
J.
,HMGB1andRAGEininflammationandcancer.
AnnuRevImmunol.
2009,28,367-388.
25.
Shaw,S.
S.
,Schmidt,A.
M.
,Banes,A.
K.
,Wang,X.
etal.
,S100B/RAGE-mediatedaugmentationofangiotensinII-inducedactivationofJAK2invascularsmoothmusclecellsisdependentonPLD2.
Diabetes2003,52,2381-2388.
26.
Klune,J.
R.
,Dhupar,R.
,Cardinal,J.
,Billiar,T.
R.
,Tsung,A.
,HMGB1:endogenousdangersignaling.
MolMed.
2008,14,476-484.
27.
Taguchi,A.
,Blood,D.
C.
,delToro,G.
,Canet,A.
etal.
,BlockadeofRAGE-amphoterinsignalingsuppressestumourgrowthandmetastases.
Nature2000,405,354-360.
28.
Mulder,D.
J.
,Water,T.
V.
D.
,Lutgers,H.
L.
,Graaff,R.
etal.
,Skinautofluorescence,anovelmarkerforglycemicandoxidativestress-derivedadvancedglycationendproducts:anoverviewofcurrentclinicalstudies,evidence,andlimitations.
DiabetesTechnolTher.
2006,8,523-535.
29.
Meerwaldt,R.
,Links,T.
,Graaff,R.
,Thorpe,S.
R.
etal.
,Simplenoninvasivemeasurementofskinautofluorescence.
AnnNYAcadSci.
2005,1043,290-298.
30.
Lutgers,H.
L.
,Graaff,R.
,Links,T.
P.
,Ubink-Veltmaat,L.
J.
etal.
,Skinautofluorescenceasanoninvasivemarkerofvasculardamageinpatientswithtype2diabetes.
DiabetesCare2006,29,2654-2659.
31.
Ramasamy,R.
,Yan,S.
F.
,Schmidt,A.
M.
,RAGE:therapeutictargetandbiomarkeroftheinflammatoryresponse-theevidencemounts.
JLeukocBiol.
2009,86,505-512.
www.
proteomics-journal.
comPage28ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
32.
Cargnello,M.
,Roux,P.
P.
,ActivationandfunctionoftheMAPKsandtheirsubstrates,theMAPK-activatedproteinkinases.
MicrobiolMolBiolRev.
2011,75,50-83.
33.
Yeh,C.
H.
,Sturgis,L.
,Haidacher,J.
,Zhang,X.
N.
etal.
,Requirementforp38andp44/p42mitogen-activatedproteinkinasesinRAGE-mediatednuclearfactor-kappaBtranscriptionalactivationandcytokinesecretion.
Diabetes2001,50,1495-1504.
34.
Tanikawa,T.
,Okada,Y.
,Tanikawa,R.
,Tanaka,Y.
,AdvancedglycationendproductsinducecalcificationofvascularsmoothmusclecellsthroughRAGE/p38MAPK.
JVascRes.
2009,46,572-580.
35.
Chang,P.
C.
,Chen,T.
H.
,Chang,C.
J.
,Hou,C.
C.
etal.
,Advancedglycosylationendproductsinduceinduciblenitricoxidesynthase(iNOS)expressionviaap38MAPK-dependentpathway.
KidneyInt.
2004,65,1664-1675.
36.
Chuang,P.
Y.
,Yu,Q.
,Fang,W.
,Uribarri,J.
,He,J.
C.
,AdvancedglycationendproductsinducepodocyteapoptosisbyactivationoftheFOXO4transcriptionfactor.
KidneyInt.
2007,72,965-976.
37.
Chung,A.
C.
,Zhang,H.
,Kong,Y.
Z.
,Tan,J.
J.
etal.
,Advancedglycationend-productsinducetubularCTGFviaTGFbeta-independentSmad3signaling.
JAmSocNephrol.
2010,21,249-260.
38.
Chen,S.
C.
,Guh,J.
Y.
,Hwang,C.
C.
,Chiou,S.
J.
etal.
,Advancedglycationend-productsactivateextracellularsignal-regulatedkinaseviatheoxidativestress-EGFreceptorpathwayinrenalfibroblasts.
JCellBiochem.
2010,109,38-48.
39.
Li,J.
H.
,Huang,X.
R.
,Zhu,H.
J.
,Oldfield,M.
etal.
,AdvancedglycationendproductsactivateSmadsignalingviaTGF-β-dependentand-independentwww.
proteomics-journal.
comPage29ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
mechanisms:implicationsfordiabeticrenalandvasculardisease.
FASEBJ.
2004,18,176-178.
40.
Li,Y.
,Liu,S.
,Zhang,Z.
,Xu,Q.
etal.
,RAGEmediatesaccelerateddiabeticveingraftatherosclerosisinducedbycombinedmechanicalstressandAGEsviasynergisticERKactivation.
PLoSOne2012,7,e35016.
41.
Xu,D.
,Young,J.
H.
,Krahn,J.
M.
,Song,D.
etal.
,StableRAGE–heparansulfatecomplexesareessentialforsignaltransduction.
ACSChemBiol.
2013,8,1611-1620.
42.
Spiekerkoetter,E.
,Guignabert,C.
,deJesusPerez,V.
,Alastalo,T.
P.
etal.
,S100A4andbonemorphogeneticprotein-2codependentlyinducevascularsmoothmusclecellmigrationviaphospho-extracellularsignal-regulatedkinaseandchlorideintracellularchannel4.
CircRes.
2009,105,639-647.
43.
Shi,L.
,Yu,X.
,Yang,H.
,Wu,X.
,AdvancedglycationendproductsinducehumancornealepithelialcellsapoptosisthroughgenerationofreactiveoxygenspeciesandactivationofJNKandp38MAPKpathways.
PLoSONE2013,8,ArticleIDe66781.
44.
Ishihara,K.
,Tsutsumi,K.
,Kawane,S.
,Nakajima,M.
,Kasaoka,T.
,Thereceptorforadvancedglycationend-products(RAGE)directlybindstoERKbyaD-domain-likedockingsite.
FEBSLett.
2003,550,107-113.
45.
Riuzzi,F.
,Sorci,G.
,Donato,R.
,TheAmphoterin(HMGB1)/receptorforadvancedglycationendproducts(RAGE)pairmodulatesmyoblastproliferation,apoptosis,adhesiveness,migration,andInvasiveness:FunctionalinactivationofRAGEinL6myoblastsresultsintumorformationinvivo.
JBiolChem.
2006,281,8242-8253.
www.
proteomics-journal.
comPage30ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
46.
Riuzzi,F.
,Sorci,G.
,Donato,R.
,RAGEexpressioninrhabdomyosarcomacellsresultsinmyogenicdifferentiationandreducedproliferation,migration,invasiveness,andtumorgrowth.
AmJPathol.
2007,171,947-961.
47.
Taguchi,A.
,Blood,D.
C.
,delToro,G.
,Canet,A.
etal.
,BlockadeofRAGE–amphoterinsignallingsuppressestumourgrowthandmetastases.
Nature2000,405,354-360.
48.
Ghavami,S.
,Rashedi,I.
,Dattilo,B.
M.
,Eshraghi,M.
etal.
,S100A8/A9atlowconcentrationpromotestumorcellgrowthviaRAGEligationandMAPkinase-dependentpathway.
JLeukocBiol.
2008,83,1484-1492.
49.
Lin,L.
,Zhong,K.
,Sun,Z.
,Wu,G.
,Ding,G.
,Receptorforadvancedglycationendproducts(RAGE)partiallymediatesHMGB1-ERKsactivationinclearcellrenalcellcarcinoma.
JCancerResClinOncol.
2012,13,11-22.
50.
Kao,Y.
H.
,Lin,Y.
C.
,Tsai,M.
S.
,Sun,C.
K.
etal.
,InvolvementofthenuclearhighmobilitygroupB1peptidesreleasedfrominjuredhepatocytesinmurinehepaticfibrogenesis.
BiochimBiophysActa2014,1842,1720-1732.
51.
Fuentes,M.
,Nigavekar,S.
,Arumugam,T.
,Logsdon,C.
etal.
,RAGEActivationbyS100PinColonCancerStimulatesGrowth,Migration,andCellSignalingPathways.
DisColonRectum2007,50,1230-1240.
52.
Tsoporis,J.
N.
,Izhar,S.
,Proteau,G.
,Slaughter,G.
,Parker,T.
G.
,S100B-RAGEdependentVEGFsecretionbycardiacmyocytesinducesmyofibroblastproliferation.
JMolCellCardiol.
2012,52,464–473.
53.
Tsoporis,J.
N.
,Izhar,S.
,Leong-Poi,H.
,Desjardins,J.
F.
etal.
,S100Binteractionwiththereceptorforadvancedglycationendproducts(RAGE):anovelreceptor-www.
proteomics-journal.
comPage31ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
mediatedmechanismformyocyteapoptosispostinfarction.
CircRes.
2010,106,93-101.
54.
Liu,R.
,Wu,C.
X.
,Zhou,D.
,Yang,F.
etal.
,Pinocembrinprotectsagainstβ-amyloid-inducedtoxicityinneuronsthroughinhibitingreceptorforadvancedglycationendproducts(RAGE)-independentsignalingpathwaysandregulatingmitochondrion-mediatedapoptosis.
BMCMed.
2012,10,105.
55.
Origlia,N.
,Righi,M.
,Capsoni,S.
,Cattaneo,A.
etal.
,ReceptorforAdvancedGlycationEndProduct-DependentActivationofp38Mitogen-ActivatedProteinKinaseContributestoAmyloid-β-MediatedCorticalSynapticDysfunction.
JNeurosci.
2008,28,3521-3530.
56.
Askarova,S.
,Yang,X.
,Sheng,W.
,Sun,G.
Y.
,Lee,J.
C.
,RoleofAbeta-receptorforadvancedglycationendproductsinteractioninoxidativestressandcytosolicphospholipaseA(2)activationinastrocytesandcerebralendothelialcells.
Neuroscience2011,199,375-385.
57.
Slowik,A.
,Merres,J.
,Elfgen,A.
,Jansen,S.
etal.
,Involvementofformylpeptidereceptorsinreceptorforadvancedglycationendproducts(RAGE)andamyloidbeta1-42-inducedsignaltransductioninglialcells.
Mol.
Neurodeg.
2012,7,55.
58.
McDonald,D.
,Bamberger,M.
,Combs,C.
,Landreth,G.
,b-amyloidfibrilactivateparallelmitogen-activatedproteinkinasepathwaysinmicrogliaandTHP-1monocytes.
JNeurosci.
1998,18,4451-4460.
59.
Kang,J.
H.
,Hwang,S.
M.
,Chung,I.
Y.
,S100A8,S100A9,andS100A12activateairwayepithelialcellstoproduceMUC5ACviaERKandNF-κBpathways.
2014www.
proteomics-journal.
comPage32ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
60.
Wolfson,R.
K.
,Chiang,E.
T.
,Garcia,J.
G.
,HMGB1induceshumanlungendothelialcellcytoskeletalrearrangementandbarrierdisruption.
Microvasc.
Res.
2011,81,189-197.
61.
He,Z.
W.
,Qin,Y.
H.
,Wang,Z.
W.
,Chen,Y.
etal.
,HMGB1actsinsynergywithlipopolysaccharideinactivatingrheumatoidsynovialfibroblastsviap38MAPKandNF-kappaBsignalingpathways.
MediatorsInflamm.
2013,596716.
62.
Qin,Y.
H.
,Dai,S.
M.
,Tang,G.
S.
,Zhang,J.
etal.
,HMGB1enhancestheproinflammatoryactivityoflipopolysaccharidebypromotingthephosphorylationofMAPKp38throughreceptorforadvancedglycationendproducts.
JImmunol.
2009,183,6244-6250.
63.
Weston,C.
R.
,Davis,R.
J.
,TheJNKsignaltransductionpathway.
CurrOpinGenetDev.
2002,12,14-21.
64.
Bogoyevitch,M.
A.
,Kobe,B.
,UsesforJNK:themanyandvariedsubstratesofthec-JunN-terminalkinases.
MicrobiolMolBiolRev.
2006,70,1061-1095.
65.
Nah,S.
S.
,Choi,I.
Y.
,Lee,C.
K.
,Oh,J.
S.
etal.
,EffectsofadvancedglycationendproductsontheexpressionofCOX-2,PGE2andNOinhumanosteoarthriticchondrocytes.
Rheumatology2008,47,425-431.
66.
Nah,S.
S.
,Choi,I.
Y.
,Yoo,B.
,Kim,Y.
G.
etal.
,Advancedglycationendproductsincreasesmatrixmetalloproteinase-1,-3,and-13,andTNF-αinhumanosteoarthriticchondrocytes.
FEBSlett.
2007,581,1928-1932.
67.
Zhang,F.
,Banker,G.
,Liu,X.
,Suwanabol,P.
A.
etal.
,ThenovelfunctionofadvancedglycationendproductsinregulationofMMP9production.
JSurgRes.
2011,171,871-876.
www.
proteomics-journal.
comPage33ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
68.
Alikhani,M.
,MacLellan,C.
M.
,Raptis,M.
,Vora,S.
etal.
,AdvancedglycationendproductsinduceapoptosisinfibroblaststhroughactivationofROS,MAPkinases,andtheFOXO1transcriptionfactor.
AmJPhysiolCellPhysiol.
2007,292,C850-C856.
69.
Chang,J.
S.
,Wendt,T.
,Qu,W.
,Kong,L.
etal.
,Oxygendeprivationtriggersupregulationofearlygrowthresponse-1bythereceptorforadvancedglycationendproducts.
CircRes.
2008,102,905-913.
70.
Shang,L.
,Ananthakrishnan,R.
,Li,Q.
,Quadri,N.
etal.
,RAGEmodulateshypoxia/reoxygenationinjuryinadultmurinecardiomyocytesviaJNKandGSK-3βsignalingpathways.
PLoSOne2010,5,e10092.
71.
Aleshin,A.
,Ananthakrishnan,R.
,Li,Q.
,Rosario,R.
etal.
,RAGEmodulatesmyocardialinjuryconsequenttoLADinfarctionviaimpactonJNKandSTATsignalinginamurinemodel.
AmJPhysiolHeartCircPhysiol.
2008,294,H1823-H1832.
72.
Volz,H.
C.
,Laohachewin,D.
,Seidel,C.
,Lasitschka,F.
etal.
,S100A8/A9aggravatespost-ischemicheartfailurethroughactivationofRAGE-dependentNF-κBsignaling.
BasicResCardiol.
2012,107,1-16.
73.
Andrassy,M.
,Volz,H.
C.
,Igwe,J.
C.
,Funke,B.
etal.
,High-mobilitygroupbox-1inischemia-reperfusioninjuryoftheheart.
Circulation2008,117,3216-3226.
74.
Volz,H.
C.
,Seidel,C.
,Laohachewin,D.
,Kaya,Z.
etal.
,HMGB1:themissinglinkbetweendiabetesmellitusandheartfailure.
BasicResCardiol.
2010,105,805-820.
75.
Koh,E.
J.
,Yoon,S.
J.
,Lee,S.
M.
,Losartanprotectsliveragainstischaemia/reperfusioninjurythroughPPAR‐γactivationandreceptorforwww.
proteomics-journal.
comPage34ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
advancedglycationend‐productsdown‐regulation.
BrJPharmacol.
2013,169,1404-1416.
76.
Kokkola,R.
,Andersson,A.
,Mullins,G.
,Ostberg,T.
etal.
,RAGEisthemajorreceptorfortheproinflammatoryactivityofHMGB1inrodentmacrophages.
ScandJImmunol.
2005,61,1-9.
77.
Harja,E.
,Bu,D.
X.
,Hudson,B.
I.
,Chang,J.
S.
etal.
,VascularandinflammatorystressesmediateatherosclerosisviaRAGEanditsligandsinapoE–/–mice.
JClinInvest.
2008,118,183-194.
78.
Higai,K.
,Shimamura,A.
,Matsumoto,K.
,Amadori-modifiedglycatedalbuminpredominantlyinducesE-selectinexpressiononhumanumbilicalveinendothelialcellsthroughNADPHoxidaseactivation.
Clinchimacta.
2006,367,137-143.
79.
Fiuza,C.
,Bustin,M.
,Talwar,S.
,Tropea,M.
etal.
,Inflammation-promotingactivityofHMGB1onhumanmicrovascularendothelialcells.
Blood.
2003,101,2652-2660.
80.
Bianchi,R.
,Adami,C.
,Giambanco,I.
,Donato,R.
,S100BbindingtoRAGEinmicrogliastimulatesCOX-2expression.
JLeukocbiol.
2007,81,108-118.
81.
Bianchi,R.
,Giambanco,I.
,Donato,R.
,S100B/RAGE-dependentactivationofmicrogliaviaNF-κBandAP1:Co-regulationofCOX-2expressionbyS100B,IL-1βandTNF-α.
NeurobiolAging2010,31,665-677.
82.
Bianchi,R.
,Kastrisianaki,E.
,Giambanco,I.
,Donato,R.
,S100BproteinstimulatesmicrogliamigrationviaRAGE-dependentup-regulationofchemokineexpressionandrelease.
JBiolChem.
2011,286,7214-7226.
www.
proteomics-journal.
comPage35ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
83.
Esposito,G.
,Scuderi,C.
,Lu,J.
,Savani,C.
etal.
,S100BinducestauproteinhyperphosphorylationviaDickopff‐1up‐regulationanddisruptstheWntpathwayinhumanneuralstemcells.
JCellMolMed.
2008,12,914-927.
84.
Onyango,I.
G.
,Tuttle,J.
B.
,BennettJr,J.
P.
,AlteredintracellularsignalingandreducedviabilityofAlzheimer'sdiseaseneuronalcybridsisreproducedbyβ-amyloidpeptideactingthroughreceptorforadvancedglycationendproducts(RAGE).
MolCellNeurosci.
2005,29,333-343.
85.
Li,M.
,Sun,L.
,Luo,Y.
,Xie,C.
etal.
,High-mobilitygroupbox1releasedfromastrocytespromotestheproliferationofculturedneuralstem/progenitorcells.
IntJMolMed.
2014,34,705-714.
86.
Ohshiro,Y.
,Lee,Y.
,King,G.
L.
,Mechanismofdiabeticnephropathy:roleofproteinkinaseCactivation.
AdvStudMed.
2005,5,s11-s19.
87.
Geraldes,P.
,King,G.
L.
,ActivationofproteinkinaseCisoformsanditsimpactondiabeticcomplications.
CircRes.
2010,106,1319-1331.
88.
Sakaguchi,M.
,Murata,H.
,Yamamoto,K.
,Ono,T.
etal.
,TIRAP,anadaptorproteinforTLR2/4,transducesasignalfromRAGEphosphorylateduponligandbinding.
PLoSOne2011,6,e23132.
89.
Cassese,A.
,Esposito,I.
,Fiory,F.
,Barbagallo,A.
P.
etal.
,Inskeletalmuscleadvancedglycationendproducts(AGEs)inhibitinsulinactionandinducetheformationofmultimolecularcomplexesincludingthereceptorforAGEs.
JBiolChem.
2008,283,36088-36099.
90.
Warboys,C.
M.
,Toh,H.
B.
,Fraser,P.
A.
,RoleofNADPHoxidaseinretinalmicrovascularpermeabilityincreasebyRAGEactivation.
InvestOphthalmol.
Vis.
Sci.
2009,50,1319-1328.
www.
proteomics-journal.
comPage36ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
91.
Thallas-Bonke,V.
,Thorpe,S.
R.
,Coughlan,M.
T.
,Fukami,K.
etal.
,InhibitionofNADPHoxidasepreventsadvancedglycationendproduct-mediateddamageindiabeticnephropathythroughaproteinkinaseC-alphadependentpathway.
Diabetes2008,57,460-469.
92.
Yamagishi,S.
I.
,Commenton:Thallas-Bonkeetal.
(2008)InhibitionofNADPHOxidasePreventsAdvancedGlycationEndProduct–MediatedDamageinDiabeticNephropathyThroughaProteinKinaseC-α–DependentPathway:Diabetes57:460–469,2008.
Diabetes2008,57,e13-e13.
93.
Yu,L.
,Zhao,Y.
,Xu,S.
,Ding,F.
etal.
,AdvancedGlycationEndProduct(AGE)-AGEReceptor(RAGE)SystemUpregulatedConnexin43ExpressioninRatCardiomyocytesviaPKCandErkMAPKPathways.
IntJMolSci.
2013,14,2242-2257.
94.
Ding,Y.
,Kantarci,A.
,Badwey,J.
A.
,Hasturk,H.
,Malabanan,A.
,VanDyke,T.
E.
,Phosphorylationofpleckstrinincreasesproinflammatorycytokinesecretionbymononuclearphagocytesindiabetesmellitus.
JImmunol.
2007,179,647-654.
95.
Shanmugam,N.
,Kim,Y.
S.
,Lanting,L.
,Natarajan,R.
,Regulationofcyclooxygenase-2expressionbyligationofthereceptorofadvancedglycationendproducts.
J.
Biol.
Chem.
2003,278,34834-3484.
96.
Downs,C.
A.
,Kreiner,L.
H.
,Johnson,N.
M.
,Brown,L.
A.
,Helms,M.
N.
,RAGERegulatesLungFluidBalanceviaPKC-gp91phoxSignalingtoEpithelialSodiumChannels(ENaC).
AmJRespirCellMolBiol.
2014.
97.
Foster,F.
M.
,Traer,C.
J.
,Abraham,S.
M.
,Fry,M.
J.
,Thephosphoinositide(PI)3-kinasefamily.
JCellSci.
2003,116,3037-3040.
www.
proteomics-journal.
comPage37ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
98.
Cantley,L.
C.
,Thephosphoinositide3-kinasepathway.
Science2002,296,1655-1657.
99.
Yang,S.
J.
,Chen,C.
Y.
,Chang,G.
D.
,Wen,H.
C.
etal.
,ActivationofAktbyadvancedglycationendproducts(AGEs):involvementofIGF-1receptorandcaveolin-1.
PloSone2013,8,e58100.
100.
Xu,D.
,Kyriakis,J.
M.
,Phosphatidylinositol3′-kinase-dependentactivationofrenalmesangialcellKi-RasandERKbyadvancedglycationendproducts.
JBiolChem.
2003,278,39349-39355.
101.
Li,J.
J.
,Kwak,S.
J.
,Jung,D.
S.
,Kim,J.
J.
etal.
,Podocytebiologyindiabeticnephropathy.
KidneyIntSuppl.
2007,72,S36-S42.
102.
DallaVestra,M.
,Masiero,A.
,Roiter,A.
M.
,Saller,A.
etal.
,IspodocyteinjuryrelevantindiabeticnephropathyStudiesinpatientswithtype2diabetes.
Diabetes2003,52,1031-1035.
103.
Cheng,C.
L.
,Tang,Y.
,Zheng,Z.
,Liu,X.
etal.
,Advancedglycationend-productsactivatetherenin-angiotensinsystemthroughtheRAGE/PI3-Ksignalingpathwayinpodocytes.
ClinInvestMed.
2012,35,E282-E293.
104.
Touré,F.
,Zahm,J.
,Garnotel,R.
,Lambert,E.
etal.
,Receptorforadvancedglycationend-products(RAGE)modulatesneutrophiladhesionandmigrationonglycoxidatedextracellularmatrix.
Biochem.
J.
2008,416,255-261.
105.
Hu,P.
,Lai,D.
,Lu,P.
,Gao,J.
,He,H.
,ERKandAktsignalingpathwaysareinvolvedinadvancedglycationendproduct-inducedautophagyinratvascularsmoothmusclecells.
IntJMolMed.
2012,29,613-618.
www.
proteomics-journal.
comPage38ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
106.
Haraba,R.
,Suica,V.
I.
,Uyy,E.
,Ivan,L.
,Antohe,F.
,Hyperlipidemiastimulatestheextracellularreleaseofthenuclearhighmobilitygroupbox1protein.
CellTissueRes.
2011,346,361-368.
107.
Shiu,S.
W.
,Wong,Y.
,Tan,K.
C.
,Effectofadvancedglycationendproductsonlectin-likeoxidizedlowdensitylipoproteinreceptor-1expressioninendothelialcells.
JAtherosclerThromb.
2011,19,1083-1092.
108.
Qin,Q.
,Niu,J.
,Wang,Z.
,Xu,W.
etal.
,Heparanaseinducedbyadvancedglycationendproducts(AGEs)promotesmacrophagemigrationinvolvingRAGEandPI3K/AKTpathway.
CardiovascDiabetol.
2013,12,37.
109.
Vincent,A.
M.
,Perrone,L.
,Sullivan,K.
A.
,Backus,C.
etal.
,Receptorforadvancedglycationendproductsactivationinjuresprimarysensoryneuronsviaoxidativestress.
Endocrinology2007,148,548-558.
110.
Tafani,M.
,Schito,L.
,Pellegrini,L.
,Villanova,L.
etal.
,Hypoxia-increasedRAGEandP2X7RexpressionregulatestumorcellinvasionthroughphosphorylationofErk1/2andAktandnucleartranslocationofNF-κB.
Carcinogenesis2011,32,1167-1175.
111.
Kuniyasu,H.
,Chihara,Y.
,Kondo,H.
,Differentialeffectsbetweenamphoterinandadvancedglycationendproductsoncoloncancercells.
IntJCancer2003,104,722-727.
112.
Xu,X.
,Zhu,H.
,Wang,T.
,Sun,Y.
etal.
,ExogenousHigh‐MobilityGroupBox1InhibitsApoptosisandPromotestheProliferationofLewisCellsviaRAGE/TLR4‐DependentSignalPathways.
ScandJImmunol.
2014,79,386-394.
www.
proteomics-journal.
comPage39ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
113.
Cai,W.
,He,J.
C.
,Zhu,L.
,Chen,X.
etal.
,AGE-receptor-1counteractscellularoxidantstressinducedbyAGEsvianegativeregulationofp66shc-dependentFKHRL1phosphorylation.
AmJPhysiolCellPhysiol.
2008,294,C145-C152.
114.
Sakaguchi,M.
,Murata,H.
,Aoyama,Y.
,Hibino,T.
etal.
,DNAX-ActivatingProtein10(DAP10)MembraneAdaptorAssociateswithReceptorforAdvancedGlycationEndProducts(RAGE)andModulatestheRAGE-triggeredSignalingPathwayinHumanKeratinocytes.
JBiolChem.
2014,jbc-M114.
115.
Rawlings,J.
S.
,Rosler,K.
M.
,Harrison,D.
A.
,TheJAK/STATsignalingpathway.
JCellSci.
2004,117,1281-1283.
116.
Bierhaus,A.
,Humpert,P.
M.
,Morcos,M.
,Wendt,T.
etal.
,UnderstandingRAGE,thereceptorforadvancedglycationendproducts.
JMolMed.
2005,83,876-886.
117.
Rojas,A.
,Figueroa,H.
,Morales,E.
,Fuelinginflammationattumormicroenvironment:theroleofmultiligand/RAGEaxis.
Carcinogenesis2010,31,334-341.
118.
Schindler,C.
W.
,SeriesIntroduction.
JAK-STATsignalinginhumandisease.
JClinInvest.
2002,109,1133-1137.
119.
Constantinescu,S.
N.
,Girardot,M.
,Pecquet,C.
,MiningforJAK-STATmutationsincancer.
TrendsBiochemSci.
2008,33,122-131.
120.
Tang,J.
Z.
,Zuo,Z.
H.
,Kong,X.
J.
,Steiner,M.
etal.
,Signaltransducerandactivatoroftranscription(STAT)-5AandSTAT5Bdifferentiallyregulatehumanmammarycarcinomacellbehaviour.
Endocrinology2010,151,43-55.
121.
Kim,J.
Y.
,Park,H.
K.
,Yoon,J.
S.
,Kim,S.
J.
etal.
,Advancedglycationendproduct(AGE)-inducedproliferationofHELcellsviareceptorforAGE-relatedsignalpathways.
IntJOncol.
2008,33,493-501.
www.
proteomics-journal.
comPage40ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
122.
Sansone,P.
,Bromberg,J.
,Targetingtheinterleukin-6/Jak/statPathwayinhumanmalignancies.
JClinOncol.
2012,30,1005-1014.
123.
Bromberg,J.
,Statproteinsandoncogenesis.
JClinInvest.
2002,109,1139-1142.
124.
O'Shea,J.
J.
,Gadina,M.
,Schreiber,R.
D.
,CytokineSignalingin2002:NewSurprisesintheJak/StatPathway.
Cell2002,109,S121-S131.
125.
Kang,R.
,Tang,D.
,Lotze,M.
T.
,Zeh,H.
J.
,AGER/RAGE-mediatedautophagypromotespancreatictumorigenesisandbioenergeticsthroughtheIL6-pSTAT3pathway.
Autophagy2012,8,989-91.
126.
Kang,R.
,Loux,T.
,Tang,D.
,Schapiro,N.
E.
etal.
,Theexpressionofthereceptorforadvancedglycationendproducts(RAGE)ispermissiveforearlypancreaticneoplasia.
ProcNatlAcadSciUSA.
2012,109,7031-7036.
127.
Chen,S.
C.
,Guh,J.
Y.
,Chen,H.
C.
,Yang,Y.
L.
etal.
,Advancedglycationend-product-inducedmitogenesisisdependentonJanuskinase2-inducedheatshockprotein70innormalratkidneyinterstitialfibroblastcells.
TranslRes.
2007,149,274-281.
128.
Zeng,S.
,Zhang,Q.
Y.
,Huang,J.
,Vedantham,S.
etal.
,OpposingrolesofRAGEandMyd88signalinginextensiveliverresection.
FASEBJ.
2012,26,882-893.
129.
Huang,J.
S.
,Guh,J.
Y.
,Chen,H.
C.
,Hung,W.
C.
etal.
,Roleofreceptorforadvancedglycationend-product(RAGE)andtheJAK/STAT-signalingpathwayinAGE-inducedcollagenproductioninNRK-49Fcells.
JCellBiochem.
2001,81,102-113.
130.
Yammani,R.
R.
,Long,D.
,Loeser,R.
F.
,Interleukin-7stimulatessecretionofS100A4byactivatingtheJAK/STATsignalingpathwayinhumanarticularchondrocytes.
ArthritisRheum.
2009,60,792-800.
www.
proteomics-journal.
comPage41ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
131.
Huang,C.
Y.
,Lai,K.
Y.
,Hung,L.
F.
,Wu,W.
L.
etal.
,AdvancedglycationendproductscausecollagenIIreductionbyactivatingJanuskinase/signaltransducerandactivatoroftranscription3pathwayinporcinechondrocytes.
Rheumatology2011,50,1379-1389.
132.
Saleh,A.
,Smith,D.
R.
,Tessler,L.
,Mateo,A.
R.
etal.
,Receptorforadvancedglycationend-products(RAGE)activatesdivergentsignalingpathwaystoaugmentneuriteoutgrowthofadultsensoryneurons.
ExpNeurol.
2013,249,149-159.
133.
Fiol,C.
J.
,Wang,A.
,Roeske,R.
W.
,Roach,P.
J.
,Orderedmultisiteproteinphosphorylation.
Analysisofglycogensynthasekinase3actionusingmodelpeptidesubstrates.
JBiolChem.
1990,265,6061-6065.
134.
Embi,N.
,RYLATT,D.
B.
,COHEN,P.
,GlycogenSynthaseKinase‐3fromRabbitSkeletalMuscle.
EurJBiochem.
1980,107,519-527.
135.
Ali,A.
,Hoeflich,K.
P.
,Woodgett,J.
R.
,Glycogensynthasekinase-3:properties,functions,andregulation.
ChemRev.
2001,101,2527-2540.
136.
Yao,H.
B.
,Shaw,P.
C.
,Wong,C.
C.
,Wan,D.
C.
C.
,Expressionofglycogensynthasekinase-3isoformsinmousetissuesandtheirtranscriptioninthebrain.
JChemNeuroanat.
2002,23,291-297.
137.
Hughes,K.
,Nikolakaki,E.
,Plyte,S.
E.
,Totty,N.
F.
,Woodgett,J.
R.
,Modulationoftheglycogensynthasekinase-3familybytyrosinephosphorylation.
EMBOJ.
1993,12,803-808.
138.
Sutherland,C.
,Leighton,I.
A.
,Cohen,P.
,Inactivationofglycogensynthasekinase-3betabyphosphorylation:newkinaseconnectionsininsulinandgrowth-factorsignaling.
BiochemJ.
1993,296,15-19.
www.
proteomics-journal.
comPage42ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
139.
Tanji,C.
,Yamamoto,H.
,Yorioka,N.
,Kohno,N.
etal.
,A-kinaseanchoringproteinAKAP220bindstoglycogensynthasekinase-3β(GSK-3β)andmediatesproteinkinaseA-dependentinhibitionofGSK-3β.
JBiolChem.
2002,277,36955-36961.
140.
Grimes,C.
A.
,Jope,R.
S.
,Themultifacetedrolesofglycogensynthasekinase3βincellularsignaling.
ProgNeurobiol.
2001,65,391-426.
141.
Doble,B.
W.
,Woodgett,J.
R.
,GSK-3:tricksofthetradeforamulti-taskingkinase.
JCellSci.
2003,116,1175-1186.
142.
Li,X.
H.
,Lv,B.
L.
,Xie,J.
Z.
,Liu,J.
etal.
,AGEsinduceAlzheimer-liketaupathologyandmemorydeficitviaRAGE-mediatedGSK-3activation.
NeurobiolAging2012,33,1400-1410.
143.
Chen,S.
,An,F.
M.
,Yin,L.
,Liu,A.
R.
etal.
,Glucagon-likepeptide-1protectshippocampalneuronsagainstadvancedglycationendproduct-inducedtauhyperphosphorylation.
Neuroscience2014,256,137-146.
144.
Li,X.
H.
,Du,L.
L.
,Cheng,X.
S.
,Jiang,X.
etal.
,Glycationexacerbatestheneuronaltoxicityofβ-amyloid.
CellDeathDis.
2013,4,e673.
145.
Li,X.
H.
,Xie,J.
Z.
,Jiang,X.
,Lv,B.
L.
etal.
,MethylglyoxalinducestauhyperphosphorylationviapromotingAGEsformation.
NeuromolecularMed.
2012,14,338-348.
146.
Qu,Z.
S.
,Li,L.
,Sun,X.
J.
,Zhao,Y.
W.
etal.
,GlycogenSynthaseKinase-3RegulatesProductionofAmyloid-βPeptidesandTauPhosphorylationinDiabeticRatBrain.
ScientificWorldJournal2014.
147.
Yu,W.
,Wu,J.
,Cai,F.
,Xiang,J.
etal.
,Curcuminalleviatesdiabeticcardiomyopathyinexperimentaldiabeticrats.
PloSone2012,7,e52013.
www.
proteomics-journal.
comPage43ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
148.
Chaves,M.
L.
,Camozzato,A.
L.
,Ferreira,E.
D.
,Piazenski,I.
etal.
,SerumlevelsofS100BandNSEproteinsinAlzheimer'sdiseasepatients.
JNeuroinflammation2010,7.
149.
Li,J.
P.
,Lu,L.
,Wang,L.
J.
,Zhang,F.
R.
,Shen,W.
F.
,IncreasedserumlevelsofS100Barerelatedtotheseverityofcardiacdysfunction,renalinsufficiencyandmajorcardiaceventsinpatientswithchronicheartfailure.
ClinBiochem.
2011,44,984-988.
150.
Sun,K.
K.
,Ji,C.
,Li,X.
,Zhang,L.
etal.
,OverexpressionofhighmobilitygroupproteinB1correlateswiththeproliferationandmetastasisoflungadenocarcinomacells.
MolMedRep.
2013,7,1678-1682.
151.
Arancio,O.
,Zhang,H.
P.
,Chen,X.
,Lin,C.
etal.
,RAGEpotentiatesAβ‐inducedperturbationofneuronalfunctionintransgenicmice.
EMBOJ.
2004,23,4096-4105.
152.
Aberle,H.
,Bauer,A.
,Stappert,J.
,Kispert,A.
,Kemler,R.
β‐cateninisatargetfortheubiquitin–proteasomepathway.
EMBOJ.
1997,16,3797-3804.
153.
Hornbeck,P.
V.
,Kornhauser,J.
M.
,Tkachev,S.
,Zhang,B.
etal.
,PhosphoSitePlus:acomprehensiveresourceforinvestigatingthestructureandfunctionofexperimentallydeterminedpost-translationalmodificationsinmanandmouse.
NucleicAcidsRes.
2012,40,D261-D270.
154.
Ghidoni,R.
,Paterlini,A.
,Benussi,L.
,TranslationalproteomicsinAlzheimer'sdiseaseandrelateddisorders.
ClinBiochem.
2013,46,480-486.
155.
Atochin,D.
N.
,Wang,A.
,Liu,V.
W.
,Critchlow,J.
D.
etal.
,ThephosphorylationstateofeNOSmodulatesvascularreactivityandoutcomeofcerebralischemiainvivo.
JClinInvest.
2007,117,1961-1967.
www.
proteomics-journal.
comPage44ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
156.
Liang,C.
,Ren,Y.
,Tan,H.
,He,Z.
etal.
,RosiglitazoneviaupregulationofAkt/eNOSpathwaysattenuatesdysfunctionofendothelialprogenitorcells,inducedbyadvancedglycationendproducts.
BrJPharmacol.
2009,158,1865-1873.
157.
Chen,J.
,Jin,J.
,Song,M.
,Dong,H.
etal.
,C-reactiveproteindown-regulatesendothelialnitricoxidesynthaseexpressionandpromotesapoptosisinendothelialprogenitorcellsthroughreceptorforadvancedglycationend-products.
Gene2012,496,128-135.
158.
Chung,J.
,Grammar,T.
C.
,Lemon,K.
P.
,Kazlauskas,A.
,Blenis,J.
,PDGF-andinsulin-dependentpp70S6kactivationmediatedbyphosphatidylinositol-3-OHkinase.
Nature1994,370,71-75.
159.
Chung,J.
,Kuo,C.
J.
,Crabtree,G.
R.
,lenis,J.
,Rapamycin-FKBPspecificallyblocksgrowth-dependentactivationofandsignalingbythe70kdS6proteinkinases.
Cell1992,69,1227-1236.
160.
Murakami,H.
,Henry,G.
,Leonil,J.
,Galmier,M.
J.
etal.
,Effectofachronicintakeofhighdietaryglycatedproteinsonskeletalmuscleproteinmetabolismduringaginginnormalrats.
FASEBJ.
2011,25,233-2.
161.
Shaw,R.
J.
,LKB1andAMP‐activatedproteinkinasecontrolofmTORsignallingandgrowth.
ActaPhysiol.
2009,196,65-80.
162.
Son,S.
M.
,Jung,E.
S.
,Shin,H.
J.
,Byun,J.
,Mook-Jung,I.
,Aβ-inducedformationofautophagosomesismediatedbyRAGE-CaMKKβ-AMPKsignaling.
NeurobiolAging2012,33,1006-e11.
163.
Catalano,S.
,Giordano,C.
,Rizza,P.
,Gu,G.
,Barone,I.
,EvidencethatleptinthroughSTATandCREBsignalingenhancescyclinD1expressionandpromoteshumanendometrialcancerproliferation.
JCellPhysiol.
2009,218,490-500.
www.
proteomics-journal.
comPage45ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
164.
Sugars,K.
L.
,Rubinsztein,D.
C.
,TranscriptionalabnormalitiesinHuntingtondisease.
TrendsGenet.
200319,233-238.
165.
Huttunen,H.
J.
,Kuja-Panula,J.
,Rauvala,H.
,Receptorforadvancedglycationendproducts(RAGE)signalinginducesCREB-dependentchromograninexpressionduringneuronaldifferentiation.
JCellBiochem.
2002,277,38635-38646.
166.
Aderem,A.
,Signaltransductionandtheactincytoskeleton:therolesofMARCKSandprofilin.
TrendsBiochemSci.
1992,17,438-443.
167.
Green,T.
D.
,Crews,A.
L.
,Park,J.
,Fang,S.
,Adler,K.
B.
,Regulationofmucinsecretionandinflammationinasthma:aroleforMARCKSproteinBiochimBiophysActa.
2011,1810,1110-1113.
www.
proteomics-journal.
comPage46ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Figure1.
SchematicrepresentationofRAGEanditsisoforms(esRAGE-endogenoussecretoryRAGE,sRAGE-solubleRAGE,flRAGE-fulllengthRAGE)www.
proteomics-journal.
comPage47ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Figure2.
DepictionofRAGEactivatedsignalingpathwayindifferentdiseases.
www.
proteomics-journal.
comPage48ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Figure3.
WorkflowusedtoanalyzethesubstratesofRAGEactivatedkinasesfromPhosphositeplusdatabasewww.
proteomics-journal.
comPage49ProteomicsThisarticleisprotectedbycopyright.
Allrightsreserved.
Figure4.
TheplotdepictsassociationofphosphosubstrateswithdifferentdiseasesandRAGEactivatedkinases,andcolourcodeindicatesthediseaseassociation.
ForexampleERK2isasubstrateforERKandJNK,andisassociatedwithallfourdiseases.
- www.proteomics-journal.com相关文档
- 证书6kkbb.com
- P6KE10CA6kkbb.com
- review6kkbb.com
- 颐和新能源N5KTL、6KTL产品顺利通过德国T
- 11.36kkbb.com
- TZyPd6kkbb.com
香港最便宜的vps要多少钱?最便宜的香港vps能用吗?
香港最便宜的vps要多少钱?最便宜的香港vps能用吗?香港vps无需备案,整体性能好,而且租用价格便宜,使用灵活,因为备受站长喜爱。无论是个人还是企业建站,都比较倾向于选择香港VPS。最便宜的香港vps能用吗?正因为有着诸多租用优势,香港VPS在业内颇受欢迎,租用需求量也在日益攀升。那么,对于新手用户来说,香港最便宜的vps租用有四大要点是务必要注意的,还有易探云香港vps租用最便宜的月付仅18元...

华纳云E5处理器16G内存100Mbps688元/月
近日华纳云商家正式上线了美国服务器产品,这次美国机房上线的产品包括美国云服务器、美国独立服务器、美国高防御服务器以及美国高防云服务器等产品,新产品上线华纳云推出了史上优惠力度最高的特价优惠活动,美国云服务器低至3折,1核心1G内存5Mbps带宽低至24元/月,20G ddos高防御服务器低至688元/月,年付周期再送2个月、两年送4个月、三年送6个月,终身续费同价,有需要的朋友可以关注一下。华纳云...
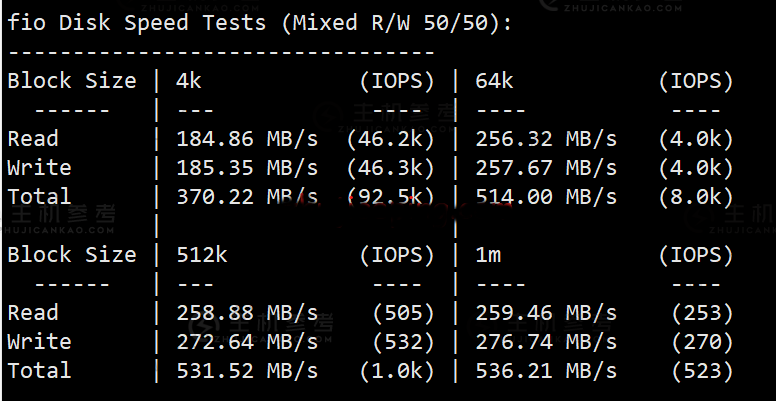
hypervmart:英国/荷兰vps,2核/3GB内存/25GB NVMe空间/不限流量/1Gbps端口/Hyper-V,$10.97/季
hypervmart怎么样?hypervmart是一家国外主机商,成立于2011年,提供虚拟主机、VPS等,vps基于Hyper-V 2012 R2,宣称不超售,支持linux和windows,有荷兰和英国2个数据中心,特色是1Gbps带宽、不限流量。现在配置提高,价格不变,性价比提高了很多。(数据中心不太清楚,按以前的记录,应该是欧洲),支持Paypal付款。点击进入:hypervmart官方网...
6kkbb.com为你推荐
-
敬汉卿姓名被抢注为什么最近b站up主都被问是否注册了商标?sherylsandbergLean In是一个怎样的组织johncusack谁知道《失控的陪审团》的电影内容是什么?约翰·库萨克在里面演的是什么角色?johncusack有喜欢演员JOHN CUSACK的吗?从哪部片子开始喜欢他的?至今为止他主要参与的电影作品有哪些?firetrap你们知道的有多少运动品牌的服饰?lunwenjiance知网论文检测查重系统网站检测请问,对网站进行监控检测的工具有哪些?mole.61.com摩尔庄园RK的秘密是什么?javbibinobibi的中文意思是?bbs2.99nets.com天堂1单机版到底怎么做